Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Babesia Life Cycle
3. Acute and Persistent Babesia Infections
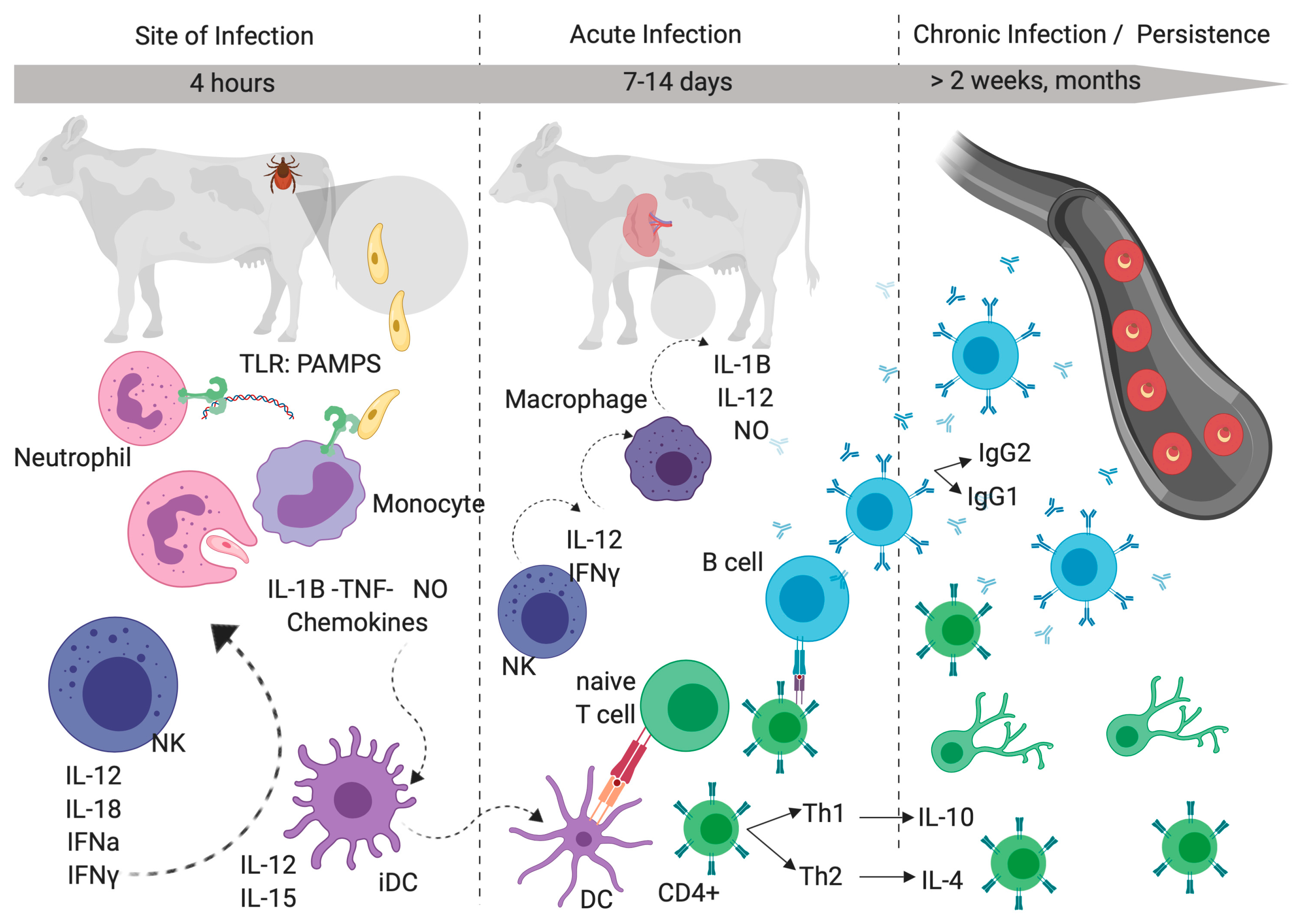
4. Immune Mechanisms of Persistent Babesia Infections
5. Interplay among Babesia and Its Hosts
6. Possible Role of Attenuation Factors in Modulating Persistent Infection and Virulence
7. Concluding Remarks: Proposed Model of a Possible Mechanism of Attenuation in Babesia bovis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suarez, C.E.; Bishop, R.P.; Alzan, H.F.; Poole, W.A.; Cooke, B.M. Advances in the application of genetic manipulation methods to 5 apicomplexan parasites. Int. J. Parasitol. 2017, 47, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Bartley, P.M.; Rocchi, M.; Benavidas-Silvan, J.; Burrells, A.; Hotchkiss, E.; Chianini, F.; Canton, G.; Katzer, F. Developing vaccines to control protozoan parasites in ruminants: Dead or alive? Vet. Parasitol. 2011, 180, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Janouškoveca, J.; Horáka, A.; Oborníkb, M.; Lukešb, J.; Keeling, P.J. A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids. Proc. Natl. Acad. Sci. USA 2010, 107, 10949–10954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- De Vos, A.J.; Bock, R.E. Vaccination against bovine babesiosis. Ann. N. Y. Acad. Sci. 2000, 916, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129 (Suppl. 1), S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of ixodes ricinus ticks in europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 1–18. [Google Scholar] [CrossRef]
- Hunfeld, K.P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef]
- Benavides, E.; Vizcaino, O.; Britto, C.M.; Romero, A.; Rubio, A. Attenuated trivalent vaccine against babesiosis and anaplasmosis in colombia. Ann. N. Y. Acad. Sci. 2000, 916, 613–616. [Google Scholar] [CrossRef]
- Callow, L.L.; Dalgliesh, R.J.; de Vos, A.J. Development of effective living vaccines against bovine babesiosis--the longest field trial? Int. J. Parasitol. 1997, 27, 747–767. [Google Scholar] [CrossRef]
- Bock, R.E.; de Vos, A.J. Immunity following use of australian tick fever vaccine: A review of the evidence. Aust. Vet. J. 2001, 79, 832–839. [Google Scholar] [CrossRef] [PubMed]
- de Waal, D.T.; Combrink, M.P. Live vaccines against bovine babesiosis. Vet. Parasitol. 2006, 138, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Dalgliesh, R.J.; Callow, L.L.; Mellors, L.T.; McGregor, W. Development of a highly infective babesia bigemina vaccine of reduced virulence. Aust. Vet. J. 1981, 57, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Nevils, M.A.; Figueroa, J.V.; Turk, J.R.; Canto, G.J.; Le, V.; Ellersieck, M.R.; Carson, C.A. Cloned lines of babesia bovis differ in their ability to induce cerebral babesiosis in cattle. Parasitol. Res. 2000, 86, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.G. An electron microscopic study of intravascular agglutination in the cerebral cortex due to babesia argentina infection. Int. J. Parasitol. 1972, 2, 209–215. [Google Scholar] [CrossRef]
- Sondgeroth, K.S.; McElwain, T.F.; Allen, A.J.; Chen, A.V.; Lau, A.O. Loss of neurovirulence is associated with reduction of cerebral capillary sequestration during acute babesia bovis infection. Parasites Vectors 2013, 6, 181. [Google Scholar] [CrossRef]
- Florin-Christensen, M.; Suarez, C.E.; Rodriguez, A.; Flores, D.; Schnittger, L. Vaccines against bovine babesiosis: Where we are now and possible roads ahead. Parasitology 2014, 141, 1563–1592. [Google Scholar] [CrossRef]
- Brown, W.C. Molecular approaches to elucidating innate and acquired immune responses to babesia bovis, a protozoan parasite that causes persistent infection. Vet. Parasitol. 2001, 101, 233–248. [Google Scholar] [CrossRef]
- Brown, W.C.; Palmer, G.H. Designing blood-stage vaccines against babesia bovis and b. Bigemina. Parasitol. Today 1999, 15, 275–281. [Google Scholar] [CrossRef]
- Shoda, L.K.; Palmer, G.H.; Florin-Christensen, J.; Florin-Christensen, M.; Godson, D.L.; Brown, W.C. Babesia bovis-stimulated macrophages express interleukin-1beta, interleukin-12, tumor necrosis factor alpha, and nitric oxide and inhibit parasite replication in vitro. Infect. Immun. 2000, 68, 5139–5145. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.R. Immune evasion by babesia bovis and plasmodium falciparum: Cliff-dwellers of the parasite world. Parasitol. Today 1995, 11, 100–105. [Google Scholar] [CrossRef]
- Smith, J.D.; Rowe, J.A.; Higgins, M.K.; Lavstsen, T. Malaria’s deadly grip: Cytoadhesion of plasmodium falciparum infected erythrocytes. Cell. Microbiol. 2013, 15, 1976–1983. [Google Scholar] [CrossRef] [PubMed]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.A.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. Embo Mol. Med. 2019, 11, e9164. [Google Scholar] [CrossRef] [PubMed]
- Jasmer, D.P.; Reduker, D.W.; Perryman, L.E.; McGuire, T.C. A babesia bovis 225-kilodalton protein located on the cytoplasmic side of the erythrocyte membrane has sequence similarity with a region of glycogen phosphorylase. Mol. Biochem. Parasitol. 1992, 52, 263–269. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Seuseu, F.J.; Eko-Wibowo, P.; Huyen, N.X.; Minoda, Y.; AbouLaila, M.; Kawai, S.; Yokoyama, N.; Xuan, X.; Igarashi, I. Secretion of a new spherical body protein of babesia bovis into the cytoplasm of infected erythrocytes. Mol. Biochem. Parasitol. 2011, 178, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Hines, S.A.; Palmer, G.H.; Brown, W.C.; McElwain, T.F.; Suarez, C.E.; Vidotto, O.; Rice-Ficht, A.C. Genetic and antigenic characterization of babesia bovis merozoite spherical body protein bb-1. Mol. Biochem. Parasitol. 1995, 69, 149–159. [Google Scholar] [CrossRef]
- Ruef, B.J.; Dowling, S.C.; Conley, P.G.; Perryman, L.E.; Brown, W.C.; Jasmer, D.P.; Rice-Ficht, A.C. A unique babesia bovis spherical body protein is conserved among geographic isolates and localizes to the infected erythrocyte membrane. Mol. Biochem. Parasitol. 2000, 105, 1–12. [Google Scholar] [CrossRef]
- O’Connor, R.M.; Lane, T.J.; Stroup, S.E.; Allred, D.R. Characterization of a variant erythrocyte surface antigen (vesa1) expressed by babesia bovis during antigenic variation. Mol. Biochem. Parasitol. 1997, 89, 259–270. [Google Scholar] [CrossRef]
- Allred, D.R. Variable and variant protein multigene families in babesia bovis persistence. Pathogens 2019, 8, 76. [Google Scholar] [CrossRef]
- Allred, D.R.; Cinque, R.M.; Lane, T.J.; Ahrens, K.P. Antigenic variation of parasite-derived antigens on the surface of babesia bovis-infected erythrocytes. Infect. Immun. 1994, 62, 91–98. [Google Scholar] [PubMed]
- Brown, W.C.; Estes, D.M.; Chantler, S.E.; Kegerreis, K.A.; Suarez, C.E. DNA and a cpg oligonucleotide derived from babesia bovis are mitogenic for bovine b cells. Infect. Immun. 1998, 66, 5423–5432. [Google Scholar] [PubMed]
- Brown, W.C.; Norimine, J.; Goff, W.L.; Suarez, C.E.; McElwain, T.F. Prospects for recombinant vaccines against babesia bovis and related parasites. Parasite Immunol. 2006, 28, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.C.; Cluff, C.W.; Goff, W.L.; Wyatt, C.R. Reactive oxygen and nitrogen intermediates and products from polyamine degradation are babesiacidal in vitro. Ann. N. Y. Acad. Sci. 1996, 791, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Goff, W.L.; Johnson, W.C.; Horn, R.H.; Barrington, G.M.; Knowles, D.P. The innate immune response in calves to boophilus microplus tick transmitted babesia bovis involves type-1 cytokine induction and nk-like cells in the spleen. Parasite Immunol. 2003, 25, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Goff, W.L.; Storset, A.K.; Johnson, W.C.; Brown, W.C. Bovine splenic nk cells synthesize ifn-gamma in response to il-12-containing supernatants from babesia bovis-exposed monocyte cultures. Parasite Immunol. 2006, 28, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.C.; Logan, K.S. Babesia bovis: Bovine helper T cell lines reactive with soluble and membrane antigens of merozoites. Exp. Parasitol. 1992, 74, 188–199. [Google Scholar] [CrossRef]
- Brown, W.C.; Woods, V.M.; Dobbelaere, D.A.; Logan, K.S. Heterogeneity in cytokine profiles of babesia bovis-specific bovine cd4+ t cells clones activated In Vitro. Infect. Immun. 1993, 61, 3273–3281. [Google Scholar]
- Goff, W.L.; Johnson, W.C.; Parish, S.M.; Barrington, G.M.; Tuo, W.; Valdez, R.A. The age-related immunity in cattle to babesia bovis infection involves the rapid induction of interleukin-12, interferon-gamma and inducible nitric oxide synthase mrna expression in the spleen. Parasite Immunol. 2001, 23, 463–471. [Google Scholar] [CrossRef]
- Echaide, I.E.; Hines, S.A.; McElwain, T.F.; Suarez, C.E.; McGuire, T.C.; Palmer, G.H. In Vivo binding of immunoglobulin m to the surfaces of babesia bigemina-infected erythrocytes. Infect. Immun. 1998, 66, 2922–2927. [Google Scholar]
- Battsetseg, B.; Sivakumar, T.; Naranbaatar, K.; Narantsatsral, S.; Myagmarsuren, P.; Enkhtaivan, B.; Davaasuren, B.; Mizushima, D.; Weerasooriya, G.; Igarashi, I.; et al. Serosurvey of babesia bovis and babesia bigemina in cattle in mongolia. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Lan, D.T.B.; Long, P.T.; Viet, L.Q.; Weerasooriya, G.; Kume, A.; Suganuma, K.; Igarashi, I.; Yokoyama, N. Serological and molecular surveys of babesia bovis and babesia bigemina among native cattle and cattle imported from thailand in hue, vietnam. J. Vet. Med. Sci. 2018, 80, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Guswanto, A.; Allamanda, P.; Mariamah, E.; Munkjargal, T.; Tuvshintulga, B.; Takemae, H.; Sivakumar, T.; AbouLaila, M.; Terkawi, M.; Ichikawa-Seki, M.; et al. Evaluation of immunochromatographic test (ict) strips for the serological detection of babesia bovis and babesia bigemina infection in cattle from western java, indonesia. Vet. Parasitol. 2017, 239, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Hines, S.A.; Palmer, G.H.; Jasmer, D.P.; McGuire, T.C.; McElwain, T.F. Neutralization-sensitive merozoite surface antigens of babesia bovis encoded by members of a polymorphic gene family. Mol. Biochem. Parasitol. 1992, 55, 85–94. [Google Scholar] [CrossRef]
- Lau, A.O.; Cereceres, K.; Palmer, G.H.; Fretwell, D.L.; Pedroni, M.J.; Mosqueda, J.; McElwain, T.F. Genotypic diversity of merozoite surface antigen 1 of babesia bovis within an endemic population. Mol. Biochem. Parasitol. 2010, 172, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Berens, S.J.; Brayton, K.A.; Molloy, J.B.; Bock, R.E.; Lew, A.E.; McElwain, T.F. Merozoite surface antigen 2 proteins of babesia bovis vaccine breakthrough isolates contain a unique hypervariable region composed of degenerate repeats. Infect. Immun. 2005, 73, 7180–7189. [Google Scholar] [CrossRef] [PubMed]
- Florin-Christensen, M.; Suarez, C.E.; Hines, S.A.; Palmer, G.H.; Brown, W.C.; McElwain, T.F. The babesia bovis merozoite surface antigen 2 locus contains four tandemly arranged and expressed genes encoding immunologically distinct proteins. Infect. Immun. 2002, 70, 3566–3575. [Google Scholar] [CrossRef]
- Lau, A.O.; Kalyanaraman, A.; Echaide, I.; Palmer, G.H.; Bock, R.; Pedroni, M.J.; Rameshkumar, M.; Ferreira, M.B.; Fletcher, T.I.; McElwain, T.F. Attenuation of virulence in an apicomplexan hemoparasite results in reduced genome diversity at the population level. Bmc Genom. 2011, 12, 410. [Google Scholar] [CrossRef]
- Pedroni, M.J.; Sondgeroth, K.S.; Gallego-Lopez, G.M.; Echaide, I.; Lau, A.O. Comparative transcriptome analysis of geographically distinct virulent and attenuated babesia bovis strains reveals similar gene expression changes through attenuation. Bmc Genom. 2013, 14, 763. [Google Scholar] [CrossRef]
- Gallego-Lopez, G.M.; Lau, A.O.T.; Brown, W.C.; Johnson, W.C.; Ueti, M.W.; Suarez, C.E. Spherical body protein 2 truncated copy 11 as a specific babesia bovis attenuation marker. Parasite Vectors 2018, 11, 1–8. [Google Scholar] [CrossRef]
- Wright, I. Ultrastructural changes in babesia argentina-infected erythrocytes in kidney capillaries. J. Parasitol. 1973, 59, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.R. Babesiosis: Persistence in the face of adversity. Trends Parasitol. 2003, 19, 51–55. [Google Scholar] [CrossRef]
- Gallego-Lopez, G.M.; Lau, A.O.T.; O’Connor, R.M.; Ueti, M.W.; Cooked, B.M.; Laughery, J.M.; Graça, T.; Madsen-Boutersea, S.A.; Oldiges, D.P.; Allred, D.R.; et al. Up-regulated expression of spherical body protein 2 truncated copy 11 in babesia bovis is associated with reduced cytoadhesion to vascular endothelial cells. Int. J. Parasitol. 2019, 49, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Al-Khedery, B.; Allred, D.R. Antigenic variation in babesia bovis occurs through segmental gene conversion of the ves multigene family, within a bidirectional locus of active transcription. Mol. Microbiol. 2006, 59, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.P.; Al-Khedery, B.; Allred, D.R. The babesia bovis vesa1 virulence factor subunit 1b is encoded by the 1beta branch of the ves multigene family. Mol. Biochem. Parasitol. 2010, 171, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.P.; Otto, T.D.; Darby, A.; Ramaprasad, A.; Xia1, D.; Echaide, I.E.; Farber, M.; Gahlot, S.; Gamble, J.; Gupta, D.; et al. The evolutionary dynamics of variant antigen genes in babesia reveal a history of genomic innovation underlying host–parasite interaction. Nucleic Acids Res. 2014, 42, 7113–7131. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xiao, Y.P.; Allred, D.R. Unusual chromatin structure associated with monoparalogous transcription of the babesia bovis ves multigene family. Int. J. Parasitol. 2012, 43, 163–172. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.M.; Allred, D.R. Selection of babesia bovis-infected erythrocytes for adhesion to endothelial cells coselects for altered variant erythrocyte surface antigen isoforms. J. Immunol. 2000, 164, 2037–2045. [Google Scholar] [CrossRef]
- Hutchings, C.L.; Li, A.; Fernandez, K.M.; Fletcher, T.; Jackson, L.A.; Molloy, J.B.; Jorgensen, W.K.; Lim, C.T.; Cooke, B.M. New insights into the altered adhesive and mechanical properties of red blood cells parasitized by babesia bovis. Mol. Microbiol. 2007, 65, 1092–1105. [Google Scholar] [CrossRef]
- Aikawa, M.; Rabbege, J.; Uni, S.; Ristic, M.; Miller, L.H. Structural alteration of the membrane of erythrocytes infected with babesia bovis. Am. J. Trop. Med. Hyg. 1985, 34, 45–49. [Google Scholar] [CrossRef]
- Weng, H.; Guo, X.; Papoin, J.; Wang, J.; Coppel, R.; Mohandas, N.; Ana, X. Interaction of plasmodium falciparum knob-associated histidine-rich protein (kahrp) with erythrocyte ankyrin r is required for its attachment to the erythrocyte membrane. Biochim. Biophys. Acta 2014, 1838, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Watermeyer, J.M.; Hale, V.L.; Hackett, F.; Clare, D.K.; Cutts, E.E.; Vakonakis, I.; Fleck, R.A.; Blackman, M.J.; Saibil, H.R. A spiral scaffold underlies cytoadherent knobs in plasmodium falciparum–infected erythrocytes. Blood 2016, 127, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Helms, G.; Dasanna, A.K.; Schwarz, U.S.; Lanzer, M. Modeling cytoadhesion of plasmodium falciparuminfected erythrocytes and leukocytes common principles and distinctive features. Febs Lett. 2016, 590, 1955–1971. [Google Scholar] [CrossRef] [PubMed]
- Flick, K.; Chen, Q. Var genes, pfemp1 and the human host. Mol. Biochem. Parasitol. 2004, 134, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.A.; Priyatosh, R.; Ashutosh, K.; Sarovar, B.N. Dynamic association of pfemp1 and kahrp in knobs mediates cytoadherence during plasmodium invasion. Sci. Rep. 2015, 5, 1–9. [Google Scholar]
- Dixon, M.W.; Kenny, S.; McMillan, P.J.; Hanssen, E.; Trenholme, K.R.; Gardiner, D.L.; Tilley, L. Genetic ablation of a maurer’s cleft protein prevents assembly of the plasmodium falciparum virulence complex. Mol. Microbiol. 2011, 81, 982–993. [Google Scholar] [CrossRef]
- Rug, M.; Prescott, S.W.; Fernandez, K.M.; Cooke, B.M.; Cowman, A.F. The role of kahrp domains in knob formation and cytoadherence of p falciparum-infected human erythrocytes. Blood 2006, 108, 370–378. [Google Scholar] [CrossRef]
- Tonkin-Hill, G.Q.; Trianty, L.; Noviyanti, R.; Nguyen, H.H.T.; Sebayang, B.F.; Lampah, D.A.; Marfurt, J.; Cobbold, S.A.; Rambhatla, J.S.; McConville, M.J.; et al. The plasmodium falciparum transcriptome in severe malaria reveals altered expression of genes involved in important processes including surface antigen±encoding var genes. PLoS Biol. 2018, 16, e2004328. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego-Lopez, G.M.; Cooke, B.M.; Suarez, C.E. Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle. Pathogens 2019, 8, 97. https://doi.org/10.3390/pathogens8030097
Gallego-Lopez GM, Cooke BM, Suarez CE. Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle. Pathogens. 2019; 8(3):97. https://doi.org/10.3390/pathogens8030097
Chicago/Turabian StyleGallego-Lopez, Gina M., Brian M. Cooke, and Carlos E. Suarez. 2019. "Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle" Pathogens 8, no. 3: 97. https://doi.org/10.3390/pathogens8030097