From Stable to Lab—Investigating Key Factors for Sudden Deaths Caused by Streptococcus suis

 ,
,
Abstract
:1. Introduction
2. Results
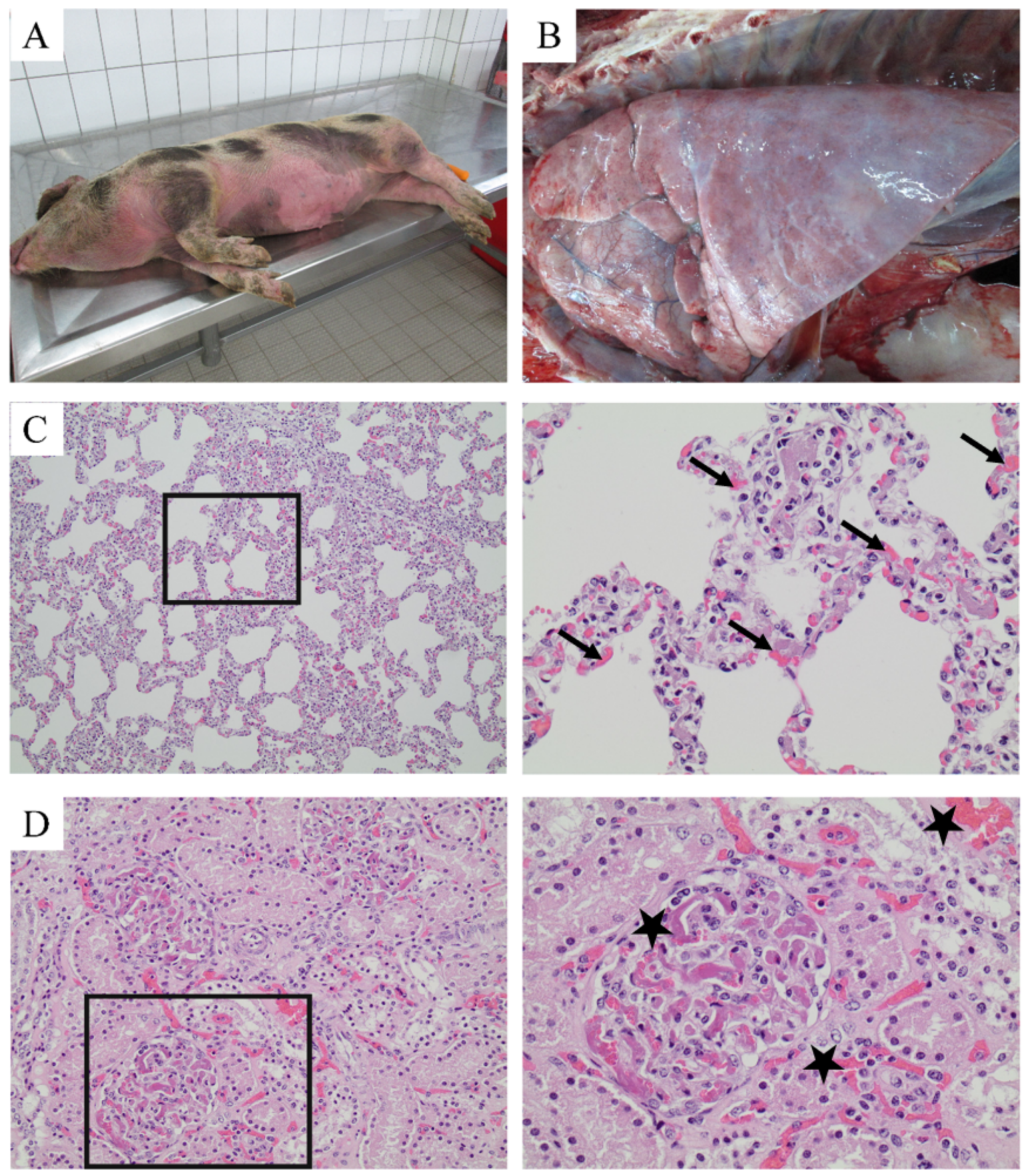
2.1. Diagnosis of Streptococcal Disease in a Zoo Pig Stock
2.2. Detection of Markers for Neutrophil Extracellular Traps (NETs) in Inner Organs Infected with S. suis and DIC Diagnosis
2.3. Genotypic Characterization of Isolated S. suis Strains
2.4. Phenotypic Characterization of S. suis Isolates
3. Discussion
4. Materials and Methods
4.1. Pathological and Histological Diagnostic of Dead Pigs
4.2. Collection of Diagnostic Samples from Pigs at the Zoo
4.3. Microbiological Analysis of Samples from Pigs
4.4. Genotypic Analysis of Isolated Strains
4.5. Cultivation of Bacteria
4.6. Cryostock Preparation for In Vitro Experiments
4.7. Ethical Statement for Collection of BALF, Porcine Blood, and Human Blood
4.8. Collection of BALF and Growth of S. suis in BALF
4.9. Staining of NETs in Histological Slices
4.10. Purification of Porcine Neutrophils and NET Induction Assay
4.11. DNase Activity Assay
4.12. Reconstituted Whole Blood Assay with Porcine Blood
4.13. Whole Blood Assay in Human Blood
4.14. Pico Green Assay
4.15. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A





References
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M. Streptococcosis. In Diseases of Swine; Zimmerman, J.J., Kar, L.A., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; Volume In Straw B, ISBN 081381703X. [Google Scholar]
- Pan, Z.; Ma, J.; Dong, W.; Song, W.; Wang, K.; Lu, C.; Yao, H. Novel Variant Serotype of Streptococcus suis Isolated from Piglets with Meningitis. Appl. Environ. Microbiol. 2015, 81, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, R.; Maruyama, F.; Ishida, S.; Tohya, M.; Sekizaki, T.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22 and 26: Streptococcus parasuis sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Tohya, M.; Arai, S.; Tomida, J.; Watanabe, T.; Kawamura, Y.; Katsumi, M.; Ushimizu, M.; Ishida-Kuroki, K.; Yoshizumi, M.; Uzawa, Y.; et al. Defining the taxonomic status of Streptococcus suis serotype 33: The proposal for Streptococcus ruminantium sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3660–3665. [Google Scholar] [CrossRef]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.-P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent—An update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, 1–20. [Google Scholar] [CrossRef]
- Clifton-Hadley, F.A. Streptococcus Suis Type 2 Infections. Br. Vet. J. 1983, 139, 1–5. [Google Scholar] [CrossRef]
- Sihvonen, L.; Kurl, D.N.; Salmela, P. Infection with Streptococcus suis serotypes 1 and 2 in the same diseased pig. Acta Vet. Scand. 1986, 27, 626–628. [Google Scholar]
- Brisebois, L.M.; Charlebois, R.; Higgins, R.; Nadeau, M. Prevalence of Streptococcus suis in four to eight week old clinically healthy piglets. Can. J. Vet. Res. 1990, 54, 174–177. [Google Scholar]
- Wisselink, H.J.; Smith, H.E.; Stockhofe-Zurwieden, N.; Peperkamp, K.; Vecht, U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet. Microbiol. 2000, 74, 237–248. [Google Scholar] [CrossRef]
- Silva, L.M.G.; Baums, C.G.; Rehm, T.; Wisselink, H.J.; Goethe, R.; Valentin-Weigand, P. Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet. Microbiol. 2006, 115, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Du, P.; Qiu, X.; Kerdsin, A.; Roy, D.; Bai, X.; Xu, J.; Vela, A.I.; Gottschalk, M. Genomic comparisons of Streptococcus suis serotype 9 strains recovered from diseased pigs in Spain and Canada. Vet. Res. 2018, 49, 1. [Google Scholar] [CrossRef] [PubMed]
- King, S.J.; Leigh, J.A.; Heath, P.J.; Luque, I.; Tarradas, C.; Dowson, C.G.; Whatmore, A.M. Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis: Identification of virulent clones and potential capsular serotype exchange. J. Clin. Microbiol. 2002, 40, 3671–3680. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.A.; Gottschalk, M.; Rossow, S.; Rendahl, A.; Gebhart, C.; Marthaler, D.G. Serotype and Genotype (Multilocus Sequence Type) of Streptococcus suis Isolates from the United States Serve as Predictors of Pathotype. J. Clin. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Torremorell, M.; Pijoan, C.; Trigo, E. Vaccination against Streptococcus suis: Effect on nursery mortality. J. Swine Heal. Prod. 1997, 5, 139–143. [Google Scholar]
- Staats, J.J.; Feder, I.; Okwumabua, O.; Chengappa, M.M. Streptococcus suis: Past and present. Vet. Res. Commun. 1997, 21, 381–407. [Google Scholar] [CrossRef]
- Clifton-Hadley, F.; Alexander, T. The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet. Rec. 1980, 107, 40–41. [Google Scholar] [CrossRef]
- Lamont, M.H.; Edwards, P.T.; Windsor, R.S. Streptococcal meningitis in pigs: Results of a five-year survey. Vet. Rec. 1980, 107, 467–469. [Google Scholar] [CrossRef]
- Dione, M.; Masembe, C.; Akol, J.; Amia, W.; Kungu, J.; Lee, H.S.; Wieland, B. The importance of on-farm biosecurity: Sero-prevalence and risk factors of bacterial and viral pathogens in smallholder pig systems in Uganda. Acta Trop. 2018, 187, 214–221. [Google Scholar] [CrossRef]
- Kernaghan, S.; Bujold, A.R.; MacInnes, J.I. The microbiome of the soft palate of swine. Anim. Heal. Res. Rev. 2012, 13, 110–120. [Google Scholar] [CrossRef]
- MacInnes, J.I.; Gottschalk, M.; Lone, A.G.; Metcalf, D.S.; Ojha, S.; Rosendal, T.; Watson, S.B.; Friendship, R.M. Prevalence of Actinobacillus pleuropneumoniae, Actinobacillus suis, Haemophilus parasuis, Pasteurella multocida, and Streptococcus suis in representative Ontario swine herds. Can. J. Vet. Res. 2008, 72, 242–248. [Google Scholar] [PubMed]
- Gottschalk, M.; Segura, M. The pathogenesis of the meningitis caused by Streptococcus suis: The unresolved questions. Vet. Microbiol. 2000, 76, 259–272. [Google Scholar] [CrossRef]
- Warneboldt, F.; Sander, S.; Beineke, A.; Valentin-Weigand, P.; Kamphues, J.; Baums, C. Clearance of Streptococcus suis in Stomach Contents of Differently Fed Growing Pigs. Pathogens 2016, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, M.L.; de Greeff, A.; van Rooijen, W.J.M.; Stockhofe-Zurwieden, N.; Nielsen, J.; Wichgers Schreur, P.J.; Pannekoek, Y.; Heuvelink, A.; van der Ende, A.; Smith, H.; et al. Host-pathogen Interaction at the Intestinal Mucosa Correlates With Zoonotic Potential of Streptococcus suis. J. Infect. Dis. 2015, 212, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, M.L.; Willemse, N.; Zaccaria, E.; Pannekoek, Y.; van der Ende, A.; Schultsz, C. Streptococcal Adhesin P (SadP) contributes to Streptococcus suis adhesion to the human intestinal epithelium. PLoS ONE 2017, 12, e0175639. [Google Scholar] [CrossRef]
- Seele, J.; Singpiel, A.; Spoerry, C.; Von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. Identification of a Novel Host-Specific IgM Protease in Streptococcus suis. J. Bacteriol. 2013, 195, 930–940. [Google Scholar] [CrossRef]
- Smith, H.E.; Damman, M.; Van Der Velde, J.; Wagenaar, F.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Smits, M.A. Identification and characterization of the cps locus of Streptococcus suis serotype 2: The capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 1999, 67, 1750–1756. [Google Scholar]
- Prüfer, T.L.; Rohde, J.; Verspohl, J.; Rohde, M.; de Greeff, A.; Willenborg, J.; Valentin-Weigand, P. Molecular typing of Streptococcus suis strains isolated from diseased and healthy pigs between 1996–2016. PLoS ONE 2019, 14, e0210801. [Google Scholar] [CrossRef]
- Zheng, H.; Qiu, X.; Roy, D.; Segura, M.; Du, P.; Xu, J.; Gottschalk, M. Genotyping and investigating capsular polysaccharide synthesis gene loci of non-serotypeable Streptococcus suis isolated from diseased pigs in Canada. Vet. Res. 2017, 48, 10. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- de Buhr, N.; Reuner, F.; Neumann, A.; Stump-Guthier, C.; Tenenbaum, T.; Schroten, H.; Ishikawa, H.; Müller, K.; Beineke, A.; Hennig-Pauka, I.; et al. Neutrophil extracellular trap formation in the Streptococcus suis-infected cerebrospinal fluid compartment. Cell. Microbiol. 2017, 19, e12649. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Zychlinsky, A. NETs: A new strategy for using old weapons. Trends Immunol. 2009, 30, 513–521. [Google Scholar] [CrossRef] [PubMed]
- de Buhr, N.; von Köckritz-Blickwede, M. How Neutrophil Extracellular Traps Become Visible. J. Immunol. Res. 2016, 2016, 4604713. [Google Scholar] [CrossRef] [PubMed]
- Nel, J.G.; Theron, A.J.; Durandt, C.; Tintinger, G.R.; Pool, R.; Mitchell, T.J.; Feldman, C.; Anderson, R. Pneumolysin activates neutrophil extracellular trap formation. Clin. Exp. Immunol. 2016, 184, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Aulik, N.A.; Hellenbrand, K.M.; Czuprynski, C.J. Mannheimia haemolytica and its leukotoxin cause macrophage extracellular trap formation by bovine macrophages. Infect. Immun. 2012, 80, 1923–1933. [Google Scholar] [CrossRef] [Green Version]
- de Buhr, N.; Neumann, A.; Jerjomiceva, N.; von Köckritz-Blickwede, M.; Baums, C.G. Streptococcus suis DNase SsnA contributes to degradation of neutrophil extracellular traps (NETs) and evasion of NET-mediated antimicrobial activity. Microbiology 2014, 160, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Buhr, N.; Stehr, M.; Neumann, A.; Naim, H.Y.; Valentin-Weigand, P.; von Köckritz-Blickwede, M.; Baums, C.G. Identification of a novel DNase of Streptococcus suis (EndAsuis) important for neutrophil extracellular trap degradation during exponential growth. Microbiology 2015, 161, 838–850. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.S.; Carmona-Rivera, C.; Kaplan, M.J. Proteins derived from neutrophil extracellular traps may serve as self-antigens and mediate organ damage in autoimmune diseases. Front. Immunol. 2012, 3, 380. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.S.; Kaplan, M.J. Lupus neutrophils: ‘NET’ gain in understanding lupus pathogenesis. Curr. Opin. Rheumatol. 2012, 24, 441–450. [Google Scholar] [CrossRef]
- Hakkim, A.; Furnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, E.; van der Vlag, J. Breaking Immunological Tolerance in Systemic Lupus Erythematosus. Front. Immunol. 2014, 5, 164. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, B.; Urrutia, R.; Yipp, B.G.; Jenne, C.N.; Kubes, P. Intravascular neutrophil extracellular traps capture bacteria from the bloodstream during sepsis. Cell Host Microbe 2012, 12, 324–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhamdi, Y.; Toh, C.-H. Recent advances in pathophysiology of disseminated intravascular coagulation: The role of circulating histones and neutrophil extracellular traps. F1000Research 2017, 6, 2143. [Google Scholar] [CrossRef] [Green Version]
- Athey, T.B.T.; Teatero, S.; Lacouture, S.; Takamatsu, D.; Gottschalk, M.; Fittipaldi, N. Determining Streptococcus suis serotype from short-read whole-genome sequencing data. BMC Microbiol. 2016, 16, 162. [Google Scholar] [CrossRef] [Green Version]
- Kock, C.; Beineke, A.; Seitz, M.; Ganter, M.; Waldmann, K.H.; Valentin-Weigand, P.; Baums, C.G. Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Vet. Immunol. Immunopathol. 2009, 132, 135–145. [Google Scholar] [CrossRef]
- Seele, J.; Hillermann, L.-M.; Beineke, A.; Seitz, M.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. The immunoglobulin M-degrading enzyme of Streptococcus suis, IdeSsuis, is a highly protective antigen against serotype 2. Vaccine 2015, 33, 2207–2212. [Google Scholar] [CrossRef]
- Unterweger, C.; Ruczizka, U.; Spergser, J.; Baums, C.G.; Hennig-Pauka, I. Effect of Early-Life Treatment of Piglets with Long-Acting Ceftiofur on Colonization of Streptococcus suis Serotype 7 and Elicitation of Specific Humoral Immunity in a Farm Dealing with Streptococcal Diseases. Pathogens 2018, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Baele, M.; Chiers, K.; Devriese, L.A.; Smith, H.E.; Wisselink, H.J.; Vaneechoutte, M.; Haesebrouck, F. The gram-positive tonsillar and nasal flora of piglets before and after weaning. J. Appl. Microbiol. 2001, 91, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Tong, J.; Vötsch, D.; Peng, J.-Y.; Cai, X.; Willenborg, M.; Herrler, G.; Wu, N.-H.; Valentin-Weigand, P. Viral Coinfection Replaces Effects of Suilysin on Streptococcus suis Adherence to and Invasion of Respiratory Epithelial Cells Grown under Air-Liquid Interface Conditions. Infect. Immun. 2019, 87, e00350-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, T.; Hudemann, C.; Hossain, H.M.; Hewer, A.; Tello, K.; Bandorski, D.; Rohde, M.; Valentin-Weigand, P.; Baums, C.G. Characterization of Five Zoonotic Streptococcus suis Strains from Germany, Including One Isolate from a Recent Fatal Case of Streptococcal Toxic Shock-Like Syndrome in a Hunter. J. Clin. Microbiol. 2015, 53, 3912–3915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecht, U.; Wisselink, H.J.; Jellema, M.L.; Smith, H.E. Identification of two proteins associated with virulence of Streptococcus suis type 2. Infect. Immun. 1991, 59, 3156–3162. [Google Scholar] [PubMed]
- Fittipaldi, N.; Xu, J.; Lacouture, S.; Tharavichitkul, P.; Osaki, M.; Sekizaki, T.; Takamatsu, D.; Gottschalk, M. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg. Infect. Dis. 2011, 17, 2239–2244. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Athey, T.B.T.; Auger, J.-P.; Teatero, S.; Dumesnil, A.; Takamatsu, D.; Wasserscheid, J.; Dewar, K.; Gottschalk, M.; Fittipaldi, N. Complex Population Structure and Virulence Differences among Serotype 2 Streptococcus suis Strains Belonging to Sequence Type 28. PLoS ONE 2015, 10, e0137760. [Google Scholar] [CrossRef]
- Chang, B.; Wada, A.; Ikebe, T.; Ohnishi, M.; Mita, K.; Endo, M.; Matsuo, H.; Asatuma, Y.; Kuramoto, S.; Sekiguchi, H.; et al. Characteristics of Streptococcus suis isolated from patients in Japan. Jpn. J. Infect. Dis. 2006, 59, 397–399. [Google Scholar]
- Lin, L.; Xu, L.; Lv, W.; Han, L.; Xiang, Y.; Fu, L.; Jin, M.; Zhou, R.; Chen, H.; Zhang, A. An NLRP3 inflammasome-triggered cytokine storm contributes to Streptococcal toxic shock-like syndrome (STSLS). PLoS Pathog. 2019, 15, e1007795. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, J.; Yu, J.; Xu, Z.; Liu, L.; Song, Y.; Sun, X.; Zhang, A.; Jin, M. HP1330 contributes to Streptococcus suis virulence by inducing toll-like receptor 2- and ERK1/2-dependent pro-inflammatory responses and influencing in vivo S. suis loads. Front. Immunol. 2017, 8, 869. [Google Scholar] [CrossRef]
- Band, D.E. The use of a phenolic disinfectant in animal husbandry. Int. Biodeterior. 1990, 26, 217–223. [Google Scholar] [CrossRef]
- Postma, M.; Backhans, A.; Collineau, L.; Loesken, S.; Sjölund, M.; Belloc, C.; Emanuelson, U.; Grosse Beilage, E.; Stärk, K.D.C.; Dewulf, J. The biosecurity status and its associations with production and management characteristics in farrow-to-finish pig herds. Animal 2016, 10, 478–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanojkovic, А.; Petrovic, М.M.; Skrbic, Z.; Mandic, V.; Stanisic, N.; Gogic, M.; Stanojkovic-Sebic, A. Biochemical characteristics of Streptococcus suis strains isolated from healthy and deceased pigs. Biotechnol. Anim. Husb. 2014, 30, 699–704. [Google Scholar] [CrossRef]
- Rieckmann, K.; Seydel, A.; Szewczyk, K.; Klimke, K.; Rungelrath, V.; Baums, C.G. Streptococcus suis cps7: An emerging virulent sequence type (ST29) shows a distinct, IgM-determined pattern of bacterial survival in blood of piglets during the early adaptive immune response after weaning. Vet. Res. 2018, 49, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, T.; Baums, C.G.; Strommenger, B.; Beyerbach, M.; Valentin-Weigand, P.; Goethe, R. Amplified fragment length polymorphism of Streptococcus suis strains correlates with their profile of virulence-associated genes and clinical background. J. Med. Microbiol. 2007, 56, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.E.; Reek, F.H.; Vecht, U.; Gielkens, A.L.J.; Smits, M.A. Repeats in an extracellular protein of weakly pathogenic strains of Streptococcus suis type 2 are absent in pathogenic strains. Infect. Immun. 1993, 61, 3318–3326. [Google Scholar]
- Sassu, E.L.; Frömbling, J.; Duvigneau, J.C.; Miller, I.; Müllebner, A.; Gutiérrez, A.M.; Grunert, T.; Patzl, M.; Saalmüller, A.; von Altrock, A.; et al. Host-pathogen interplay at primary infection sites in pigs challenged with Actinobacillus pleuropneumoniae. BMC Vet. Res. 2017, 13, 64. [Google Scholar] [CrossRef] [Green Version]
- de Buhr, N.; Bonilla, M.C.; Pfeiffer, J.; Akhdar, S.; Schwennen, C.; Kahl, B.C.; Waldmann, K.; Valentin-Weigand, P.; Hennig-Pauka, I.; von Köckritz-Blickwede, M. Degraded neutrophil extracellular traps promote the growth of Actinobacillus pleuropneumoniae. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Beineke, A.; Bennecke, K.; Neis, C.; Schröder, C.; Waldmann, K.-H.; Baumgärtner, W.; Valentin-Weigand, P.; Baums, C.G. Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet. Microbiol. 2008, 128, 423–430. [Google Scholar] [CrossRef]
- Benga, L.; Fulde, M.; Neis, C.; Goethe, R.; Valentin-Weigand, P. Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet. Microbiol. 2008, 132, 211–219. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Clinical findings | Pathological findings | Bacteriological findings | Diagnosis |
|---|---|---|---|---|
| 10/18 | sudden death, 38.5 kg | disseminated intravascular coagulopathy | S. suis (lung, heart, spleen, kidney) | systemic disease |
| 10/18 | sudden death, 33.5 kg | disseminated intravascular coagulopathy | S. suis (lung, heart, spleen, kidney) | systemic disease |
| 12/18 | sudden death, 47 kg | fibrinopurulent carpitis, fibrinopurulent leptomeningitis, hemorrhagic gastritis and jejunitis | no isolate | treated (antibiotics) |
| 12/18 | sudden death, 103 kg | fibrinopurulent epi- and pericarditis, leptomeningitis | no isolate | treated (antibiotics) |
| 12/18 | sudden death, 38 kg | purulent leptomeningitis, pleuritis, epicarditis, hemorrhagic gastritis, jejunitis | Haemophilus parasuis ST 13 (brain, lung, heart) | Glaesser’s disease |
| Name | Date | Origin | Capsular type | mrp | sly | epf* | gdh | arcA | ST |
|---|---|---|---|---|---|---|---|---|---|
| 483 | 10/18 | pig 1, lung | 2 | + | - | - | + | + | 28 |
| 484 | 10/18 | pig 2, spleen | 2 | + | - | - | + | + | 28 |
| T17 | 01/19 | pig 3, tonsil | n.t. | - | - | - | + | + | 1173 |
| T28 | 01/19 | pig 4, tonsil | n.t. | - | - | - | + | + | x |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hennig-Pauka, I.; Imker, R.; Mayer, L.; Brügmann, M.; Werckenthin, C.; Weber, H.; Menrath, A.; de Buhr, N. From Stable to Lab—Investigating Key Factors for Sudden Deaths Caused by Streptococcus suis. Pathogens 2019, 8, 249. https://doi.org/10.3390/pathogens8040249
Hennig-Pauka I, Imker R, Mayer L, Brügmann M, Werckenthin C, Weber H, Menrath A, de Buhr N. From Stable to Lab—Investigating Key Factors for Sudden Deaths Caused by Streptococcus suis. Pathogens. 2019; 8(4):249. https://doi.org/10.3390/pathogens8040249
Chicago/Turabian StyleHennig-Pauka, Isabel, Rabea Imker, Leonie Mayer, Michael Brügmann, Christiane Werckenthin, Heike Weber, Andrea Menrath, and Nicole de Buhr. 2019. "From Stable to Lab—Investigating Key Factors for Sudden Deaths Caused by Streptococcus suis" Pathogens 8, no. 4: 249. https://doi.org/10.3390/pathogens8040249