Prevalence, Pathogenicity, Virulence, Antibiotic Resistance, and Phylogenetic Analysis of Biofilm-Producing Listeria monocytogenes Isolated from Different Ecological Niches in Egypt: Food, Humans, Animals, and Environment


Abstract
1. Introduction
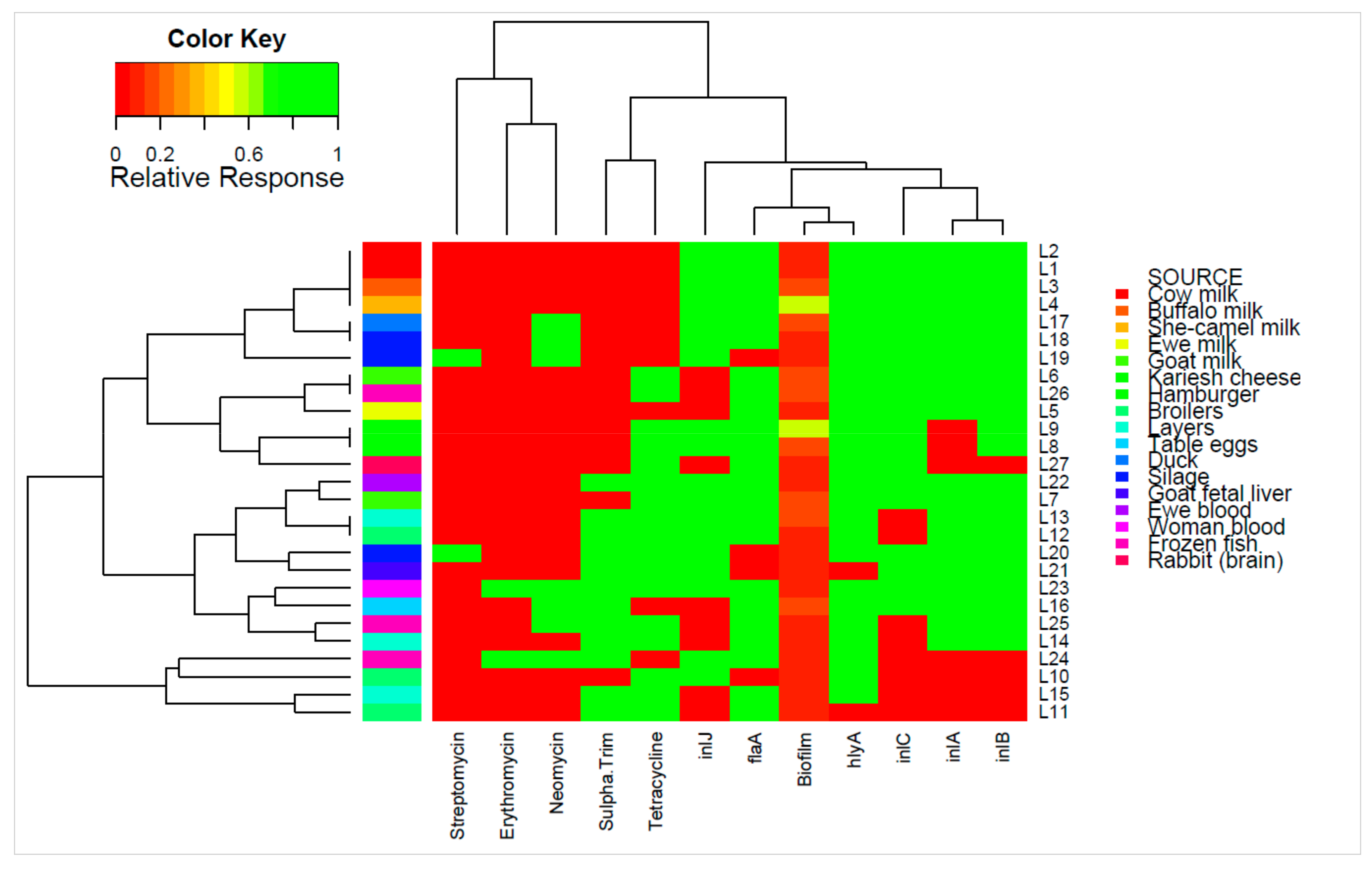
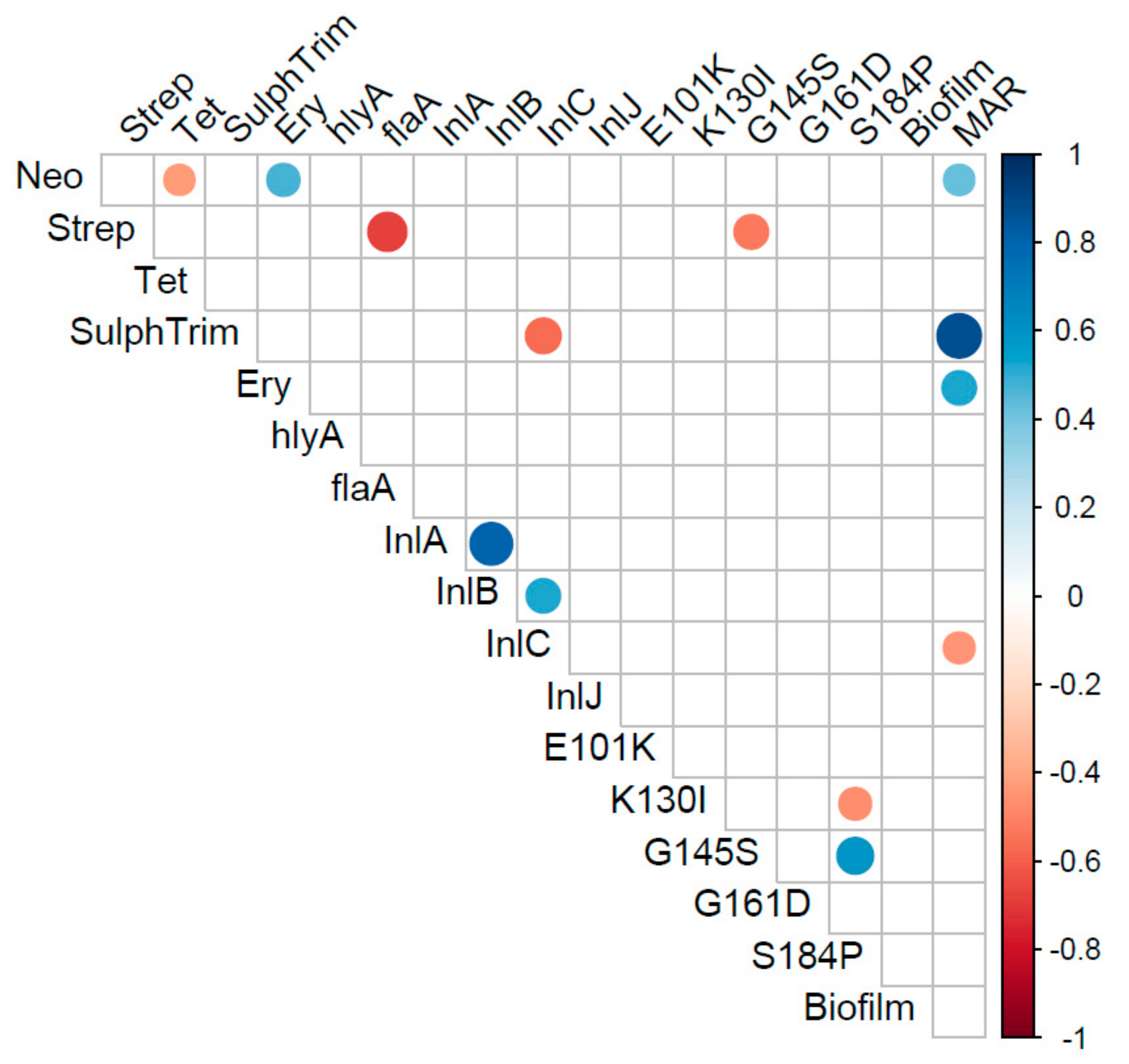
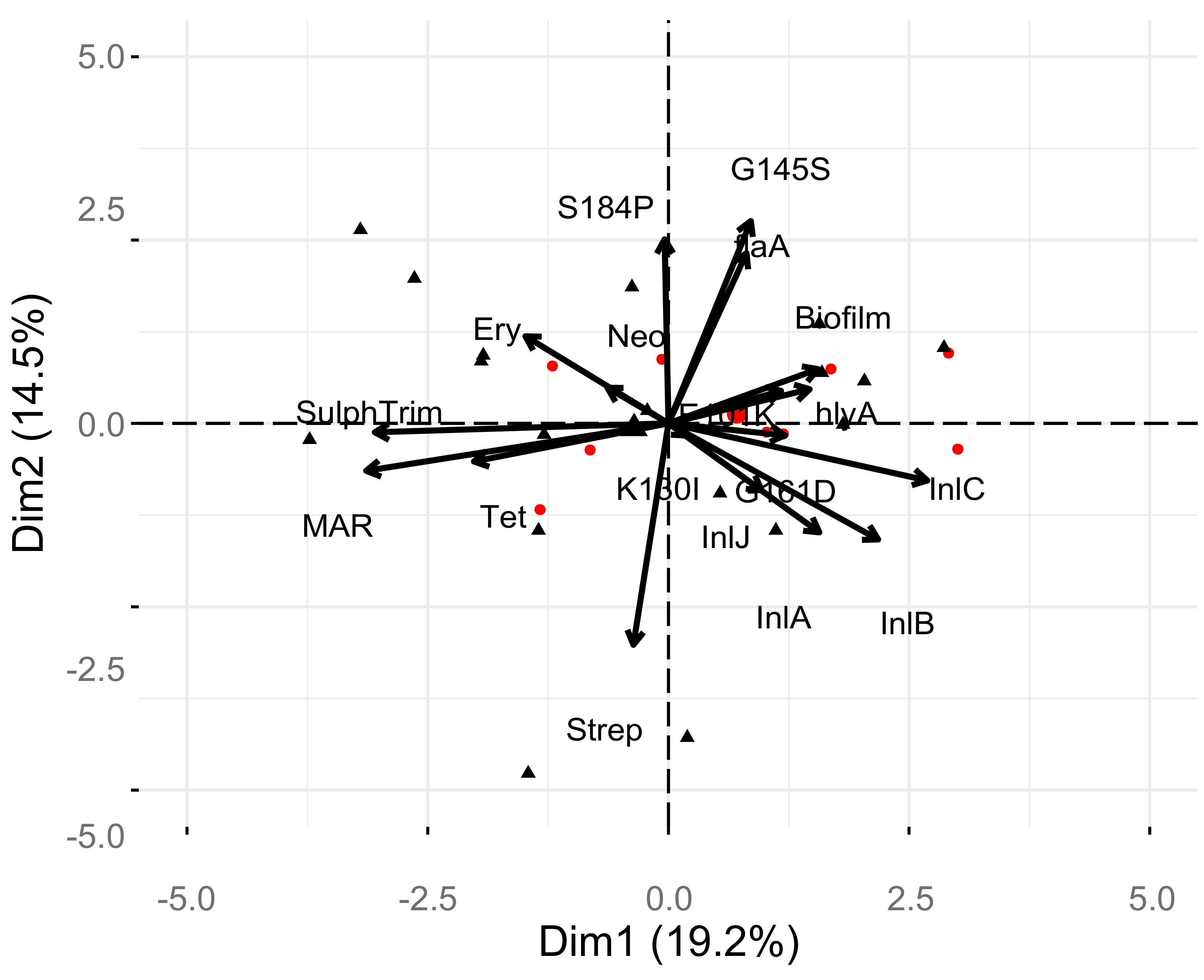
2. Results
prfA Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Food and Clinical Samples
4.2. Isolation, Identification, and Serotyping of L. monocytogenes
4.3. Antibiogram Profile
4.4. Pathogenicity and Biofilm Formation
4.5. Molecular Confirmation of L. monocytogenes and Detection of Virulence Genes
4.6. Statistical Analysis
4.7. Phylogenetic Analysis of the prfA gene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Adgamov, R.; Zaytseva, E.; Thiberge, J.; Brisse, S.; Ermolaeva, S. Genetically related Listeria Monocytogenes strains isolated from lethal human cases and wild animals. In Genetic Diversity in Microorganisms; Caliskan, M., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Linke, K.; Rückerl, I.; Brugger, K.; Karpiskova, R.; Walland, J.; Muri-Klinger, S.; Tichy, A.; Wagner, M.; Stessl, B. Reservoirs of Listeria species in three environmental ecosystems. Appl. Environ. Microbiol. 2014, 80, 5583–5592. [Google Scholar] [CrossRef] [PubMed]
- Jami, M.; Ghanbari, M.; Zunabovic, M.; Domig, K.J.; Kneifel, W. Listeria monocytogenes in aquatic food products—A review. Comp. Rev. Food Sci. Food Saf. 2014, 13, 798–813. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSAJ 2017, 15, e5077. [Google Scholar]
- Nastasijevica, I.; Milanov, D.; Velebit, B.; Djordjevic, V.; Swift, C.; Painset, A.; Lakicevic, B. Tracking of Listeria monocytogenes in meat establishment using Whole Genome Sequencing as a food safety management tool: A proof of concept. Int. J. Food Microbiol. 2017, 257, 157–164. [Google Scholar] [CrossRef]
- Editors of Encyclopædia Britannica, Listeriosis Pathology, Last Updated: 29 November 2018.
- Tan, M.F.; Siow, C.C.; Dutta, A.A.; Mutha, N.V.R.; Wee, W.Y.; Heydari, H.; Tan, S.Y.; Ang, M.Y.; Wong, G.J.; Choo, S.W. Development of ListeriaBase and comparative analysis of Listeria monocytogenes. BMC Genom. 2015, 16, 755. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef]
- Buchanana, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Hingston, P.; Chen, J.; Dhillon, B.K.; Laing, C.; Bertelli, C.; Gannon, V.; Tasara, T.; Allen, K.; Brinkman, F.S.; Truelstrup Hansen, L.; et al. Genotypes Associated with Listeria monocytogenes isolates displaying impaired or enhanced tolerances to cold, salt, acid, or desiccation stress. Front. Microbiol. 2017, 8, 369. [Google Scholar] [CrossRef]
- Jennison, A.V.; Masson, J.J.; Fang, N.X.; Graham, R.M.; Bradbury, M.I.; Fegan, N.; Gobius, K.S.; Graham, T.M.; Guglielmino, C.J.; Brown, J.L.; et al. Analysis of the Listeria monocytogenes population structure among isolates from 1931 to 2015 in Australia. Front. Microbiol. 2017, 8, 603. [Google Scholar] [CrossRef]
- Chenal-Francisque, V.; Lopez, J.; Cantinelli, T.; Caro, V.; Tran, C.; Leclercq, A.; Lecuit, M.; Brisse, S. Worldwide distribution of major clones of Listeria monocytogenes. Emerg. Infect. Dis. 2011, 17, 1110–1112. [Google Scholar] [CrossRef]
- Ariza-Miguel, J.; Fernández-Natal, M.I.; Soriano, F.; Hernández, M.; Stessl, B.; Rodríguez-Lázaro, D. Molecular epidemiology of invasive listeriosis due to Listeria monocytogenes in a Spanish hospital over a nine-year study period, 2006–2014. BioMed Res. Int. 2015, 2015, 191409. [Google Scholar] [CrossRef] [PubMed]
- Voronina, O.L.; Ryzhova, N.; Marina, K.; Andrey, S.; Ekaterina, A.; Irina, E.; Denis, K.; Ermolaeva, S.; Alexander, L.G. Diversity and pathogenic potential of Listeria monocytogenes isolated from environmental sources in the Russian Federation. Int. J. Modern Eng. Res. (IJMER) 2015, 5, 5–15. [Google Scholar]
- Marini, E.; Magi, G.; Vincenzi, C.; Manso, E.; Facinelli, B. Ongoing outbreak of invasive listeriosis due to serotype 1/2a Listeria monocytogenes, Ancona province, Italy, January 2015 to February 2016. Euro. Surveill. 2016, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- EFSA Biohaz Panel (EFSA Panel on Biological Hazards); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Salvador, P.F.E.; Girones, R.; Herman, L.; Koutsoumanis, K. Scientific Opinion on the Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSAJ 2018, 16, e05134. [Google Scholar]
- Wadhwa, D.R.; Smith, M.A. Pregnancy-related listeriosis. Birth Defects Res. 2017, 109, 324–335. [Google Scholar] [CrossRef]
- Madjunkov, M.; Chaudhry, S.; Ito, S. Listeriosis during pregnancy. Arch. Gynecol. Obstet. 2017, 296, 143–152. [Google Scholar] [CrossRef]
- Acciari, V.A.; Iannetti, L.; Gattuso, A.; Sonnessa, M.; Scavia, G.; Montagna, C.; Addante, N.; Torresi, M.; Zocchi, L.; Scattolini, S.; et al. Tracing sources of Listeria contamination in traditional Italian cheese associated with a US outbreak: Investigations in Italy. Epidemiol. Infect. 2016, 144, 2719–2727. [Google Scholar] [CrossRef]
- Althaus, D.; Hofer, E.; Corti, S.; Julmi, A.; Stephan, R. Bacteriological Survey of Ready-to-Eat Lettuce, Fresh-Cut Fruit, and Sprouts Collected from the Swiss Market. J. Food Prot. 2012, 75, 1338–1341. [Google Scholar] [CrossRef]
- Heiman, K.E.; Garalde, V.B.; Gronostaj, M.; Jackson, K.A.; Beam, S.; Joseph, L.; Saupe, A.; Ricotta, E.; Waechter, H.; Wellman, A.; et al. Multistate outbreak of listeriosis caused by imported cheese and evidence of cross-contamination of other cheeses, USA, 2012. Epidemiol. Infect. 2016, 144, 2698–2708. [Google Scholar] [CrossRef]
- Henriques, A.R.; Cristino, J.M.; Fraqueza, M.J. Genetic characterization of Listeria monocytogenes Isolates from industrial and retail ready-to-eat meat-based foods and their relationship with clinical strains from human listeriosis in Portugal. J. Food Prot. 2017, 80, 551–560. [Google Scholar] [CrossRef]
- Henriques, A.R.; Gama, L.T.; Fraqueza, M.J. Tracking Listeria monocytogenes contamination and virulence-associated characteristics in the ready-to-eat meat-based food products industry according to the hygiene level. Int. J. Food Microbiol. 2017, 242, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods (Basel, Switzerland) 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control; European Food Safety Authority. Multi-country outbreak of Listeria monocytogenes sequence type 8 infections linked to consumption of salmon products –25 October 2018. Stockholm and Parma. ECDC/EFSA J. 2018, 15, 1496E. [Google Scholar]
- WHO. Listeriosis Fact sheet February; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Chen, J.Q.; Regan, P.; Laksanalamai, P.; Healey, S.; Hu, Z. Prevalence and methodologies for detection, characterization and subtyping of Listeria monocytogenes and L. ivanovii in foods and environmental sources. Food Sci. Human Wellness 2017, 6, 97–120. [Google Scholar] [CrossRef]
- Usman, U.B.; Kwaga, J.K.; Kabir, J.; Olonitola, O.S.; Radu, S.; Bande, F. Molecular characterization and phylogenetic analysis of Listeria monocytogenes isolated from milk and milk products in Kaduna, Nigeria. Can. J. Infect. Dis. Med. Microbiol. 2016, 2016, 4313827. [Google Scholar] [CrossRef]
- Raheem, D. Outbreaks of listeriosis associated with deli meats and cheese: an overview. AIMS Microbiol. 2016, 2, 230–250. [Google Scholar] [CrossRef]
- Vasanthakrishnan, R.B.; de Las Heras, A.; Scortti, M.; Deshayes, C.; Colegrave, N.; Vázquez-Boland, J.A. PrfA regulation offsets the cost of Listeria virulence outside the host. Environ. Microbiol. 2015, 17, 4566–4579. [Google Scholar] [CrossRef]
- D’Amico, D.J.; Donnelly, C.W. Microbiological quality of raw milk used for small-scale artisan cheese production in Vermont: Effect of farm characteristics and practices. J. Dairy Sci. 2010, 93, 134e147. [Google Scholar] [CrossRef]
- Osman, K.M.; Zolnikov, T.R.; Samir, A.; Orabi, A. Prevalence, pathogenic capability, virulence genes, biofilm formation, and antibiotic resistance of Listeria in goat and sheep milk confirms need of hygienic milking conditions. Pathog. Glob. Health 2014, 108, 21–29. [Google Scholar] [CrossRef]
- Osman, K.M.; Samir, A.; Abo-Shama, U.H.; Mohamed, E.H.; Orabi, A.; Zolnikov, T. Determination of virulence and antibiotic resistance pattern of biofilm producing Listeria species isolated from retail raw milk. BMC Microbiol. 2016, 16, 1–13. [Google Scholar] [CrossRef]
- Osman, K.M.; Samir, A.; Orabi, A.; Zolnikov, T.R. Confirmed low prevalence of Listeria mastitis in she-camel milk delivers a safe, alternative milk for human consumption. Acta Trop. 2014, 130, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Walland, J.; Lauper, J.; Frey, J.; Imhof, R.; Stephan, R.; Seuberlich, T.; Oevermann, A. Listeria monocytogenes infection in ruminants: Is there a link to the environment, food and human health? A review. Schweiz. Arch. Tierheilkd. 2015, 157, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, S.; Nucera, D.; Filipello, V. The evolution and epidemiology of Listeria monocytogenes in Europe and the United States. Infect. Genet. Evol. 2015, 35, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Şanlıbaba, P.; Tezel, B.U.; Çakmak, G.A. Prevalence and Antibiotic Resistance of Listeria monocytogenes Isolated from Ready-to-Eat Foods in Turkey. J. Food Quality 2018, 2018, 7693782. [Google Scholar] [CrossRef]
- CDC. Centers for Disease Control and Prevention, Listeria (Listeriosis) | Listeria | National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Foodborne, Waterborne, and Environmental Diseases (DFWED) Page last reviewed: December 18, 2018.
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSAJ 2018, 16, e05500. [Google Scholar]
- Gerard, A.; El-Hajjaji, S.; Niyonzima, E.; Daube, G.; Sindic, M. Prevalence and survival of Listeria monocytogenes in various types of cheese—A review. Int. J. Dairy Technol. 2018, 71, 825–843. [Google Scholar] [CrossRef]
- Li, W.; Bai, L.; Fu, P.; Han, H.; Liu, J.; Guo, Y. The Epidemiology of Listeria monocytogenes in China. Foodborne Pathog. Dis. 2018, 15, 459–466. [Google Scholar] [CrossRef]
- Leong, D.; NicAogáin, K.; Luque-Sastre, L.; McManamon, O.; Hunt, K.; Alvarez-Ordóñez, A.; Scollard, J.; Schmalenberger, A.; Fanning, S.; O’Byrne, C.; et al. A 3-year multi-food study of the presence and persistence of Listeria monocytogenes in 54 small food businesses in Ireland. Int. J. Food Microbiol. 2017, 249, 18–26. [Google Scholar] [CrossRef]
- Kurpas, M.; Wieczorek, K.; Osek, J. Ready-to-eat meat products as a source of Listeria monocytogenes. J. Vet. Res. 2018, 61, 49–55. [Google Scholar] [CrossRef]
- Rodrigues, C.S.; de Sá, C.V.G.C.; de Melo, C.B. An overview of Listeria monocytogenes contamination in ready to eat meat, dairy and fishery foods. Ciência Rural 2017, 47, e20160721. [Google Scholar] [CrossRef]
- Indrawattana, N.; Nibaddhasobon, T.; Sookrung, N.; Chongsa-Nguan, M.; Tungtrongchitr, A.; Makino, S.; Tungyong, W.; Chaicumpa, W. Prevalence of Listeria monocytogenes in raw meats marketed in Bangkok and characterization of the isolates by phenotypic and molecular methods. J. Heal. Popul. Nutr. 2011, 29, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cheng, J.; Wu, Q.; Zhang, J.; Chen, Y.; Xue, L.; Lei, T.; Zeng, H.; Wu, S.; Ye, Q.; et al. Occurrence, antibiotic resistance, and population diversity of Listeria monocytogenes Isolated from fresh aquatic products in China. Front. Microbiol. 2018, 9, 2215. [Google Scholar] [CrossRef] [PubMed]
- Jamali, H.; Paydar, M.; Ismail, S.; Looi, C.Y.; Wong, W.F.; Radmehr, B.; Abedini, A. Prevalence, antimicrobial susceptibility and virulotyping of Listeria species and Listeria monocytogenes isolated from open-air fish markets. BMC Microbiol. 2015, 15, 144. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Osek, J. Prevalence, genetic diversity and antimicrobial resistance of Listeria monocytogenes isolated from fresh and smoked fish in Poland. Food Microbiol. 2017, 64, 164–171. [Google Scholar] [CrossRef]
- Obaidat, M.M.; Bani Salman, A.E.; Lafi, S.Q.; Al-Abboodi, A.R. Characterization of Listeria monocytogenesfrom three countries and antibiotic resistance differencesamong countries and Listeria monocytogenes serogroups. Lett. Appl. Microbiol. 2015, 60, 609–614. [Google Scholar] [CrossRef]
- Skowron, K.; Kwiecińska-Piróg, J.; Grudlewska, K.; Świeca, A.; Paluszak, Z.; Bauza-Kaszewska, J.; Wałecka-Zacharska, E.; Gospodarek-Komkowska, E.; Gospodarek-Komkowska, E. The occurrence, transmission, virulence and antibiotic resistance of Listeria monocytogenes in fish processing plant. Int. J. Food Microbiol. 2018, 282, 71–83. [Google Scholar] [CrossRef]
- Schjørring, S.; Gillesberg Lassen, S.; Jensen, T.; Moura, A.; Kjeldgaard, J.S.; Müller, L.; Thielke, S.; Leclercq, A.; Maury, M.M.; Tourdjman, M.; et al. Cross-border outbreak of listeriosis caused by cold-smoked salmon, revealed by integrated surveillance and whole genome sequencing (WGS), Denmark and France, 2015 to 2017. Eur. Surveill. 2017, 22, pii:17-00762. [Google Scholar] [CrossRef]
- Gillesberg Lassen, S.; Ethelberg, S.; Björkman, J.T.; Jensen, T.; Sørensen, G.; Kvistholm Jensen, A.; Müller, L.; Nielsen, E.M.; Mølbak, K. Two listeria outbreaks caused by smoked fish consumptionusingwhole-genome sequencing for outbreak investigations. Clin. Microbiol. Infect. 2016, 22, 620e624. [Google Scholar] [CrossRef]
- Bae, D.; Mezal, E.; Smiley, R.; Cheng, C.; Khan, A. The sub-species characterization and antimicrobialresistance of Listeria monocytogenes isolated from domesticand imported food products from 2004 to 2011. Food Res. Int. 2014, 64, 656–663. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Domínguez-Bernal, G.; González-Zorn, B.; Kreft, J.; Goebel, W. Pathogenicity islands and virulence evolution in Listeria. Microb. Infect. 2001, 3, 571–584. [Google Scholar] [CrossRef]
- Soni, D.K.; Dubey, S.K. Phylogenetic analysis of the Listeria monocytogenes based on sequencing of 16S rRNA and hlyA genes. Mol. Biol. Rep. 2014, 41, 8219–8229. [Google Scholar] [CrossRef] [PubMed]
- Al-Nabulsi, A.; Osaili, T.M.; Awad, A.A.; Olaimat, A.N.; Shaker, R.R.; Holley, R.A. Occurrence and antibiotic susceptibility of Listeria monocytogenes isolated from raw and processed meat products in Amman, Jordan. CyTA J. Food 2014, 13, 1–7. [Google Scholar]
- Ndahi, M.D.; Kwaga, J.K.; Bello, M.; Kabir, J.; Umoh, V.J.; Yakubu, S.E.; Nok, A.J. Prevalence and antimicrobial susceptibility of Listeria monocytogenes and methicillin-resistant Staphylococcus aureus strains from raw meat and meat products in Zaria, Nigeria. Lett. Appl. Microbiol. 2014, 58, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.M.; Tyler, S.D.; Ewan, E.P.; Ashton, F.E.; Wang, G.; Rozee, K.R. Detection of genes coding for listeriolysin and Listeria monocytogenes antigen A (ImaA) in Listeria spp. by the polymerase chain reaction. Microb. Pathog. 1992, 12, 79–86. [Google Scholar] [CrossRef]
- Cotter, P.D.; Draper, L.A.; Lawton, E.M.; Daly, K.M.; Groeger, D.S.; Casey, P.G.; Ross, R.P.; Hill, C. Listeriolysin S, a Novel Peptide Haemolysin Associated with a Subset of Lineage I Listeria monocytogenes. PLoS Pathog. 2008, 4, e1000144. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wang, H.; Ma, T.; Han, X.; Yang, Y.; Sun, J.; Chen, Z.; Yu, H.; Hang, Y.; Liu, F.; et al. Flagellar Basal Body Structural Proteins FlhB, FliM, and FliY Are Required for Flagellar-Associated Protein Expression in Listeria monocytogenes. Front. Microbiol. 2018, 9, 208. [Google Scholar] [CrossRef]
- Kumar, S.; Parvathi, A.; George, J.; Krohne, G.; Karunasagar, I.; Karunasagar, I. A study on the effects of some laboratory-derived genetic mutations on biofilm formation by Listeria monocytogenes. World J. Microbiol. Biotechnol. 2009, 25, 527–531. [Google Scholar] [CrossRef]
- Price, R.; Jayeola, V.; Niedermeyer, J.; Parsons, C.; Kathariou, S. The Listeria monocytogenes Key Virulence Determinants hly and prfA are involved in Biofilm Formation and Aggregation but not Colonization of Fresh Produce. Pathogens 2018, 7, 18. [Google Scholar] [CrossRef]
- Piercey, M.J.; Hingston, P.A.; Truelstrup Hansen, L. Genes involved in Listeria monocytogenes biofilm formation at a simulated food processing plant temperature of 15 °C. Int. J. Food Microbiol. 2016, 223, 63–74. [Google Scholar] [CrossRef]
- Ripio, M.T.; Domínguez-Bernal, G.; Lara, M.; Suárez, M.; Vázquez-Boland, J.A. A Gly145Ser substitution in the transcriptional activator PrfA causes constitutive overexpression of virulence factors in Listeria monocytogenes. J. Bacteriol. 1997, 179, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Xayarath, B.; Freitag, N.E. Optimizing the balance between host and environmental survival skills: lessons learned from Listeria monocytogenes. Future Microbiol. 2012, 7, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Xayarath, B.; Volz, K.W.; Smart, J.I.; Freitag, N.E. Probing the role of protein surface charge in the activation of PrfA, the central regulator of listeria monocytogenes pathogenesis. PLoS ONE 2011, 6, e23502. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sheehan, B.; Klarsfeld, A.; Ebright, R.; Cossart, P. A single substitution in the putative helix-turn-helix motif of the pleiotropic activator PrfA attenuates Listeria monocytogenes virulence. Mol. Microbiol. 1996, 20, 785–797. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, H.; Zhu, Y.; Gao, P. Structural insights into glutathione-mediated activation of the master regulator PrfA in Listeria monocytogenes. Protein Cell. 2017, 8, 308–312. [Google Scholar] [CrossRef][Green Version]
- Bierne, H.; Sabet, C.; Personnic, N.; Cossart, P. Internalins: A complex family of leucine-rich repeat-containing proteins in Listeria monocytogenes. Microbes Infect. 2007, 9, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Kim, S.; Lee, S.; Lee, H.; Ha, J.; Lee, J.; Choi, Y.; Choi, K.-H.; Yoon, Y. Prevalence and Genetic Characteristics of Meatborne Listeria monocytogenes Isolates from Livestock Farms in Korea. Korean J. Food Sci. Anim. Res. 2016, 36, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Jespersen, L. Prevalence and characteristics of Listeria monocytogenes isolates in raw milk, heated milkand nunu, a spontaneously fermented milk beverage, in Ghana. Beverages 2018, 4, 40. [Google Scholar] [CrossRef]
- Chen, J.Q.; Healey, S.; Regan, P.; Laksanalamai, P.; Hu, Z. PCR-based methodologies for detection and characterization of Listeria monocytogenes and Listeria ivanovii in foods and environmental sources. Food Sci. Human Wellness 2017, 6, 39–59. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Gunasekaran, P.; Rajendhran, J. Molecular Serotyping and Pathogenic Potential of Listeria monocytogenes isolated from milk and milk products in Tamil Nadu, India. Foodborne Pathog. Dis. 2015, 12, 522–528. [Google Scholar] [CrossRef]
- Mammina, C.; Aleo, A.; Romani, C.; Pellissier, N.; Nicoletti, P.; Pecile, P.; Nastasi, A.; Pontello, M.M. Characterization of Listeria monocytogenes isolates from human listeriosis cases in Italy. J. Clin. Microbiol. 2009, 47, 2925–2930. [Google Scholar] [CrossRef] [PubMed]
- Sant Ana, A.S.; Igarashi, M.C.; Landgraf, M.; Destro, M.T.; Franco, B.D.G.M. Prevalence, populations and pheno- and genotypic characteristics of Listeria monocytogenes isolated from ready-to-eat vegetables marketed in São Paulo, Brazil. Int. J. Food Microbiol. 2012, 155, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jamali, H.; Radmehr, B. Frequency, virulence genes and antimicrobial resistance of Listeria spp. isolated from bovine clinical mastitis. Vet. J. 2013, 198, 541–542. [Google Scholar] [CrossRef] [PubMed]
- Jamali, H.; Thong, K.L. Genotypic characterization and antimicrobial resistance of Listeria monocytogenes from ready-to-eat foods. Food Control 2014, 44, 1–6. [Google Scholar] [CrossRef]
- GelbíčoVá, T.; KaRpíšKoVá, R. Outdoor environment as a source of Listeria monocytogenes in food chain. Czech J. Food Sci. 2012, 30, 83–88. [Google Scholar] [CrossRef]
- Newman, T. Listeria: What you need to know: Medical News Today. MediLexicon. Intl. 27 April 2017. Available online: https://www.medicalnewstoday.com/articles/180370.php (accessed on 12 November 2019).
- Wilson, A.; Gray, J.; Chandry, P.S.; Fox, E.M. Phenotypic and Genotypic Analysis of Antimicrobial Resistance among Listeria monocytogenes Isolated from Australian Food Production Chains. Genes 2018, 9, 80. [Google Scholar] [CrossRef]
- Krawczyk-Balska, A.; Markiewicz, Z. The intrinsic cephalosporin resistome of Listeria monocytogenes in the context of stress response, gene regulation, pathogenesis and therapeutics. J. Appl. Microbiol. 2016, 120, 251–265. [Google Scholar] [CrossRef]
- Bertsch, D.; Muelli, M.; Weller, M.; Uruty, A.; Lacroix, C.; Meile, L. Antimicrobial susceptibility and antibiotic resistance gene transfer analysis of foodborne, clinical, and environmental Listeria spp. isolates including Listeria monocytogenes. Microbiol. Open 2014, 3, 118–127. [Google Scholar] [CrossRef]
- Haubert, L.; Mendonça, M.; Lopes, G.V.; de Itapema Cardoso, M.R.; da Silva, W.P. Listeria monocytogenes isolates from food and food environment harbouring tetM and ermB resistance genes. Lett. Appl. Microbiol. 2016, 62, 23–29. [Google Scholar] [CrossRef]
- Li, L.; Olsen, R.H.; Shi, L.; Ye, L.; He, J.; Meng, H. Characterization of a plasmid carrying cat, ermB and tetS genes in a foodborne Listeria monocytogenes strain and uptake of the plasmid by cariogenic Streptococcus mutans. Int. J. Food Microbiol. 2016, 238, 68–71. [Google Scholar] [CrossRef]
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courvalin, P.; Courtieu, A.L. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Zeng, H.; Xue, L.; Lei, T.; Pang, R.; Wu, S.; Wu, H.; et al. Isolation, potential virulence, and population diversity of listeria monocytogenes from meat and meat products in China. Front. Microbiol. 2019, 10, 946. [Google Scholar] [CrossRef] [PubMed]
- Clinical, and Laboratory Standards Institute [CLSI]. Performance Standards for Antimicrobial Susceptibility Testing: 24th Informational Supplement (M100S24); Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.r-project.org/ (accessed on 12 November 2019).
- Zhao, X.; Sandelin, A. GMD: Measuring the distance between histograms with applications on high-throughput sequencing reads. Bioinformatics 2012, 28, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Simko, V. The Corrplot Package. CRAN Repos. 2016. Available online: http://www.sthda.com/french/wiki/matrice-de-correlation-la-fonction-r-qui-fait-tout (accessed on 12 November 2019).
- Benjamini, Y.; Hochberg, Y.; Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the Results of Multivariate Data Analyses. In R Packag Version 1; 2016; Available online: https://rpkgs.datanovia.com/factoextra/index.html. (accessed on 12 November 2019).
- Dixon, P. Vegan, a package for R funcitons for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bryant, D.; Moulton, V. Neighbor-net: An agglomerative method for the construction of phylogenetic networks. Mol. Biol. Evol. 2004, 21, 255–265. [Google Scholar] [CrossRef]
- Huson, D.H. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Genotype | Biofilm Formation | PrfA Mutations | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Isolated L. monocytogenes | hlyA | flaA | inlA | inlB | inlC | inlJ | CT | MPA (O.D.) | E101K | K130I | G145S | G161D | S184P | Accession Numbers |
| Cow milk | + | + | + | + | + | + | Strong | 0.12 | + | - | + | - | + | KP271933 |
| Cow milk | + | + | + | + | + | + | Strong | 0.12 | - | - | + | - | + | KP271934 |
| Buffalo milk | + | + | + | + | + | + | Moderate | 0.15 | - | - | + | + | - | KP271935 |
| She-camel milk | + | + | + | + | + | + | Strong | 0.56 | - | - | + | - | + | KP271936 |
| Ewe milk | + | + | + | + | + | ND | Strong | 0.11 | - | + | + | - | - | KP271937 |
| Goat milk | + | + | + | + | + | ND | Moderate | 0.2 | - | + | + | - | - | KP271938 |
| Goat milk | + | + | + | + | + | + | Strong | 0.2 | - | - | - | - | - | KP271939 |
| Kariesh cheese | + | + | ND | + | + | + | Strong | 0.16 | - | - | - | - | - | KP271940 |
| Hamburger | + | + | ND | + | + | + | Strong | 0.56 | - | - | + | - | + | KP271941 |
| Broilers | + | ND | ND | ND | ND | + | Strong | 0.12 | - | - | + | - | + | KP271942 |
| Broilers | ND | + | ND | ND | ND | ND | Strong | 0.11 | - | - | - | - | - | KP271943 |
| Broilers | + | + | + | + | ND | + | Strong | 0.11 | - | - | - | - | - | KP271944 |
| Layers | + | + | + | + | ND | + | Moderate | 0.2 | - | + | + | - | - | KP271945 |
| Layers | + | + | + | + | ND | ND | Strong | 0.1 | - | + | + | - | - | KP271946 |
| Layers | + | + | ND | ND | ND | ND | Strong | 0.12 | - | - | + | - | + | KP271947 |
| Table eggs | + | + | + | + | + | ND | Moderate | 0.15 | - | - | + | - | + | KP271948 |
| Duck | + | + | + | + | + | + | Moderate | 0.15 | - | - | + | - | + | KX906914 |
| Silage | + | + | + | + | + | + | Strong | 0.1 | - | - | + | - | + | KX906909 |
| Silage | + | ND | + | + | + | + | Strong | 0.1 | - | - | - | - | - | KX906910 |
| Silage | + | ND | + | + | + | + | Strong | 0.1 | - | - | - | - | - | KX906911 |
| Goat fetal liver | ND | ND | + | + | + | + | Strong | 0.12 | - | - | + | - | + | KX906913 |
| Ewe blood (Septicemia) | + | + | + | + | + | + | Strong | 0.12 | - | - | + | - | + | KX906912 |
| Woman blood (Septicemia) | + | + | + | + | + | + | Strong | 0.12 | - | - | + | - | + | KX906908 |
| Frozen fish | + | + | ND | ND | ND | + | Strong | 0.1 | - | - | + | - | + | KX906905 |
| Frozen fish | + | + | + | + | ND | ND | Strong | 0.12 | - | - | + | - | + | KX906906 |
| Herring | + | + | + | + | + | ND | Moderate | 0.15 | - | - | + | - | - | KX906907 |
| Rabbit (brain) | + | + | ND | ND | + | ND | Strong | 0.1 | - | - | + | - | + | KX906915 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, K.M.; Kappell, A.D.; Fox, E.M.; Orabi, A.; Samir, A. Prevalence, Pathogenicity, Virulence, Antibiotic Resistance, and Phylogenetic Analysis of Biofilm-Producing Listeria monocytogenes Isolated from Different Ecological Niches in Egypt: Food, Humans, Animals, and Environment. Pathogens 2020, 9, 5. https://doi.org/10.3390/pathogens9010005
Osman KM, Kappell AD, Fox EM, Orabi A, Samir A. Prevalence, Pathogenicity, Virulence, Antibiotic Resistance, and Phylogenetic Analysis of Biofilm-Producing Listeria monocytogenes Isolated from Different Ecological Niches in Egypt: Food, Humans, Animals, and Environment. Pathogens. 2020; 9(1):5. https://doi.org/10.3390/pathogens9010005
Chicago/Turabian StyleOsman, Kamelia M., Anthony D. Kappell, Edward M. Fox, Ahmed Orabi, and Ahmed Samir. 2020. "Prevalence, Pathogenicity, Virulence, Antibiotic Resistance, and Phylogenetic Analysis of Biofilm-Producing Listeria monocytogenes Isolated from Different Ecological Niches in Egypt: Food, Humans, Animals, and Environment" Pathogens 9, no. 1: 5. https://doi.org/10.3390/pathogens9010005
APA StyleOsman, K. M., Kappell, A. D., Fox, E. M., Orabi, A., & Samir, A. (2020). Prevalence, Pathogenicity, Virulence, Antibiotic Resistance, and Phylogenetic Analysis of Biofilm-Producing Listeria monocytogenes Isolated from Different Ecological Niches in Egypt: Food, Humans, Animals, and Environment. Pathogens, 9(1), 5. https://doi.org/10.3390/pathogens9010005