
Dual Antimicrobial Effect of Medium-Chain Fatty Acids against an Italian Multidrug Resistant Brachyspira hyodysenteriae Strain


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Chemicals and Test Solutions
2.3. Antimicrobial Susceptibility Testing
2.3.1. Agar Dilution Method
2.3.2. Broth Dilution Method
2.4. Virulence Gene Expression Study
2.5. Statistical Analysis
3. Results
3.1. Antimicrobial Susceptibility Testing
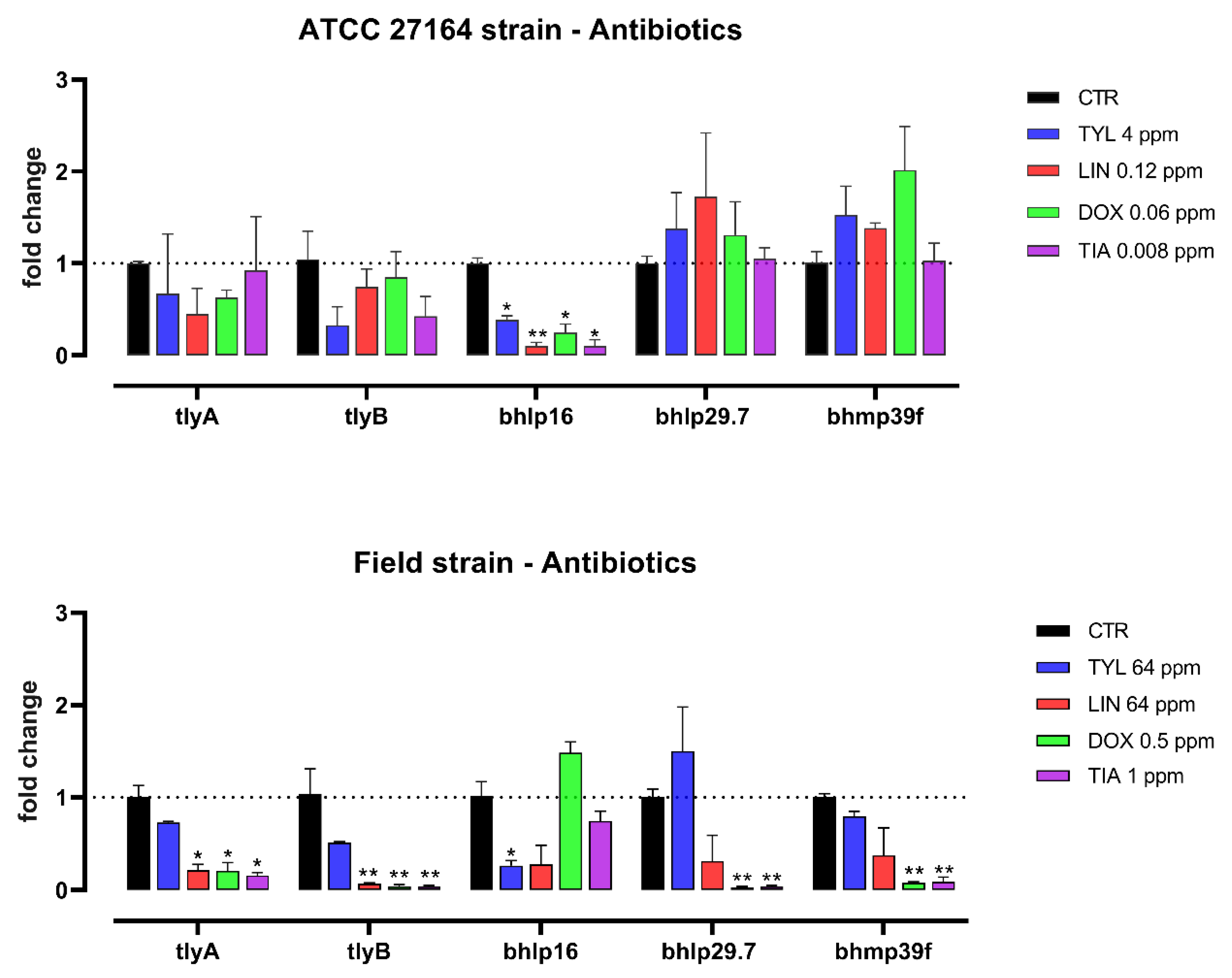
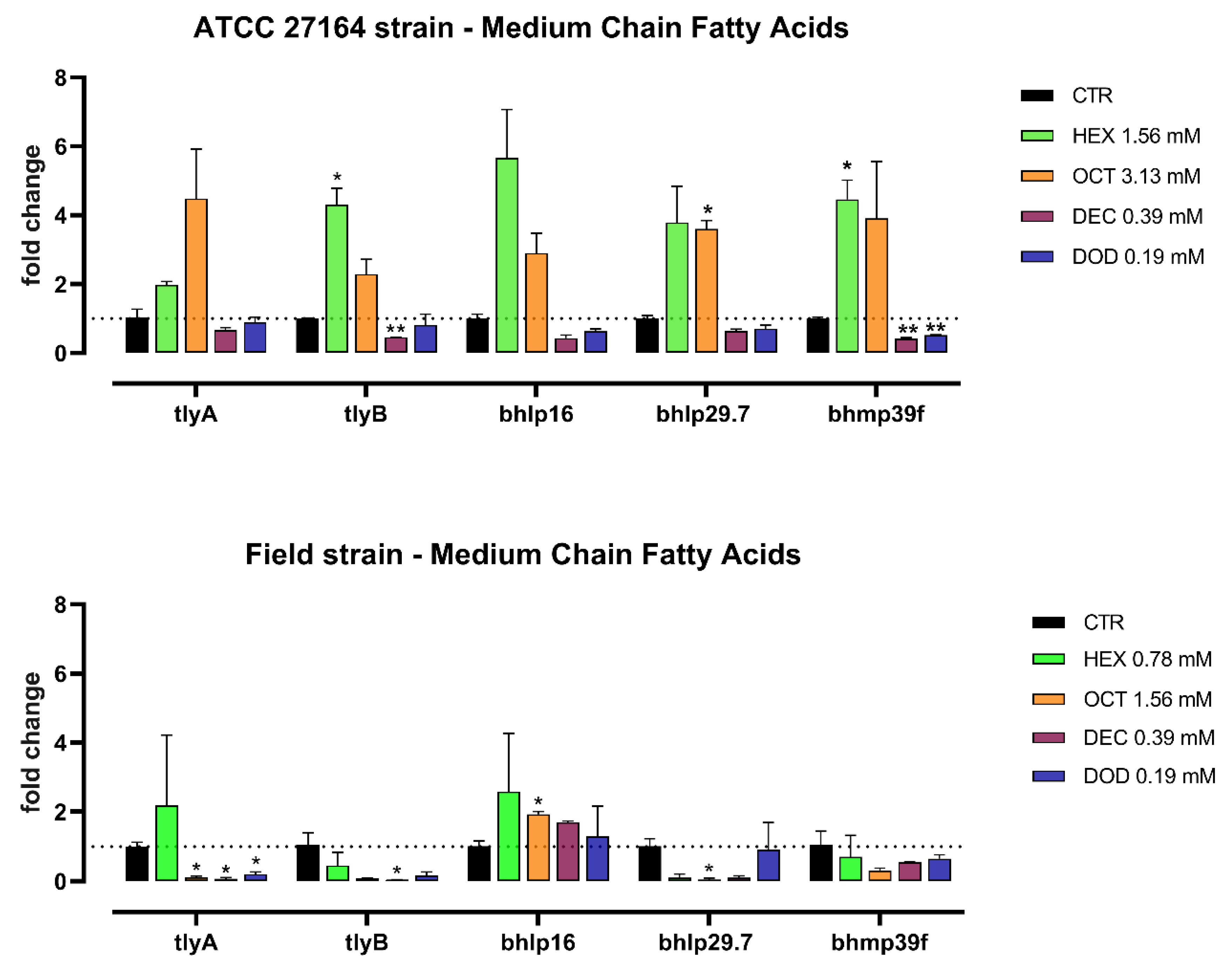
3.2. Gene Expression Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hampson, D.J.; Burrough, E.R. Swine Dysentery and Brachyspiral Colitis. In Diseases of Swine; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 951–970. ISBN 978-1-119-35092-7. [Google Scholar]
- Mahu, M.; De Pauw, N.; Vande Maele, L.; Verlinden, M.; Boyen, F.; Ducatelle, R.; Haesebrouck, F.; Martel, A.; Pasmans, F. Variation in Hemolytic Activity of Brachyspira Hyodysenteriae Strains from Pigs. Vet. Res. 2016, 47, 66. [Google Scholar] [CrossRef] [Green Version]
- Joerling, J.; Willems, H.; Ewers, C.; Herbst, W. Differential Expression of Hemolysin Genes in Weakly and Strongly Hemolytic Brachyspira Hyodysenteriae Strains. BMC Vet. Res. 2020, 16, 169. [Google Scholar] [CrossRef]
- Muir, S.; Koopman, M.B.; Libby, S.J.; Joens, L.A.; Heffron, F.; Kusters, J.G. Cloning and Expression of a Serpula (Treponema) Hyodysenteriae Hemolysin Gene. Infect. Immun. 1992, 60, 529–535. [Google Scholar] [CrossRef] [Green Version]
- ter Huurne, A.A.; Muir, S.; van Houten, M.; van der Zeijst, B.A.; Gaastra, W.; Kusters, J.G. Characterization of Three Putative Serpulina Hyodysenteriae Hemolysins. Microb. Pathog. 1994, 16, 269–282. [Google Scholar] [CrossRef]
- Hsu, T.; Hutto, D.L.; Minion, F.C.; Zuerner, R.L.; Wannemuehler, M.J. Cloning of a Beta-Hemolysin Gene of Brachyspira (Serpulina) Hyodysenteriae and Its Expression in Escherichia Coli. Infect. Immun. 2001, 69, 706–711. [Google Scholar] [CrossRef] [Green Version]
- Trott, D.J.; Alt, D.P.; Zuerner, R.L.; Wannemuehler, M.J.; Stanton, T.B. The Search for Brachyspira Outer Membrane Proteins That Interact with the Host. Anim. Health Res. Rev. 2001, 2, 19–30. [Google Scholar] [CrossRef]
- Hampson, D.J.; Lugsomya, K.; La, T.; Phillips, N.D.; Trott, D.J.; Abraham, S. Antimicrobial Resistance in Brachyspira—An Increasing Problem for Disease Control. Vet. Microbiol. 2019, 229, 59–71. [Google Scholar] [CrossRef]
- Regione Emilia Romagna Linee Guida: Uso Prudente Degli Antibiotici Nell’allevamento Suino 2018. Available online: https://www.alimenti-salute.it/sites/default/files/Linee%20Guida%20SUINI%202018.pdf (accessed on 28 October 2021).
- Desbois, A.P.; Smith, V.J. Antibacterial Free Fatty Acids: Activities, Mechanisms of Action and Biotechnological Potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Tugnoli, B.; Giovagnoni, G.; Piva, A.; Grilli, E. From Acidifiers to Intestinal Health Enhancers: How Organic Acids Can Improve Growth Efficiency of Pigs. Animals 2020, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-Chain Fatty Acids and Monoglycerides as Feed Additives for Pig Production: Towards Gut Health Improvement and Feed Pathogen Mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- CLSI VET06: Methods for Antimicrobial Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria Isolated from Animals 2017. Available online: https://clsi.org/standards/products/veterinary-medicine/documents/vet06/ (accessed on 14 October 2021).
- Giovagnoni, G.; Rossi, B.; Tugnoli, B.; Ghiselli, F.; Bonetti, A.; Piva, A.; Grilli, E. Thymol and Carvacrol Downregulate the Expression of Salmonella Typhimurium Virulence Genes during an In Vitro Infection on Caco-2 Cells. Microorganisms 2020, 8, 862. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods San Diego Calif. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Karlsson, M.; Fellström, C.; Gunnarsson, A.; Landén, A.; Franklin, A. Antimicrobial Susceptibility Testing of Porcine Brachyspira (Serpulina) Species Isolates. J. Clin. Microbiol. 2003, 41, 2596–2604. [Google Scholar] [CrossRef] [Green Version]
- Stubberfield, E.; Pringle, M.; Landén, A.; Veldman, K.T.; Geurts, Y.; Jouy, E.; Le Devendec, L.; Rubin, J.E.; Kulathunga, D.G.R.S.; Kristensen, K.A.; et al. Validation of an Antimicrobial Susceptibility Testing Protocol for Brachyspira Hyodysenteriae and Brachyspira Pilosicoli in an International Ring Trial. Vet. Microbiol. 2020, 244, 108645. [Google Scholar] [CrossRef]
- Rohde, J.; Kessler, M.; Baums, C.G.; Amtsberg, G. Comparison of Methods for Antimicrobial Susceptibility Testing and MIC Values for Pleuromutilin Drugs for Brachyspira Hyodysenteriae Isolated in Germany. Vet. Microbiol. 2004, 102, 25–32. [Google Scholar] [CrossRef]
- Pringle, M.; Aarestrup, F.M.; Bergsjø, B.; Fossi, M.; Jouy, E.; Landén, A.; Mevius, D.; Perry, K.; Teale, C.; Thomson, J.; et al. Quality-Control Ranges for Antimicrobial Susceptibility Testing by Broth Dilution of the Brachyspira Hyodysenteriae Type Strain (ATCC 27164T). Microb. Drug Resist. Larchmt. N 2006, 12, 219–221. [Google Scholar] [CrossRef]
- Mirajkar, N.S.; Gebhart, C.J. Comparison of Agar Dilution and Antibiotic Gradient Strip Test with Broth Microdilution for Susceptibility Testing of Swine Brachyspira Species. J. Vet. Diagn. Investig. 2016, 28, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Ordóez, A.; Martínez-Lobo, F.J.; Arguello, H.; Carvajal, A.; Rubio, P. Swine Dysentery: Aetiology, Pathogenicity, Determinants of Transmission and the Fight against the Disease. Int. J. Environ. Res. Public. Health 2013, 10, 1927–1947. [Google Scholar] [CrossRef] [Green Version]
- Burrough, E.R. Swine Dysentery: Etiopathogenesis and Diagnosis of a Reemerging Disease. Vet. Pathol. 2017, 54, 22–31. [Google Scholar] [CrossRef]
- Pringle, M.; Landén, A.; Unnerstad, H.E.; Molander, B.; Bengtsson, B. Antimicrobial Susceptibility of Porcine Brachyspira Hyodysenteriae and Brachyspira Pilosicoli Isolated in Sweden between 1990 and 2010. Acta Vet. Scand. 2012, 54, 54. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Oxberry, S.L.; Hampson, D.J. Antimicrobial Susceptibility Testing of Australian Isolates of Brachyspira Hyodysenteriae Using a New Broth Dilution Method. Vet. Microbiol. 2002, 84, 123–133. [Google Scholar] [CrossRef]
- Lugsomya, K.; Zeeh, F.; La, T.; Phillips, N.; Hampson, D.J. First Identification and Characterisation of Brachyspira Hyodysenteriae in Pigs in Hong Kong. Porc. Health Manag. 2019, 5, 27. [Google Scholar] [CrossRef]
- Lobová, D.; Smola, J.; Cizek, A. Decreased Susceptibility to Tiamulin and Valnemulin among Czech Isolates of Brachyspira Hyodysenteriae. J. Med. Microbiol. 2004, 53, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Rugna, G.; Bonilauri, P.; Carra, E.; Bergamini, F.; Luppi, A.; Gherpelli, Y.; Magistrali, C.F.; Nigrelli, A.; Alborali, G.L.; Martelli, P.; et al. Sequence Types and Pleuromutilin Susceptibility of Brachyspira Hyodysenteriae Isolates from Italian Pigs with Swine Dysentery: 2003-2012. Vet. J. Lond. Engl. 1997 2015, 203, 115–119. [Google Scholar] [CrossRef] [Green Version]
- De Luca, S.; Nicholson, P.; Magistrali, C.F.; García-Martín, A.B.; Rychener, L.; Zeeh, F.; Frey, J.; Perreten, V. Transposon-Associated Lincosamide Resistance Lnu(C) Gene Identified in Brachyspira Hyodysenteriae ST83. Vet. Microbiol. 2018, 214, 51–55. [Google Scholar] [CrossRef]
- Karlsson, M.; Fellström, C.; Heldtander, M.U.K.; Johansson, K.-E.; Franklin, A. Genetic Basis of Macrolide and Lincosamide Resistance in Brachyspira (Serpulina) Hyodysenteriae. FEMS Microbiol. Lett. 1999, 172, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, Á.; Carvajal, A.; Vester, B.; Pringle, M.; Naharro, G.; Rubio, P. Trends towards Lower Antimicrobial Susceptibility and Characterization of Acquired Resistance among Clinical Isolates of Brachyspira Hyodysenteriae in Spain. Antimicrob. Agents Chemother. 2011, 55, 3330–3337. [Google Scholar] [CrossRef] [Green Version]
- Mahu, M.; Pasmans, F.; Vranckx, K.; De Pauw, N.; Vande Maele, L.; Vyt, P.; Vandersmissen, T.; Martel, A.; Haesebrouck, F.; Boyen, F. Presence and Mechanisms of Acquired Antimicrobial Resistance in Belgian Brachyspira Hyodysenteriae Isolates Belonging to Different Clonal Complexes. Vet. Microbiol. 2017, 207, 125–132. [Google Scholar] [CrossRef]
- Achard, A.; Villers, C.; Pichereau, V.; Leclercq, R. New Lnu(C) Gene Conferring Resistance to Lincomycin by Nucleotidylation in Streptococcus Agalactiae UCN36. Antimicrob. Agents Chemother. 2005, 49, 2716–2719. [Google Scholar] [CrossRef] [Green Version]
- Šperling, D.; Smola, J.; Čížek, A. Characterisation of Multiresistant Brachyspira Hyodysenteriae Isolates from Czech Pig Farms. Vet. Rec. 2011, 168, 215. [Google Scholar] [CrossRef]
- Zmudzki, J.; Szczotka, A.; Nowak, A.; Strzelecka, H.; Grzesiak, A.; Pejsak, Z. Antimicrobial Susceptibility of Brachyspira Hyodysenteriae Isolated from 21 Polish Farms. Pol. J. Vet. Sci. 2012, 15, 259–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchgässner, C.; Schmitt, S.; Borgström, A.; Wittenbrink, M.M. Antimicrobial Susceptibility of Brachyspira Hyodysenteriae in Switzerland. Schweiz. Arch. Tierheilkd. 2016, 158, 405–410. [Google Scholar] [CrossRef] [PubMed]
- García-Martín, A.B.; Perreten, V.; Rossano, A.; Schmitt, S.; Nathues, H.; Zeeh, F. Predominance of a Macrolide-Lincosamide-Resistant Brachyspira Hyodysenteriae of Sequence Type 196 in Swiss Pig Herds. Vet. Microbiol. 2018, 226, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, I.N.; Terzulli, S.; O’Byrne, C. Weak Organic Acids: A Panoply of Effects on Bacteria. Sci. Prog. 2003, 86, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vande Maele, L.; Heyndrickx, M.; Maes, D.; De Pauw, N.; Mahu, M.; Verlinden, M.; Haesebrouck, F.; Martel, A.; Pasmans, F.; Boyen, F. In Vitro Susceptibility of Brachyspira Hyodysenteriae to Organic Acids and Essential Oil Components. J. Vet. Med. Sci. 2016, 78, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.B.; Diez-Gonzalez, F. The Effects of Fermentation Acids on Bacterial Growth. Adv. Microb. Physiol. 1998, 39, 205–234. [Google Scholar] [CrossRef]
- Bonetti, A.; Tugnoli, B.; Rossi, B.; Giovagnoni, G.; Piva, A.; Grilli, E. Nature-Identical Compounds and Organic Acids Reduce E. Coli K88 Growth and Virulence Gene Expression In Vitro. Toxins 2020, 12, 468. [Google Scholar] [CrossRef]
- Heras, B.; Scanlon, M.J.; Martin, J.L. Targeting Virulence Not Viability in the Search for Future Antibacterials. Br. J. Clin. Pharmacol. 2015, 79, 208–215. [Google Scholar] [CrossRef]
- Burrough, E.R.; Strait, E.L.; Kinyon, J.M.; Bower, L.P.; Madson, D.M.; Wilberts, B.L.; Schwartz, K.J.; Frana, T.S.; Songer, J.G. Comparative Virulence of Clinical Brachyspira Spp. Isolates in Inoculated Pigs. J. Vet. Diagn. Investig. Off. Publ. Am. Assoc. Vet. Lab. Diagn. Inc. 2012, 24, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- ter Huurne, A.A.H.M.; van Houten, M.; Muir, S.; Kusters, J.G.; van der Zeijst, B.A.M.; Gaastra, W. Inactivation of a Serpula (Treponema) Hyodysenteriae Hemolysin Gene by Homologous Recombination: Importance of This Hemolysin in Pathogenesis of S. Hyodysenteriae in Mice. FEMS Microbiol. Lett. 1992, 92, 109–113. [Google Scholar] [CrossRef]
- Hyatt, D.R.; ter Huurne, A.A.; van der Zeijst, B.A.; Joens, L.A. Reduced Virulence of Serpulina Hyodysenteriae Hemolysin-Negative Mutants in Pigs and Their Potential to Protect Pigs against Challenge with a Virulent Strain. Infect. Immun. 1994, 62, 2244–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gömmel, M.; Barth, S.; Heydel, C.; Baljer, G.; Herbst, W. Adherence of Brachyspira Hyodysenteriae to Porcine Intestinal Epithelial Cells Is Inhibited by Antibodies against Outer Membrane Proteins. Curr. Microbiol. 2013, 66, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, T.; Gill, R.T. Organic Acid Toxicity, Tolerance, and Production in Escherichia Coli Biorefining Applications. Microb. Cell Fact. 2005, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Piva, A.; Pizzamiglio, V.; Morlacchini, M.; Tedeschi, M.; Piva, G. Lipid Microencapsulation Allows Slow Release of Organic Acids and Natural Identical Flavors along the Swine Intestine. J. Anim. Sci. 2007, 85, 486–493. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Gene | Function | Sequence (5′ → 3′) | Product Length (bp) | Accession Number | Reference |
|---|---|---|---|---|---|
| tlyA | Hemolysin A | F: AAAGGCGTTTGTAGAATTTGGAAT R: TGTCCTACATCAAGAGCATAAACTTTTT | 131 | MT304819.1 | [3] |
| tlyB | Clp protease | F: AAGGATTCGATAAGAAGTATGGTGCTA R: TTCGGTACTCACATAATCCTCTATCTCT | 79 | MT304820.1 | [3] |
| bhlp16 (smpA) | Outer membrane protein | F: GCAGGTGTAGAAAAGGGATTTGG R: TCTGAAGAACTTGCTCCACCTT | 107 | CP015910.2 | This study |
| bhlp29.7 (bmpB) | Outer membrane protein | F: TGGTTTTGCTGGAGAGTCTGA R: TCTCCGTCATTCAAAGCCTGAT | 132 | AY706761.1 | This study |
| bhmp39f | Outer membrane protein | F: AGCCTTTCGGTATTGGCGTA R: ACAGCTATTTGAACAGGAACTGC | 130 | AY027775.1 | This study |
| gyrB | Housekeeping | F: TGCAGGCGGTACTGCTAAAG R: GCACCTACACCGCATCCTAA | 159 | CP015910.2 | This study |
| rpoD | Housekeeping | F: AGCTTTTGCCTCTATCTGACGA R: ACAGTTTGCCGGACAGAGAA | 137 | CP015910.2 | This study |
| ATCC 27164 Strain | Field Strain | ||||
|---|---|---|---|---|---|
| Agar Dilution | Broth Dilution | Agar Dilution | Broth Dilution | ||
| Antibiotics | Tylosin | 16 μg/mL | 8 μg/mL | >64 μg/mL | >64 μg/mL |
| Lincomycin | 2 μg/mL | 0.25 μg/mL | >64 μg/mL | >64 μg/mL | |
| Doxycycline | 2 μg/mL | 0.12 μg/mL | 4 μg/mL | 1 μg/mL | |
| Tiamulin | 0.125 μg/mL | 0.016 μg/mL | 4 μg/mL | 2 μg/mL | |
| MCFA | Hexanoic acid | 25 mM | 3.13 mM | 25 mM | 1.56 mM |
| Octanoic acid | >25 mM | 6.25 mM | 25 mM | 3.13 mM | |
| Decanoic acid | 3.13 mM | 0.78 mM | 3.13 mM | 0.78 mM | |
| Dodecanoic acid | 0.39 mM | 0.39 mM | 0.39 mM | 0.39 mM | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovagnoni, G.; Tugnoli, B.; Piva, A.; Grilli, E. Dual Antimicrobial Effect of Medium-Chain Fatty Acids against an Italian Multidrug Resistant Brachyspira hyodysenteriae Strain. Microorganisms 2022, 10, 301. https://doi.org/10.3390/microorganisms10020301
Giovagnoni G, Tugnoli B, Piva A, Grilli E. Dual Antimicrobial Effect of Medium-Chain Fatty Acids against an Italian Multidrug Resistant Brachyspira hyodysenteriae Strain. Microorganisms. 2022; 10(2):301. https://doi.org/10.3390/microorganisms10020301
Chicago/Turabian StyleGiovagnoni, Giulia, Benedetta Tugnoli, Andrea Piva, and Ester Grilli. 2022. "Dual Antimicrobial Effect of Medium-Chain Fatty Acids against an Italian Multidrug Resistant Brachyspira hyodysenteriae Strain" Microorganisms 10, no. 2: 301. https://doi.org/10.3390/microorganisms10020301
APA StyleGiovagnoni, G., Tugnoli, B., Piva, A., & Grilli, E. (2022). Dual Antimicrobial Effect of Medium-Chain Fatty Acids against an Italian Multidrug Resistant Brachyspira hyodysenteriae Strain. Microorganisms, 10(2), 301. https://doi.org/10.3390/microorganisms10020301