
Molecular Epidemiology and Presence of Hybrid Pathogenic Escherichia coli among Isolates from Community-Acquired Urinary Tract Infection

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Bacterial Isolates
2.2. Phylogenetic Origin
2.3. Virulence Genes Characterization
2.4. Random Amplified Polymorphic DNA (RAPD)
2.5. Cell Culture, Maintenance and Adherence Assay
2.6. Biofilm Assay
2.7. Statistical Analyses
3. Results
3.1. Epidemiological Data on Infections Caused by UPEC Strains
3.2. Classification of ExPEC Virulence Profile, Uropathogenicity, and Phylogenetic Origin of Hybrid UPEC Strains
3.3. Adherence Pattern of Hybrid UPEC Strains
3.4. Interaction with HEK 293T Cells
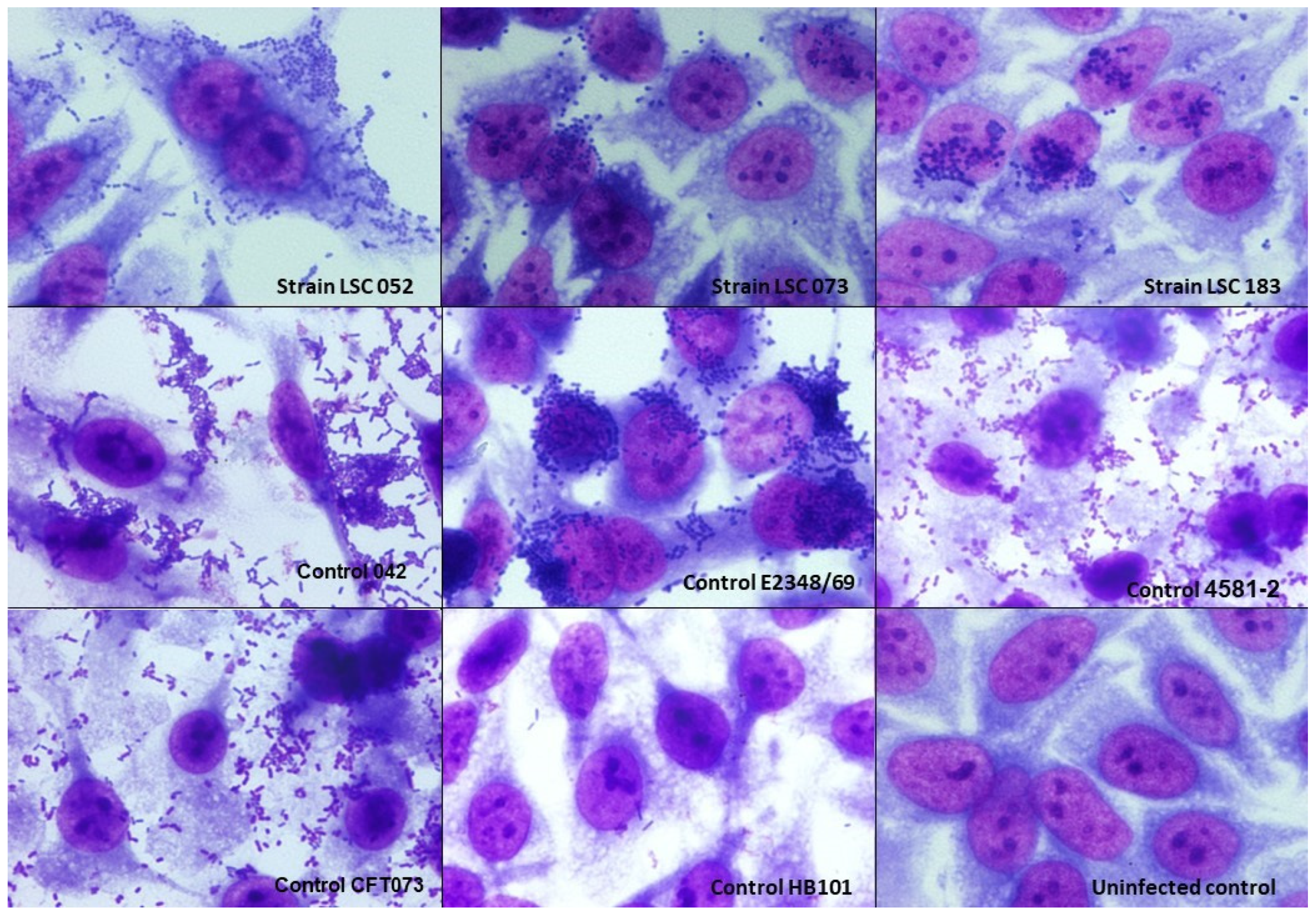
3.5. Interaction with T24 Cells
3.6. Biofilm Formation Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubbs, S.B.; Sommerkamp, S.K. Evaluation and Management of Urinary Tract Infection in the Emergency Department. Emerg. Med. Clin. N. Am. 2019, 37, 707–723. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Foxman, B. The Epidemiology of Urinary Tract Infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef]
- Tamadonfar, K.O.; Omattage, N.S.; Spaulding, C.N.; Hultgren, S.J. Reaching the End of the Line: Urinary Tract Infections. Bact. Intracell. 2020, 7, 83–99. [Google Scholar] [CrossRef]
- Geerlings, S.E. Clinical Presentations and Epidemiology of Urinary Tract Infections. In Urinary Tract Infections: Molecular Pathogenesis and Clinical Management, 2nd ed.; Mulvey, M.A., Klumpp, D.J., Stapleton, A.E., Eds.; ASM Press: Washington, DC, USA, 2017; pp. 27–40. [Google Scholar] [CrossRef]
- Tandogdu, Z.; Wagenlehner, F.M.E. Global Epidemiology of Urinary Tract Infections. Curr. Opin. Infect. Dis. 2016, 29, 73–79. [Google Scholar] [CrossRef]
- Foxman, B. Urinary Tract Infection Syndromes. Occurrence, Recurrence, Bacteriology, Risk Factors, and Disease Burden. Infect. Dis. Clin. N. Am. 2014, 28, 1–13. [Google Scholar] [CrossRef]
- Leimbach, A.; Hacker, J.; Dobrindt, U. E. coli as an All-Rounder: The Thin Line Between Commensalism and Pathogenicity. Curr. Top. Microbiol. Immunol. 2013, 358, 3–32. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Commensal Bacteria in the Gut: Learning Who Our Friends Are. Curr. Opin. Gastroenterol. 2004, 20, 565–571. [Google Scholar] [CrossRef]
- Martinson, J.N.V.; Walk, S.T. Escherichia coli Residency in the Gut of Healthy Human Adults. EcoSal Plus 2020, 9, 10. [Google Scholar] [CrossRef]
- Croxen, M.A.; Finlay, B.B. Molecular Mechanisms of Escherichia coli Pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli. EcoSal Plus 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Dobrindt, U.; Chowdary, M.G.; Krumbholz, G.; Hacker, J. Genome Dynamics and Its Impact on Evolution of Escherichia coli. Med. Microbiol. Immunol. 2010, 199, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Braz, S.; Melchior, K.; Moreira, C.G. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. Front. Cell. Infect. Microbiol. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Touchon, M.; Perrin, A.; de Sousa, J.A.M.; Vangchhia, B.; Burn, S.; O’Brien, C.L.; Denamur, E.; Gordon, D.; Rocha, E.P.C. Phylogenetic Background and Habitat Drive the Genetic Diversification of Escherichia coli. PLoS Genet. 2020, 16, e1008866. [Google Scholar] [CrossRef]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The Population Genetics of Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [Green Version]
- Gomes, T.A.T.; Elias, W.P.; Scaletsky, I.C.A.; Guth, B.E.C.; Rodrigues, J.F.; Piazza, R.M.F.; Ferreira, L.C.S.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47, 3–30. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Extraintestinal Pathogenic Escherichia coli: “The Other Bad E. coli”. Clin. Microbiol. Rev. 2002, 139, 155–162. [Google Scholar] [CrossRef]
- Pitout, J.D.D. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, L.W. Pandemic Lineages of Extraintestinal Pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic (Uropathogenic) Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 383–404. [Google Scholar] [CrossRef] [PubMed]
- Subashchandrabose, S.; Mobley, H.L.T. Virulence and Fitness Determinants of Uropathogenic Escherichia coli. Microbiol. Spectr. 2015, 1, 235–261. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.C.M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of Hybrid- and Hetero-Pathogenic Escherichia coli and Their Potential Implication in More Severe Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, J.; Ambikan, A.; Jernberg, C.; Ehricht, R.; Scheutz, F.; Xiong, Y.; Matussek, A. Molecular Characterization and Comparative Genomics of Clinical Hybrid Shiga Toxin-Producing and Enterotoxigenic Escherichia coli (STEC/ETEC) Strains in Sweden. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H.; et al. Heteropathogenic Virulence and Phylogeny Reveal Phased Pathogenic Metamorphosis in Escherichia coli O2:H6. EMBO Mol. Med. 2014, 6, 347–357. [Google Scholar] [CrossRef]
- Lindstedt, B.-A.; Finton, M.D.; Porcellato, D.; Brandal, L.T. High Frequency of Hybrid Escherichia coli Strains with Combined Intestinal Pathogenic Escherichia coli (IPEC) and Extraintestinal Pathogenic Escherichia coli (ExPEC) Virulence Factors Isolated from Human Faecal Samples. BMC Infect. Dis. 2018, 18, 544. [Google Scholar] [CrossRef]
- Toledo, M.; Fontes, C.; Trabulsi, L. EPM-Modificação Do Meio de Rugai e Araujo Para a Realização Simultânea Dos Testes de Produção de Gás a Partir Da Glicose, H2S, Urease e Triptofano Desaminase. Rev. Microbiol. 1982, 13, 309–315. [Google Scholar]
- Toledo, M.; Fontes, C.; Trabulsi, L. MILi-Um Meio Para a Realização Dos Testes de Motilidade, Indol e Lisina Descarboxilase. Rev. Microbiol. 1982, 13, 230–235. [Google Scholar]
- Starr, M.P. Edwards and Ewing’s Identification of Enterobacteriaceae. Int. J. Syst. Bacteriol. 1986, 36, 581–582. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli Phylo-Typing Method Revisited: Improvement of Specificity and Detection of New Phylo-Groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and Molecular Characterization of Nalidixic Acid-Resistant Extraintestinal Pathogenic Escherichia coli from Retail Chicken Products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli Isolates That Carry vat, fyuA, chuA, and yfcV Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, K.L.; Dynesen, P.; Larsen, P.; Frimodt-Møller, N. Faecal Escherichia coli from Patients with E. coli Urinary Tract Infection and Healthy Controls Who Have Never Had a Urinary Tract Infection. J. Med. Microbiol. 2014, 63, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.A.S.; Santos, F.F.; Valiatti, T.B.; Santos-Neto, J.F.; Santos, A.C.M.; Cayô, R.; Gales, A.C.; Gomes, T.A.T. Frequency and Diversity of Hybrid Escherichia coli Strains Isolated from Urinary Tract Infections. Microorganisms 2021, 9, 693. [Google Scholar] [CrossRef]
- Santos, F.F.; Yamamoto, D.; Abe, C.M.; Bryant, J.A.; Hernandes, R.T.; Kitamura, F.C.; Castro, F.S.; Valiatti, T.B.; Piazza, R.M.F.; Elias, W.P.; et al. The Type III Secretion System (T3SS)-Translocon of Atypical Enteropathogenic Escherichia coli (aEPEC) Can Mediate Adherence. Front. Microbiol. 2019, 10, 1527. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, N.; Nishi, J.; Sheikh, J.; Nataro, J.P.; Sarantuya, J.; Iwashita, M.; Manago, K.; Tokuda, K.; Yoshinaga, M.; Kawano, Y. Quantitative Biofilm Assay Using a Microtiter Plate to Screen for Enteroaggregative Escherichia coli. Am. J. Trop. Med. Hyg. 2004, 71, 687–690. [Google Scholar] [CrossRef]
- Heilberg, I.P.; Schor, N. Abordagem diagnóstica e terapêutica na infecção do trato urinário: ITU. Rev. Assoc. Med. Bras. 2003, 49, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-Antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Foxman, B.; Brown, P. Epidemiology of Urinary Tract Infections: Transmission and Risk Factors, Incidence, and Costs. Infect. Dis. Clin. N. Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- Ronald, A. The Etiology of Urinary Tract Infection: Traditional and Emerging Pathogens. Dis. Mon. 2003, 49, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Hooton, T.M. Clinical Practice. Uncomplicated Urinary Tract Infection. N. Engl. J. Med. 2012, 366, 1028–1037. [Google Scholar] [CrossRef]
- Korbel, L.; Howell, M.; Spencer, J.D. The Clinical Diagnosis and Management of Urinary Tract Infections in Children and Adolescents. Paediatr. Int. Child Health 2017, 37, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, A.E. The Vaginal Microbiota and Urinary Tract Infection. Microbiol. Spectr. 2016, 4, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Dielubanza, E.J.; Schaeffer, A.J. Urinary Tract Infections in Women. Med. Clin. N. Am. 2011, 95, 27–41. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Bautista, A.; Reyes, R.; Vicente, A.; Vaca-Paniagua, F.; Díaz, C.E.; Martínez, S.; Domínguez, P.; García, L.R.; et al. Multiple Antibiotic Resistances and Virulence Markers of Uropathogenic Escherichia coli from Mexico. Pathog. Glob. Health. 2018, 112, 415–420. [Google Scholar] [CrossRef]
- Rezatofighi, S.E.; Mirzarazi, M.; Salehi, M. Virulence Genes and Phylogenetic Groups of Uropathogenic Escherichia coli Isolates from Patients with Urinary Tract Infection and Uninfected Control Subjects: A Case-Control Study. BMC Infect. Dis. 2021, 21, 361. [Google Scholar] [CrossRef]
- Lin, W.H.; Wang, M.C.; Liu, P.Y.; Chen, P.S.; Wen, L.L.; Teng, C.H.; Kao, C.Y. Escherichia coli Urinary Tract Infections: Host Age-Related Differences in Bacterial Virulence Factors and Antimicrobial Susceptibility. J. Microbiol. Immunol. Infect. 2021; in press. [Google Scholar] [CrossRef]
- Lara, F.B.M.; Nery, D.R.; de Oliveira, P.M.; Araujo, M.L.; Carvalho, F.R.Q.; Messias-Silva, L.C.F.; Ferreira, L.B.; Faria-Junior, C.; Pereira, A.L. Virulence Markers and Phylogenetic Analysis of Escherichia coli Strains with Hybrid EAEC/UPEC Genotypes Recovered from Sporadic Cases of Extraintestinal Infections. Front. Microbiol. 2017, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Olesen, B.; Scheutz, F.; Andersen, R.L.; Menard, M.; Boisen, N.; Johnston, B.; Hansen, D.S.; Krogfelt, K.A.; Nataro, J.P.; Johnson, J.R. Enteroaggregative Escherichia coli O78:H10, the Cause of an Outbreak of Urinary Tract Infection. J. Clin. Microbiol. 2012, 50, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, R.C.S.; Marangoni, D.V.; Smith, S.P.; Alves, E.M.; Pellegrino, F.L.P.C.; Riley, L.W.; Moreira, B.M. Clonal Composition of Escherichia coli Causing Community-Acquired Urinary Tract Infections in the State of Rio de Janeiro, Brazil. Microb. Drug. Resist. 2009, 15, 303–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiatti, T.B.; Santos, F.F.; Santos, A.C.M.; Nascimento, J.A.S.; Silva, R.M.; Carvalho, E.; Sinigaglia, R.; Gomes, T.A.T. Genetic and Virulence Characteristics of a Hybrid Atypical Enteropathogenic and Uropathogenic Escherichia coli (aEPEC/UPEC) Strain. Front. Cell Infect. Microbiol. 2020, 10, 492. [Google Scholar] [CrossRef] [PubMed]
- Lüthje, P.; Brauner, A. Virulence Factors of Uropathogenic E. coli and Their Interaction with the Host. Adv. Microb. Physiol. 2014, 65, 337–372. [Google Scholar] [CrossRef]
- Zangane Matin, F.; Rezatofighi, S.E.; Roayaei Ardakani, M.; Akhoond, M.R.; Mahmoodi, F. Virulence Characterization and Clonal Analysis of Uropathogenic Escherichia coli Metallo-Beta-Lactamase-Producing Isolates. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 50. [Google Scholar] [CrossRef]
- Zeng, Q.; Xiao, S.; Gu, F.; He, W.; Xie, Q.; Yu, F.; Han, L. Antimicrobial Resistance and Molecular Epidemiology of Uropathogenic Escherichia coli Isolated From Female Patients in Shanghai, China. Front. Cell. Infect. Microbiol. 2021, 11, 751. [Google Scholar] [CrossRef]
- Johnson, J.R. Virulence Factors in Escherichia coli Urinary Tract Infection. Clin. Microbiol. Rev. 1991, 4, 80–128. [Google Scholar] [CrossRef]
- O’Brien, V.P.; Hannan, T.J.; Yu, L.; Livny, J.; Roberson, E.D.O.; Schwartz, D.J.; Souza, S.; Mendelsohn, C.L.; Colonna, M.; Lewis, A.L.; et al. A Mucosal Imprint Left by Prior Escherichia coli Bladder Infection Sensitizes to Recurrent Disease. Nat. Microbiol. 2016, 2, 16196. [Google Scholar] [CrossRef]
- Warren, J.W.; Mobley, H.L.T.; Trifillis, A.L. Internalization of Escherichia coli into Human Renal Tubular Epithelial Cells. J. Infect. Dis. 1988, 158, 221–223. [Google Scholar] [CrossRef]
- Otto, G.; Magnusson, M.; Svensson, M.; Braconier, J.; Svanborg, C. pap Genotype and P Fimbrial Expression in Escherichia coli Causing Bacteremic and Nonbacteremic Febrile Urinary Tract Infection. Clin. Infect. Dis 2001, 32, 1523–1531. [Google Scholar] [CrossRef] [Green Version]
- Bert, F.; Johnson, J.R.; Ouattara, B.; Leflon-Guibout, V.; Johnston, B.; Marcon, E.; Valla, D.; Moreau, R.; Nicolas-Chanoine, M.H. Genetic Diversity and Virulence Profiles of Escherichia coli Isolates Causing Spontaneous Bacterial Peritonitis and Bacteremia in Patients with Cirrhosis. J. Clin. Microbiol. 2010, 48, 2709–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olesen, B.; Kolmos, H.J.; Ørskov, F.; Ørskov, I. Cluster of Multiresistant Escherichia coli 078:H10 in Greater Copenhagen. Scand. J. Infect. Dis 1994, 26, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R.; Mende, K.; Murray, C.K.; Johnston, B.D.; Sokurenko, E.V.; Tchesnokova, V.; Johnson, J.R. Clonal Distribution and Associated Characteristics of Escherichia coli Clinical and Surveillance Isolates from a Military Medical Center. Diagn. Microbiol. Infect. Dis. 2017, 87, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, A.C.C.; Andrade, N.L.; Ferdous, M.; Chlebowicz, M.A.; Santos, C.C.; Correal, J.C.D.; lo Ten Foe, J.R.; Rosa, A.C.P.; Damasco, P.V.; Friedrich, A.W.; et al. Comprehensive Molecular Characterization of Escherichia coli Isolates from Urine Samples of Hospitalized Patients in Rio de Janeiro, Brazil. Front. Microbiol. 2018, 9, 243. [Google Scholar] [CrossRef]
- Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Nishiyama, M.Y.; et al. Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection. Microorganisms 2020, 8, 827. [Google Scholar] [CrossRef]
- Nüesch-Inderbinen, M.T.; Baschera, M.; Zurfluh, K.; Hächler, H.; Nüesch, H.; Stephan, R. Clonal Diversity, Virulence Potential and Antimicrobial Resistance of Escherichia coli Causing Community Acquired Urinary Tract Infection in Switzerland. Front. Microbiol. 2017, 8, 2334. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Johnston, B.D.; Porter, S.; Thuras, P.; Aziz, M.; Price, L.B. Accessory Traits and Phylogenetic Background Predict Escherichia coli Extraintestinal Virulence Better Than Does Ecological Source. J. Infect. Dis. 2019, 219, 121–132. [Google Scholar] [CrossRef]
- Flament-Simon, S.C.; Nicolas-Chanoine, M.H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Merino, I.; Porter, S.B.; Johnston, B.; Clabots, C.; Thuras, P.; Ruiz-Garbajosa, P.; Cantón, R.; Johnson, J.R. Molecularly defined extraintestinal pathogenic Escherichia coli status predicts virulence in a murine sepsis model better than does virotype, individual virulence genes, or clonal subset among E. coli ST131 isolates. Virulence 2020, 11, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Porter, S.; Johnston, B.; Kuskowski, M.A.; Spurbeck, R.R.; Mobley, H.L.; Williamson, D.A. Host Characteristics and Bacterial Traits Predict Experimental Virulence for Escherichia coli Bloodstream Isolates From Patients With Urosepsis. Open Forum Infect. Dis. 2015, 2, 1–9. [Google Scholar] [CrossRef]
- Mamani, R.; Flament-Simon, S.C.; García, V.; Mora, A.; Alonso, M.P.; López, C.; García-Meniño, I.; Díaz-Jiménez, D.; Blanco, J.E.; Blanco, M.; et al. Sequence Types, Clonotypes, Serotypes, and Virotypes of Extended-Spectrum β-Lactamase-Producing Escherichia coli Causing Bacteraemia in a Spanish Hospital Over a 12-Year Period (2000 to 2011). Front. Microbiol. 2019, 10, 1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fröding, I.; Hasan, B.; Sylvin, I.; Coorens, M.; Nauclér, P.; Giske, C.G. Extended-Spectrum-β-Lactamase- and Plasmid AmpC-Producing Escherichia coli Causing Community-Onset Bloodstream Infection: Association of Bacterial Clones and Virulence Genes with Septic Shock, Source of Infection, and Recurrence. Antimicrob. Agents Chemother. 2020, 64, e02351-19. [Google Scholar] [CrossRef] [PubMed]
- Toval, F.; Köhler, C.D.; Vogel, U.; Wagenlehner, F.; Mellmann, A.; Fruth, A.; Schmidt, M.A.; Karch, H.; Bielaszewska, M.; Dobrindt, U. Characterization of Escherichia coli Isolates from Hospital Inpatients or Outpatients with Urinary Tract Infection. J. Clin. Microbiol. 2014, 52, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, C.A.; Santos, A.C.M.; Pignatari, A.C.; Silva, R.M.; Elias, W.P. Serine Protease Autotransporters of Enterobacteriaceae (SPATEs) Are Largely Distributed among Escherichia coli Isolated from the Bloodstream. Braz. J. Microbiol. 2020, 51, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Tapader, R.; Basu, S.; Pal, A. Secreted Proteases: A New Insight in the Pathogenesis of Extraintestinal Pathogenic Escherichia coli. Int. J. Med. Microbiol. 2019, 309, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Modgil, V.; Kaur, H.; Mohan, B.; Taneja, N. Molecular, phylogenetic and antibiotic resistance analysis of enteroaggregative Escherichia coli/uropathogenic Escherichia coli hybrid genotypes causing urinary tract infections. Indian J. Med. Microbiol. 2020, 38, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Mandomando, I.; Vubil, D.; Boisen, N.; Quintó, L.; Ruiz, J.; Sigaúque, B.; Nhampossa, T.; Garrine, M.; Massora, S.; Aide, P.; et al. Escherichia coli ST131 Clones Harbouring AggR and AAF/V Fimbriae Causing Bacteremia in Mozambican Children: Emergence of New Variant of fimH27 Subclone. PLoS Negl. Trop. Dis. 2020, 14, e0008274. [Google Scholar] [CrossRef]
- Cointe, A.; Birgy, A.; Mariani-Kurkdjian, P.; Liguori, S.; Courroux, C.; Blanco, J.; Delannoy, S.; Fach, P.; Loukiadis, E.; Bidet, P.; et al. Emerging Multidrug-Resistant Hybrid Pathotype Shiga Toxin–Producing Escherichia coli O80 and Related Strains of Clonal Complex 165, Europe. Emerg. Infect. Dis. 2018, 24, 2262–2269. [Google Scholar] [CrossRef] [Green Version]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and evolution of hybrid Shiga toxin-producing and uropathogenic (STEC/UPEC) Escherichia coli of sequence type 141. J. Clin. Microbiol. 2019, 58, 1309–1319. [Google Scholar] [CrossRef]
- Mariani-Kurkdjian, P.; Lemaître, C.; Bidet, P.; Perez, D.; Boggini, L.; Kwon, T.; Bonacorsi, S. Haemolytic-Uraemic Syndrome with Bacteraemia Caused by a New Hybrid Escherichia coli Pathotype. New Microbes New Infect 2014, 2, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Munhoz, D.D.; Santos, F.F.; Mitsunari, T.; Schüroff, P.A.; Elias, W.P.; Carvalho, E.; Piazza, R.M.F. Hybrid Atypical Enteropathogenic and Extraintestinal Escherichia coli (aEPEC/ExPEC) BA1250 Strain: A Draft Genome. Pathogens 2021, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Riveros, M.; García, W.; García, C.; Durand, D.; Mercado, E.; Ruiz, J.; Ochoa, T.J. Molecular and Phenotypic Characterization of Diarrheagenic Escherichia coli Strains Isolated from Bacteremic Children. Am. J. Trop. Med. Hyg. 2017, 97, 1329–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipour, T.; Poursina, F. The Frequency of Hybrid Enteroaggregative/Uropathogenic Escherichia coli Isolated from Clinical Samples of Isfahan Hospitals, Iran. Gene Rep. 2021, 23, 101042. [Google Scholar] [CrossRef]
- Abe, C.M.; Salvador, F.A.; Falsetti, I.N.; Vieira, M.A.M.; Blanco, J.; Blanco, J.E.; Blanco, M.; Machado, A.M.O.; Elias, W.P.; Hernandes, R.T.; et al. Uropathogenic Escherichia coli (UPEC) Strains May Carry Virulence Properties of Diarrhoeagenic E. coli. FEMS Microbiol. Immunol. 2008, 52, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Santos, V.I.; Ruíz-Rosas, M.; Ramirez-Peralta, A.; García, O.Z.; Resendiz-Reyes, L.A.; Romero-Pineda, O.J.; Castro-Alarcón, N. Enteroaggregative Escherichia coli Is Associated with Antibiotic Resistance and Urinary Tract Infection Symptomatology. PeerJ 2021, 9, e11726. [Google Scholar] [CrossRef] [PubMed]
- Gati, N.S.; Temme, I.J.; Middendorf-Bauchart, B.; Kehl, A.; Dobrindt, U.; Mellmann, A. Comparative Phenotypic Characterization of Hybrid Shiga Toxin-Producing/Uropathogenic Escherichia coli, Canonical Uropathogenic and Shiga Toxin-Producing Escherichia coli. Int. J. Med. Microbiol. 2021, 311, 151533. [Google Scholar] [CrossRef]
- Høiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.J.; Moser, C.; Jensen, P.Ø.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The Clinical Impact of Bacterial Biofilms. Int. J. Oral Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Watts, R.E.; Hancock, V.; Ong, C.L.Y.; Vejborg, R.M.; Mabbett, A.N.; Totsika, M.; Looke, D.F.; Nimmo, G.R.; Klemm, P.; Schembri, M.A. Escherichia coli Isolates Causing Asymptomatic Bacteriuria in Catheterized and Noncatheterized Individuals Possess Similar Virulence Properties. J. Clin. Microbiol. 2010, 48, 2449–2458. [Google Scholar] [CrossRef] [Green Version]
- Novais, Â.; Pires, J.; Ferreira, H.; Costa, L.; Montenegro, C.; Vuotto, C.; Donelli, G.; Coque, T.M.; Peixe, L. Characterization of Globally Spread Escherichia coli ST131 Isolates (1991 to 2010). Antimicrob. Agents Chemother. 2012, 56, 3973–3976. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, P.; Natarajan, V.; Sevanan, M. In Vitro Biofilm Formation by Uropathogenic Escherichia coli and Their Antimicrobial Susceptibility Pattern. Asian Pac. J. Trop. Med. 2012, 5, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, J.; Mishra, B.; Srivastava, S.; Srivastava, R. Genotypic Characteristics and Biofilm Formation among Escherichia coli Isolates from Indian Women with Acute Cystitis. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Tapiainen, T.; Hanni, A.M.; Salo, J.; Ikäheimo, I.; Uhari, M. Escherichia coli Biofilm Formation and Recurrences of Urinary Tract Infections in Children. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Le Bouguenec, C.; Archambaud, M.; Labigne, A. Rapid and specific detection of the pap, afa, and sfa adhesin-encoding operons in uropathogenic Escherichia coli strains by polymerase chain reaction. J. Clin. Microbiol. 1992, 5, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalioui, L.; Le Bouguénec, C. afa-8 Gene cluster is carried by a pathogenicity island inserted into the tRNA(Phe) of human and bovine pathogenic Escherichia coli isolates. Infect. Immun. 2001, 69, 937–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, D.; Greune, L.; Heusipp, G.; Karch, H.; Fruth, A.; Tschäpe, H.; Schmidt, M.A. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl. Environ. Microbiol. 2007, 73, 3380–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, J.; Kurazono, H.; Takeda, Y. Distribution of the cytolethal distending toxin A gene (cdtA) among species of Shigella and Vibrio, and cloning and sequencing of the cdt gene from Shigella dysenteriae. Microb. Pathog. 1995, 18, 167–172. [Google Scholar] [CrossRef]
- Santos, A.C.; Zidko, A.C.; Pignatari, A.C.; Silva, R.M. Assessing the diversity of the virulence potential of Escherichia coli isolated from bacteremia in São Paulo, Brazil. Braz. J. Med. Biol. Res. 2013, 46, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Vidal, M.; Kruger, E.; Durán, C.; Lagos, R.; Levine, M.; Prado, V.; Toro, C.; Vidal, R. Single multiplex PCR assay to identify simultaneously the six categories of diarrheagenic Escherichia coli associated with enteric infections. J. Clin. Microbiol. 2005, 43, 5362–5365. [Google Scholar] [CrossRef] [Green Version]
- Gannon, V.P.; Rashed, M.; King, R.K.; Thomas, E.J. Detection and characterization of the eae gene of Shiga-like toxin-producing Escherichia coli using polymerase chain reaction. J. Clin. Microbiol. 1993, 31, 1268–1274. [Google Scholar] [CrossRef] [Green Version]
- Marc, D.; Dho-Moulin, M. Analysis of the fim cluster of an avian O2 strain of Escherichia coli: Serogroup-specific sites within fimA and nucleotide sequence of fimI. J. Med. Microbiol. 1996, 44, 444–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Terai, A.; Yuri, K.; Kurazono, H.; Takeda, Y.; Yoshida, O. Detection of urovirulence factors in Escherichia coli by multiplex polymerase chain reaction. FEMS Immunol. Med. Microbiol. 1995, 12, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antáo, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef]
- Szalo, I.M.; Goffaux, F.; Pirson, V.; Piérard, D.; Ball, H.; Mainil, J. Presence in bovine enteropathogenic (EPEC) and enterohaemorrhagic (EHEC) Escherichia coli of genes encoding for putative adhesins of human EHEC strains. Res. Microbiol. 2002, 153, 653–658. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A.; Tarr, P.I.; Carlino, U.; Bilge, S.S.; Vary, J.C., Jr.; Stell, A.L. Molecular epidemiological and phylogenetic associations of two novel putative virulence genes, iha and iroN (E. coli), among Escherichia coli isolates from patients with urosepsis. Infect. Immun. 2000, 68, 3040–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; O’Bryan, T.T.; Low, D.A.; Ling, G.; Delavari, P.; Fasching, C.; Russo, T.A.; Carlino, U.; Stell, A.L. Evidence of commonality between canine and human extraintestinal pathogenic Escherichia coli strains that express papG allele III. Infect. Immun. 2000, 68, 3327–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioppo, N.M.; Elias, W.P., Jr.; Vidotto, M.C.; Linhares, R.E.; Saridakis, H.O.; Gomes, T.A.; Trabulsi, L.R.; Pelayo, J.S. Prevalence of HEp-2 cell-adherent Escherichia coli and characterisation of enteroaggregative E. coli and chain-like adherent E. coli isolated from children with and without diarrhoea, in Londrina, Brazil. FEMS Microbiol. Lett. 2000, 190, 293–298. [Google Scholar] [CrossRef]
- Paton, A.W.; Paton, J.C. Direct detection and characterization of Shiga toxigenic Escherichia coli by multiplex PCR for stx1, stx2, eae, ehxA, and saa. J. Clin. Microbiol. 2002, 40, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Czeczulin, J.R.; Whittam, T.S.; Henderson, I.R.; Navarro-Garcia, F.; Nataro, J.P. Phylogenetic analysis of enteroaggregative and diffusely adherent Escherichia coli. Infect. Immun. 1999, 67, 2692–2699. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Delicato, E.R.; de Brito, B.G.; Konopatzki, A.P.; Gaziri, L.C.; Vidotto, M.C. Occurrence of the temperature-sensitive hemagglutinin among avian Escherichia coli. Avian Dis. 2002, 46, 713–716. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pathotype | Molecular Markers |
|---|---|
| EAEC | aggR |
| ETEC | elt and/or est |
| EIEC | invE |
| typical EPEC | bfpB, eae, escV |
| atypical EPEC | eae, escV |
| STEC | stx1 and/or stx2 |
| EHEC | stx1 and/or stx2, eae, escV |
| ExPEC+ (intrinsic virulence) a | ≥2 among papC, sfaDE, afaBCIII, iucD, kpsMTII |
| UPEC+ (uropathogenic potential) a | vat, chuA, fyuA, and yfcV |
| All Isolates | ExPEC+ a | UPEC+ b | Non-UPEC+/ExPEC+ | ||
|---|---|---|---|---|---|
| Traits | VFs | n = 172 | n = 59 | n = 31 | n = 101 |
| Adhesins /Invasins | fimA | 163 (94.8) | 57 (96.6) | 29 (93.5) | 95 (94) |
| ompA | 143 (83.1) | 46 (77.9) | 27 (87.1) | 86 (85.1) | |
| yfcV | 72 (41.8) | 36 (61) *** | 31 (100) *** | 24 (23.7) | |
| mat | 72 (41.8) | 30 (50.8) | 13 (41.9) | 37 (36.6) | |
| hra | 61 (35.4) | 32 (54.2) ** | 17 (54.8) * | 26 (25.7) | |
| papC | 48 (27.9) | 40 (67.8) *** | 14 (45.2) *** | 7 (6.9) | |
| saa | 37 (21.5) | 23 (38.9) *** | 12 (38.7) ** | 10 (9.9) | |
| sfaDE | 28 (16.2) | 23 (38.9) *** | 8 (25.8) * | 5 (4.9) | |
| iha | 19 (11) | 7 (11.8) | 4 (12.9) | 10 (9.9) | |
| shf | 14 (8.1) | 2 (3.3) | 2 (6.4) | 10 (9.9) | |
| daaE | 2 (1.1) | 2 (3.3) | 0 (0) | 0 (0) | |
| eae | 2 (1.1) | 0 (0) | 1 (3.2) | 1 (0.9) | |
| tsh | 1 (0.5) | 0 (0) | 1 (3.2) | 0 (0) | |
| Protectins | ompT | 109 (63.3) | 52 (88.1) *** | 28 (90.3) *** | 46 (45.5) |
| kpsMTII | 77 (44.7) | 51 (86.4) *** | 24 (77.4) *** | 20 (19.8) | |
| traT | 72 (41.9) | 34 (57.6) * | 20 (64.5) * | 32 (31.6) | |
| kpsMTIII | 2 (1.1) | 1 (1.6) | 2 (6.4) * | 0 (0) | |
| cvaC | 16 (9.3) | 8 (13.6) | 5 (16.1) | 7 (6.9) | |
| Iron acquisition systems | chuA | 99 (57.6) | 53 (89.8) *** | 31 (100) *** | 34 (33.6) |
| fyuA | 89 (51.7) | 37 (62.7) * | 31 (100) *** | 40 (39.6) | |
| sitA | 88 (51.1) | 35 (59.3) | 20 (64.5) | 46 (45.5) | |
| iroN | 57 (33.1) | 33 (55.9) *** | 21 (67.7) *** | 18 (17.8) | |
| iucD | 41 (23.8) | 29 (49.1) *** | 9 (29) * | 11 (10.9) | |
| ireA | 15 (8.7) | 8 (13.6) * | 4 (12.9) | 4 (3.9) | |
| Toxins | vat | 38 (22) | 19 (32.2) *** | 31 (100) *** | 7 (6.9) |
| sat | 22 (12.7) | 12 (20.3) * | 4 (12.9) | 8 (7.9) | |
| hlyA | 18 (10.4) | 16 (27.1) *** | 5 (16.1) * | 2 (1.9) | |
| pet | 17 (9.9) | 11 (18.6) * | 7 (22.6) ** | 3 (2.9) | |
| pic | 9 (5.2) | 7 (11.8) * | 2 (6.4) | 1 (0.9) | |
| cdtA | 6 (3.5) | 4 (6.7) | 4 (12.9) * | 1 (0.9) | |
| Transcriptional regulator | aggR | 1 (0.5) | 0 (0) | 0 (0) | 1 (0.9) |
| Secretion system | escV | 2 (1.1) | 0 (0) | 1 (3.2) | 1 (0.9) |
| ExPEC+ | 59 (34.3) | N.A. | 19 (61.2) | N.A. | |
| UPEC+ | 31 (18) | 19 (32.2) | N.A. | N.A. | |
| Hybrid pathogenic E. coli c | 3 (1.7) | 0 (0) | 1 (3.2) | 2 (1.9) | |
| Phylogroups | |||||
| A | 27 (15.7) | 0 (0) (***) | 0 (0) (**) | 27 (26.7) | |
| B1 | 21 (12.2) | 2 (3.3) (*) | 0 (0) (*) | 19 (18.8) | |
| B2 | 79 (46) | 49 (83) *** | 30 (96.8) *** | 19 (18.8) | |
| C | 14 (8.1) | 4 (6.7) | 0 (0) | 10 (9.9) | |
| D | 4 (2.3) | 2 (3.3) | 0 (0) | 2 (1.9) | |
| E | 9 (5.2) | 2 (3.3) | 0 (0) | 7 (6.9) | |
| F | 5 (2.9) | 0 (0) | 0 (0) | 5 (4.9) | |
| NC | 13 (7.5) | 0 (0) (*) | 1 (3.2) | 12 (11.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, J.A.S.; Santos, F.F.; Santos-Neto, J.F.; Trovão, L.O.; Valiatti, T.B.; Pinaffi, I.C.; Vieira, M.A.M.; Silva, R.M.; Falsetti, I.N.; Santos, A.C.M.; et al. Molecular Epidemiology and Presence of Hybrid Pathogenic Escherichia coli among Isolates from Community-Acquired Urinary Tract Infection. Microorganisms 2022, 10, 302. https://doi.org/10.3390/microorganisms10020302
Nascimento JAS, Santos FF, Santos-Neto JF, Trovão LO, Valiatti TB, Pinaffi IC, Vieira MAM, Silva RM, Falsetti IN, Santos ACM, et al. Molecular Epidemiology and Presence of Hybrid Pathogenic Escherichia coli among Isolates from Community-Acquired Urinary Tract Infection. Microorganisms. 2022; 10(2):302. https://doi.org/10.3390/microorganisms10020302
Chicago/Turabian StyleNascimento, Júllia A. S., Fernanda F. Santos, José F. Santos-Neto, Liana O. Trovão, Tiago B. Valiatti, Isabel C. Pinaffi, Mônica A. M. Vieira, Rosa M. Silva, Ivan N. Falsetti, Ana C. M. Santos, and et al. 2022. "Molecular Epidemiology and Presence of Hybrid Pathogenic Escherichia coli among Isolates from Community-Acquired Urinary Tract Infection" Microorganisms 10, no. 2: 302. https://doi.org/10.3390/microorganisms10020302