
Metabolites and Their Bioactivities from the Genus Cordyceps


State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, School of Life Sciences, Yunnan University, Kunming 650091, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Microorganisms 2022, 10(8), 1489; https://doi.org/10.3390/microorganisms10081489
Submission received: 5 July 2022
/
Revised: 21 July 2022
/
Accepted: 21 July 2022
/
Published: 24 July 2022
(This article belongs to the Special Issue Secondary Metabolism of Microorganisms)
Abstract
:The Cordyceps genus is a group of ascomycete parasitic fungi, and all known species of this genus are endoparasites; they mainly feed on insects or arthropods and a few feed on other fungi. Fungi of this genus have evolved highly specific and complex mechanisms to escape their host’s immune system and coordinate their life cycle coefficients with those of their hosts for survival and reproduction; this mechanism has led to the production of distinctive metabolites in response to the host’s defenses. Herein, we review approximately 131 metabolites discovered in the genus Cordyceps (including mycelium, fruiting bodies and fungal complexes) in the past 15 years, which can be used as an important source for new drug research and development. We summarize chemical structures, bioactivity and the potential application of these natural metabolites. We have excluded some reports that originally belonged to Cordyceps, but whose taxonomic attribution is no longer the Cordyceps genus. This can and will serve as a resource for drug discovery.
1. Introduction
Cordyceps sinensis is a renowned Chinese herbal medicine and has been widely used for medicinal treatment in China for over 300 years [1]. C. sinensis grows in very limited habitats, its yields decrease year by year, and its use is finite because of its high price and limited availability [2]. Researchers have been seeking substitute materials by investigating the fermentation and culture of fungi separated from C. sinensis and other Cordyceps species [3]. Cordyceps is the most numerous and diverse genus of the Clavellaceae family, of which 629 species have been identified, according to MycoBank (https://www.mycobank.org; accessed on 8 June 2022). It is a class of ascomycete parasitic fungi. All known species act as endoparasites, feeding mostly on insects and other arthropods and a little on other fungi. This survival mechanism of Cordyceps leads to the production of distinctive metabolites in response to host defenses, which is an important source for new drug research and development [4,5]. There are many species of Cordyceps. They are abundant in humid climates and tropical forests, widely distributed in North America; Europe; East and Southeast Asian countries, especially China, Japan, Nepal, Vietnam, Bhutan, Korea and Thailand, although some other species are also found in different habitats in other regions, indicating a global distribution. Species in the genus Cordyceps are widely accepted for using as food and medicine and good reviews have been published. For example, a review published by Zhou et al. reported on natural products, pharmacological functions, and novel products in Cordyceps sinensis [6]. Similarly, Olatunji et al. reviewed the advanced developments in traditional uses, phytochemistry and pharmacology of Cordyceps fungi, with a primary focus on C. sinensis and C. militaris [7]. On the other hand, Chen et al. provided an overview on the safety concerns of fungal fruiting bodies of several Cordyceps species in terms of their existence as food supplements or as animal feed by-products and analyzed the conservatism of gene clusters between Cordyceps and other fungi involved in toxin production [8]. In the past 15 years, researchers have reported many new strains and active metabolites, and the development of biotechnology has provided more possibilities for investigating these novel and diverse metabolites. Moreover, many polysaccharides have been isolated and identified in Cordyceps and have showed various biological activities. Due to the taxonomic characteristics of fungi, some metabolites of fungi that previously belonged to the genus Cordyceps but are no longer characterized as Cordyceps are often cited. We will summarize the bioactive compounds from the Cordyceps genus that have been reported in the literature since 2007. The species of the genus Cordyceps present an important source of therapeutic effect for various diseases due to the presence of bioactive components, which can serve as potential leads for drug discovery.
2. Nucleosides and Their Activities
Nucleoside compounds are the main metabolites and active components of Cordyceps fungi. Cordysinin B (1) was separated from the mycelium of C. sinensis and inhibited the formation of superoxide anions and the release of elastase with an IC50 value of 0.15 μM [9]. Two new nucleosides, N6-4-methylbutyrat-adenosine (2) and 3′-deoxy-6-O-methylinosine (3), were isolated from C. militaris and did not show anti-inflammatory inhibitory activity under 100 μM [10]. 3’-Deoxyinosine (4) was isolated from C. militaris and showed a weak toxic effect on cancer cells A549, PANC-1 and McF-7 and exhibited powerful cytotoxicity against MCF-7 cells at a concentration of 30 μM [11]; moreover, it was previously reported to have antigenic activity. In addition, when compound 4 was combined with doxorubicin, it inhibited the metastasis and proliferation of breast cancer cells. According to the Chou-Talalay method, when 80 μmol/L 3’-deoxyinosine (4) and 1 μmol/L doxorubicin were used together, the synergistic effect was the strongest, and the CI value and the cell inhibition rates were 0.665 and 60.31 ± 1.06%, respectively [12]. Two metabolites, (2-amino-N-((2S,3S,4R,5R)-5-(6-amino-9H-purin-9-yl)-4-hydroxy-2-(hydroxymethyl)–tetrahydrofuran-3-yl)-6-ureido-hexanamide) (5) and (2-amino-N-((2S,3S,4R,5R)-5-(6-amino-9H-purin-9-yl)-4-hydroxy-2-(hydroxymethyl)tetrahydrofuran-3-yl)-6-guanidinohexanamide) (6), were identified from C. militaris. Anti-tumor experiments showed that compounds 5 and 6 considerably inhibited the propagation of HepG2 liver cancer cells at 72 h and their IC50 values were 0.09 μM and 0.51 μM, respectively [13]. A new ribonucleotide of 5’-(3’’-deoxy-β-D-ribofuranosyl)-3’-deoxyadenosine (7) as well as a known 3’-deoxyadenosine (8) were isolated from the C. militaris; 3’-deoxyadenosine 8 was shown to inhibit the expression of the NF-κB reporter gene in HeLa cells induced by TNF at concentrations of 3–100 μM, but compound 7 showed no inhibitory activity [14]. A new nucleoside cordyrrole B (9), an acetylated form of N6-(2-hydroxyethyl)adenosine, was obtained from the active components of C. militaris and showed the significant inhibition of pancreatic lipase activity at 100 μM [15]. 6-Acetylpurine (10) was isolated from C. militaris [16]. The structures of 1–10 are shown in Figure 1.
Cordycepin is a vital active component of Cordyceps. During in-depth research on cordycepin, it was found that cordycepin has a wider range of biological activities, and many new mechanisms of action were discovered. Due to the emergence of mutant strains, the coronavirus disease 2019 (COVID-19) is ongoing globally, often causing severe acute respiratory syndrome and leading to the death of some patients. Using computational methods, researchers predicted a possible inhibitory affinity of cordycepin to the main SARS-CoV-2 protein target [17]. The latest research shows that cordycepin can effectively inhibit the reproduction of new SARS-CoV-2 drug-resistant strains, and its EC50 was about 2 μM in in vitro anti-SARS-CoV-2 assays, which is superior to remdesivir and its active metabolite GS-441524 [18]. In addition, it was found that cordycepin can inhibit Dengue virus replication and significantly reduced DENV protein at EC50 of 26.94 μM [19]. Other research on the activity of cordycepin found it could protect PC12 cells from the neurotoxicity induced by 6-hydroxydopamine through its powerful antioxidant activity [20]. Additionally, cordycepin modulated adenosine A1 receptors to increase long-term enhancement-capability formation and neuronal survival in the BCCAO model and glutamate-enticed HT22 neuronal cell death via the p38/JNK/ERK pathway [21,22]. In recent years, progress has been made in the study of the anti-cancer mechanisms of cordycepin, and many reviews show that cordycepin may facilitate tumor cell death via cysteine–aspartic proteases (caspases), mitogen-activated protein kinase (MAPK) and glycogen synthase kinase (GSK)-3β pathways mediated by putative adenosine receptors, death receptors and/or epidermal growth factor receptors (EGFR) [23,24]. In particular, cordycepin regulates the phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) signaling pathway and inhibits cyclin-dependent kinase 2 (Cdk-2), extracellular signal-regulated kinase 1/2 (ERK1/2) and Rb/E2F1 and fibroblast growth factor receptors 1–4 (FGFR 1–4) regulate the cell cycle and further reduce the growth of testicular tumors, gastric cancer cells and cervical cancer cells [25,26]. In addition, cordycepin also regulates diverse signaling proteins, such as hedgehog, glioblastoma protein (GLI), DNA-dependent protein kinase (DNA-PK) and ERK, to induce cancer cell apoptosis [27,28].
3. Non-Ribosomal Peptides and Alkaloids
Three new cordysinins C-E (11–13) were isolated from the mycelia of C. sinensis, but showed no antioxidant activity even at 500 μM [9]. Two new pyrrole alkaloid derivatives, 2-carboxaldehyde-1-(4-aminobutyl-5-(methoxymethyl)-1H-Pyrrole (14), and 2-carboxaldehyde-5-(methoxymethyl)-1-(2-oxo-3-piperidinyl)-1H-pyrrole (15), were separated from C. militaris, and the anti-inflammatory effects results showed no significant inhibitory activity higher than 100 μM [10]. A new alkaloid cordyrrole A (16) was separated from the extract of C. militaris and demonstrated the inhibition of adipocyte differentiation and pancreatic lipase activity at a concentration of 100 μM [15]. Two new alkaloid metabolites, cordytakaoamides A (17) and B (18), were isolated from cultures of C. takaomontana NBRC 101754, and the absolute configuration of cordytakaoamides A and B was expounded by the total synthesis of cordytakaoamide A and the experimental circular dichroism [29]. Five new alkaloids, cordycepamides A-E (19–23) were isolated from C. sinensis. Cordycepamide D (22) showed moderate radical scavenging activity, its IC50 value concentration was 51.42 ± 3.08 μM in an analysis of DPPH [30]. Three new macrocyclotetralactamides, gunnilactams A-C (24–26), were separated from the deep fermentation liquid of C. gunnii; 25 and 26 were isomers. Gunnilactam A (24) was selectively cytotoxic against human prostate cancer C42B cells and its IC50 was 5.4 μM [31].
A cordycepic pigment cordycepene (27) was identified from C. militaris, and it had remarkable DPPH radical-scavenging activity; the IC50 was 0.81 mg/mL in antioxidant assays. In anti-aging assays, the activities of CAT, GSH-Px and SOD increased by 201.05%, 708.26% and 341.50%, respectively, and the degree of MDA decreased by 29.92%, when the concentration of compound 27 enhanced from 0 to 50 μg/mL [32]. Two new compounds, cordyceamides A (28) and B (29), were separated from C. sinensis (BERK.) SACC. Compounds 28 and 29 had cytotoxic effects on the cells of lines A375, L929 and Hela [33]. A new cerebroside, cordycerebroside B (30), was isolated from C. militaris and showed remarkable inhibition activity against PTP1B with an IC50 value of 4.68 ± 0.18 μM [34]. A new lactam-fused 4-pyrone cordylactam (31) was obtained from the Cordyceps sp. BCC 12671; however, its biological activity was not tested [35]. The structures of 11–31 are shown in Figure 2.
Cardinalisamides A–C (32–34) were separated from C. cardinalis NBRC 103832. Cardinalisamides A–C showed antitrypanosomal activity against Trypanosoma brucei and their IC50 were 8.56, 8.65 and 8.63 mg/mL, respectively, in vitro, and had IC50 values of 18.48, 14.00 and 23.84 mg/mL, respectively, against normal human diploid fibroblasts (MRC-5 cells) in cytotoxicity assays [36]. Cordycecin A (35) and two known compounds beauvericins E (36) and J (37) were separated from the ascocarps and insect-body tranches of C. cicadae. Beauvericin J (37) was cytotoxic against HepG2 and HepG2/ADM cells, whose IC50 values were in the range of 5.04 ± 0.20 μM and 2.67 ± 0.09 μM, respectively; beauvericin E (36) revealed moderate inhibitory effects and its IC50 value was in the range of 13.67 ± 2.59 μM and 14.48 ± 1.68 μM, respectively [37]. Cordycommunin (38) was separated from the Ophiocordyceps communis BCC 16475 and it had a growth inhibiting effect on Mycobacterium tuberculosis H37Ra and its MIC value was 15 µM. Compound 38 also exhibited slight cytotoxicity to KB cells and its IC50 value was 45 µM [38]. Beauverolide Jb (39) was obtained from C. javanica [39]. A tripeptide, arginylphenylalanyl-methionine (40), isolated from C. gunnii, with the molecular formula of C20H32N6O4S, is a sedative and hypnotic active substance [40]. Other polypeptide compounds have had less report published in recent years. However, recent studies have shown that cordyceps polypeptide complexes are helpful for improving learning and memory, but their mechanism of action needs further study [41]. The structures of 32–40 are shown in Figure 3.
4. Polyketides
Three new compounds, paecilomycones A-C (41–43), were identified from a methanol extract of C. gunnii. Paecilomycones A-C showed significant tyrosinase inhibitory activity, and the IC50 values were 0.11 μM, 0.17 μM and 0.14 μM, respectively. Structure and activity research showed that the tyrosinase inhibition activity was connected to the amount of hydroxyl groups on the paecilomycones.
The structure of compound 41 is very similar to the anti-HIV target flurane (anti-HIV, the IC50 value was 1.7 μM), while the compound 43 has a NH2 group in C-9 rather than the usual -OH group, which indicates that compounds 41 and 43 may be promising anti-HIV products with high importance for follow-up studies [42]. Two new compounds, 2-(3-carboxy-2-hydroxypropyl)-3-methyl-2-cyclopentenone (44) and 5-(2- hydroxyethyl)-2-furanacetic acid (45), were isolated from the C. cicadae mycelia [43]. Two compounds, rugulosin (46) and skyrin (47), were identified from C. formosana; the LD50 of compounds 46 and 47 were 18.3 ± 0.2 and 103.7 ± 5.9 μg/mL against CHO cells in cytotoxic assays [44]. Opaliferin (48), which has a novel C19 skeleton, was separated from the Cordyceps sp. NBRC 106954. Opaliferin showed slight cytotoxicity against HeLa, HSC-2 and RERF-LC-KJ tumor cell lines at 100 μM, and the inhibitory rates were 60%, 30% and 20%, respectively [45].
Four novel isoflavone methyl-glycosides, daidzein 7-O-β-D-glucoside 4″-O-methylate (49), glycitein 7-O-β-D-glucoside 4″-O-methylate (50), genistein 7-O-β-D-glucoside 4″-O-methylate (51) and genistein 4′-O-β-D-glucoside 4″-O-methylate (52) were obtained from C. militaris which was grown on germinated soybeans. Compound 52 showed marked antiallergic activity [46]. Three new compounds fumosoroseanosides A (53) and B (54) and fumosoroseain A (55) were isolated from C. fumosorosea. At experimental doses of 10–30 μg, compounds 53 and 54 exhibited excellent (p < 0.05) antibacterial and antifungal activity compared to the negative control group. In a range of concentrations from 100 μM to 400 μM, compound 55 could lengthen the life of nematodes; at 200 μM, it showed optimal activity (p < 0.05), and the average life of the nematodes was lengthened by 11.3% [47]. The structures of 41–56 are shown in Figure 4.
Two new compounds 2-(5-(3-oxobutyl) furan-2-yl) acetic acid (56) and cordycepone (57) were isolated from Cordyceps spp., a strain formed by cell fusion in Cordyceps militaris and Cordyceps cicadae. Compound 56 showed a slightly inhibiting effect on AChE, and its inhibition rate was 16.41% [48]. A new glycosylated asperfuran (58) was isolated from C. javanica, whose biological activity has not been reported [39]. A new stereoisomer of clonostachydiol, cordybislactone (59), and its hydrolyzed derivative (60), were obtained from Cordyceps sp. BCC 49294 [49], whose biological activity has not been reported. Five new anthraquinones, morakotins A–E (61–65), were purified from the C. morakotii BCC 56811 and demonstrated antimicrobial activity. Morakotin C (63) showed moderate antifungal activity against Candida albicans (the value of IC50 was 25.87 µg/mL) and antibacterial effect on Bacillus cereus (the value of MIC was 12.5 µg/mL). Morakotin D (64) showed marked antibacterial activity against B. cereus (the value of MIC was 3.13 µg/mL) and Staphylococcus aureus (the value of MIC was 6.25 µg/mL) [50]. A novel derivative MA-1 [(2R,3R,4R,5R)-1,6-bis(4-(2,4,4-trimethylpentan-2-yl)phenoxy)hexane-2,3,4,5-tetraol] (66), was synthesized from the constituents of C. militaris. MA-1 (66) demonstrated powerful anti-cancer potential for the cure of H358, A549 and H460 cell lines, and its value of IC50 was 5 μM for A549 human lung cancer cells [51]. 6,7,2′,4′,5′-Pentamethoxyflavone (67) was isolated from C. militaris and its inhibition of hemolysis was 92.08 ± 1.85% at 250 μg/mL [52]. Cordypyrones A (68) and B (69) were isolated by the heterologous expression of a biosynthetic gene cluster from C. militaris in Aspergillus nidulans. Cordypyrones A (68) and B (69) showed moderate inhibitory effects on ATCC 9372 and both of their MIC values were 16 µg/mL; the MIC value were 16 and 8 µg/mL, respectively, against ATCC 49064; and the MIC values were 32 µg/mL against ATCC 25922 [53]. The structures of 57–69 are shown in Figure 5.
5. Sterols and Terpenoids
It has been reported that sterols in C. sinensis have anti-tumor activity, immunosuppression, anti-arteriosclerosis and antibacterial activities. However, the active sterols in C. militaris are seldom reported.
Reports from 2015 showed a new anti-cancer compound named (3R,9R,10S,13S,14S,17S)-17-((2S,5R,E)-5,6-dimethylhept-3-en-2-yl)-10,13-dimethyl-2,3,4,9,10,11,12,13,14,15,16,17-dodecahydro-1H-cyclopenta[α]phenanthren-3-ol (70), was purified from C. militaris. Within 48 h, compound 70 exhibited dosage-dependent and time-dependent inhibition on the growth of A549 cells, and lung cancer cells were almost no longer growing at a concentration of 120 μg/mL. These results revealed that compound 70 was likely to inhibit the proliferation of human lung cancer A549 cells [54]. Cholest-5-en-3β-ol (71), 22-tetraen-3-one (72), 5α-cholest-3,6-dione (73) and cholest-4-en-3-one (74) were separated from C. militaris for the first time [14]. Four novel carotenoids were separated from C. militaris and confirmed as xanthophylls and their names were cordyxanthin-I (2,3,2′,3′-tetradehydro-18,16′,17′,18′-tetranor-ε,ε-carotene-5,5′,1′-triol) (75), cordyxanthin-II (2,3,2′,3′-tetradehydro-18,1′,16′,17′,18′-pentanor-ε,ε-carotene-5,5′,-diol) (76), cordyxanthin-III (2,3,2′,3′-tetrade hydro-18,17′,18′-trinor-ε,ε-carotene-5,5′,-diol) (77) and cordyxanthin-IV (2,3′,3′-tetradehydro-18,18′-dinor-ε,ε-carotene-5,5,-diol) (78) [55]. The structures of 70–78 are shown in Figure 6.
6. Aromatics and Their Derivatives
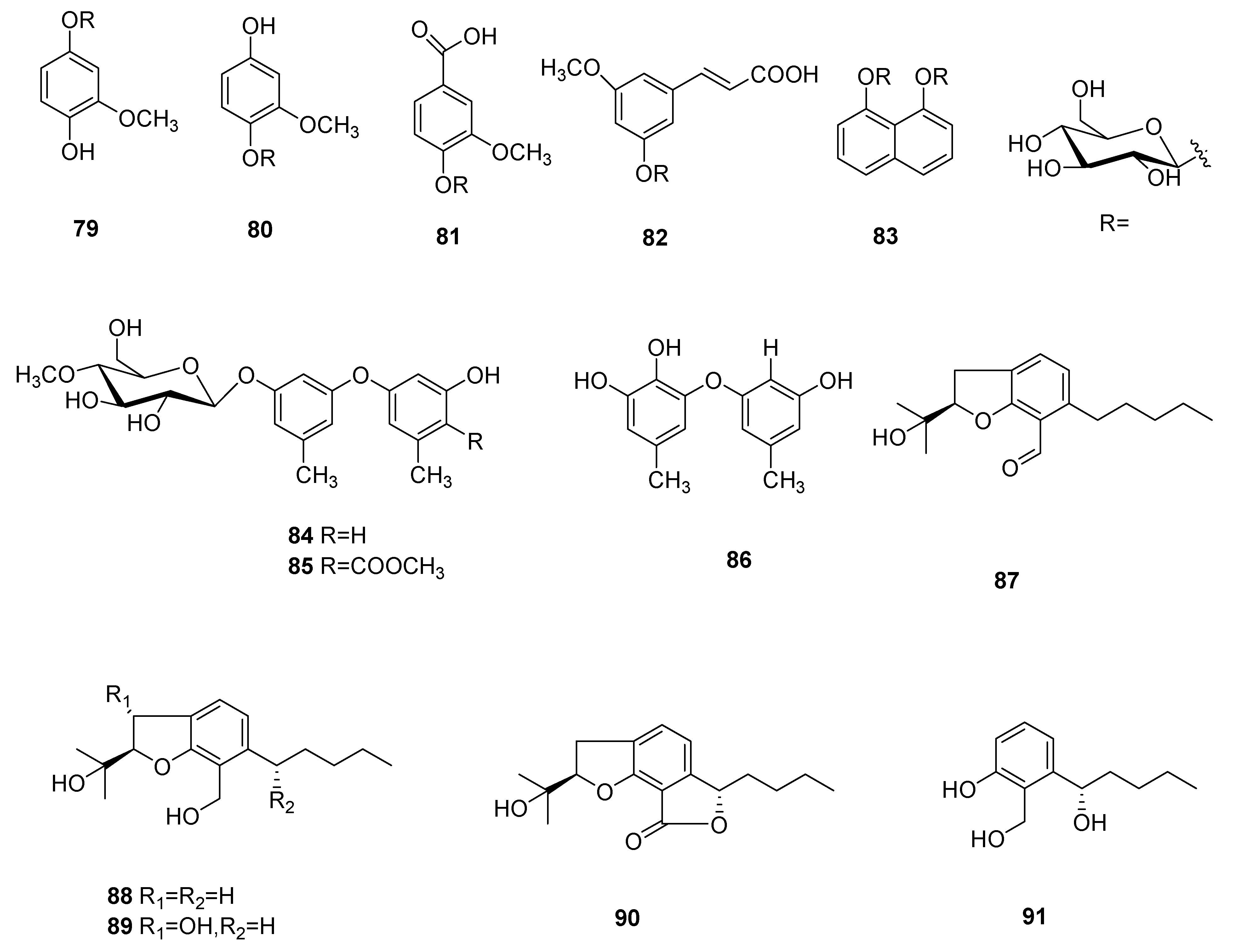
Five new aromatics bearing a 4-O-methylglucose unit, named 3-methoxy- 1,4-hydroquinone 1-(4′-O-methyl-β-glucopyranoside) (= 4-hydroxy-3-methoxy phenyl 4-O-methyl-β-glucopyranoside) (79), 3-methoxy-1,4-hydroquinone 4-(4′-O-methyl-β-glucopyranoside) (=4-hydroxy-2-methoxyphenyl 4-O-methyl-β-glucopyranoside) (80), vanillic acid 4-(4′-O-methyl-β-glucopyranoside) (=3-methoxy-4-[(O-methyl-β-glucopyranosyl) oxy]benzoic acid) (81), 5-methoxycinnamic acid 3-O-(4′-O-methyl-β-glucopyranoside) (=(2E)-3-[3-methoxy-5-[(4-O-methyl-β-glucopyranosyl)oxy]phenyl]prop-2-enoic acid (82) and naphthalene-1,8-diol 1,8-bis(4′-O-methyl-β-glucopyranoside) (=naphthalene-1,8-diyl bis(4-O-methyl-β-gluco- pyranoside) (83) were separated from the mycelia of C. cicadae [56].
Cordyols A–C (84–86) were separated from the Cordyceps sp. BCC 1861. Cordyol C (86) showed anti-HSV-1 activity, its IC50 value was 1.3 μg/mL and it had a cytotoxic effect on BC and NCI-H187 cancer cell lines, whose IC50 values were 8.65 μg/mL and 3.72 μg/mL, respectively. Cordyol A (84) showed slight antifungal activity and its MIC value was 100 μg/mL [57]. Annullatins A–E (87–91) were identified from C. annullata. Suberoyl bis-hydroxamic acid (SBHA), a histone deacetylase (HDAC) inhibiting agent, was added to the culture medium. Compounds 87 and 88 reduced cAMP levels by 47.9% and 40.8%, respectively, in a CB1-expressing assay. Compounds 87 and 88 decreased cAMP levels by 69.9% and 15.1%, respectively, and compound 89 increased the cAMP standards by 47.5% (CB1 and CB2 were cannabinoid receptors) in CB2-expressing assays [58]. The structures of 79–91 are shown in Figure 7.
7. Protein
There are also proteins in C. sinensis. As far as we know, these proteins have a variety of activities, such as antifungal, anticancer and antiviral activities, etc. These research results show that C. sinensis proteins also play an important role in biology. In recent years, studies on C. militaris proteins have also been gradually increasing. A new anti-tumor protein was separated from the seed entity of C. militaris, named CMIP (92). CMIP showed anti-metastasis activity by reducing the amount of tumor nodules in the lung of tumor-bearing mice and extended their lives on a mouse model of 4T1 breast cancer lung metastasis. The results showed that CMIP possessed immune regulatory activity [59].
In addition, a new protein with a molecular mass of 18.0 kDa was separated from C. militaris and named CMP (93); however, it is a nocuous protein that can cause cell apoptosis via a mitochondrion-dependent mechanism [60]. A glycopeptide (Cs-GP1) (94) was purified from the strain C. sinensis Cs-HK1, and its molecular weight was 6.0 kDa. Furthermore, Cs-GP1, mostly consisting of glucose and mannose at 3.2:1.0 molar ratio, showed notable antioxidant activities (1183.8 μmol Trolox/g and 611.1 μmol Fe(II)/g) [61]. A protein designated the Cordyceps militaris protein (CMP I) (95) was the first protein from C. militaris; it is a 12 kDa protein in the form of dimers. CMP I exhibited powerful antifungal activities against the growth of the Fusarium oxysporum. In the CCK assays, CMP I (95) significantly inhibited the viability of MCF-7 cells, its IC50 value was 9.3 μM and the IC50 against 5637 cells was 8.1 mΜ. However, CMP I showed almost no inhibitory effect on A-549 cells [62]. Two peptides, named VPRKL(Se)M (Se-P1) (96) and RYNA(Se)MNDYT (Se-P2) (97), were purified from Se-enriched C. militaris and demonstrated neuroprotective effects, and compared to the damage group, Se-P1 and Se-P2 increased PC-12 cell viability by 30 and 33%, respectively. In addition, Se-Ps may moderate cognitive damage in LPS-injured mice (p < 0.05). Therefore, Se-Ps has the potential to be an alternative to drugs to prevent and/or treat AD (Alzheimer’s disease) [63].
A fibrinolytic enzyme (98) was identified from C. militaris, and its enzyme activity was 1682 U/mg; its molecular weight and pI were 32 kDa and 9.3 ± 0.2, respectively; its first-rank pH and temperature were 7.4 and 37 °C, respectively. This fibrinolytic enzyme could hydrolyze fibrin(ogen) rapidly and cleave the α-chains more effectively than β- and γ-chains, and it also could degrade thrombin. Therefore, it could be a potential natural agent for oral fibrinolytic medical treatment or to prevent the formation of blood clots [59]. An antifungal peptide, cordymin (99) with a unique N-terminal amino acid sequence was purified from the C. militaris; its molecular mass was 10,906 Da. Cordymin inhibited the mycelial growth of Bipolaris maydis, Rhizoctonia solani, Mycosphaerella arachidicola and Candida albicans and their IC50 values were 50, 80, 10 and 0.75 μM, respectively. Cordymin also had an IC50 of 55 μM for inhibiting HIV-1 reverse transcriptase [64]. A novel protease (100) was purified and characterised from the edible fungus C. sinensis; its molecular weight was approximately 43 kDa; and its first-rate pH and temperature were 9.5 and 30 °C, respectively [65]. A novel fibrinolytic enzyme named CMase (101) was purified from C. militaris for the first time; its molecular mass was approximated to be 27.3 kDa and its optimal pH and temperature were pH 6.0 and 25 °C, respectively [66]. These reports suggest that C. militaris represents a new source of proteins.
8. Polysaccharide
Cordyceps polysaccharide is the important bioactive component in C. sinensis, which has demonstrated anti-cancer, anti-oxidant, anti-viral, immunomodulation properties and the improvement of liver function [67]. Furthermore, studies have shown that the sulfated exosaccharide of C. sinensis can enhance its antioxidant activity [68].
A polysaccharide (SDQCP-1) (102) was separated from C. militaris that was cultivated on hull-less barley; it mainly consisted of mannose, glucose and galactose at 13.3:1.0:9.7 molar ratio; its average molecular weight was 19.3 kDa. The antioxidant and immunomodulatory activities showed that SDQCP-1 had great antioxidant capacity, its ORACFL value was 24.7 mmol Trolox/g and TEAC value was 202.4 μmol Trolox/g. SDQCP-1 also motivated macrophages to liberate NO, IL-6, TNF-α and IL-10 and mostly facilitated the M1 polarization of macrophages. The findings suggest that SDQCP-1 has potential as a natural antioxidant and immunomodulator in functional foods or drugs [69].
A novel polysaccharide CMP-1 (103), with an average molecular weight of 4.3 kDa, was isolated from the fruit body of cultured C. militaris with antioxidant, immune stimulatory and anti-tumor activity. CMP-1 showed free radical-scavenging effects, ferrous-ion chelating ability and reducing power in antioxidant assays. Furthermore, CMP-1 considerably encouraged mouse splenocyte proliferation in vitro. It also inhibited the proliferation of HepG2, HeLa, HT-29 and K562 cells and the IC50 values were 176.29, 162.59, 137.66 and 364.01 μg/mL, respectively, in cytotoxicity assays [70]. A novel polysaccharide CM3-SII (104) was isolated from C. militaris with potential hypolipidemic effect; it consisted of mannose, glucose and galactose at a 10.6:1.0:3.7 molar ratio. The interference of CM3-SII considerably increased the protein expression of LDLR and intracellular levels of PCSK9 at the concentration of 100 and 200 μg/mL [71]. A homogeneous exopolysaccharide (EPS-III) (105) was obtained from C. militaris with hypoglycemic activity; its average molecular weight was 1.56 × 103 kDa. In a hypoglycemic experiment of EPS-III in vivo, the inhibition rate of α-glucosidase was considerably enhanced when the concentration of EPS-III was increased, and at a concentration of 3 mg/mL the inhibition rate reached 55.94 ± 1.34%. In addition, studies showed that EPS-III moderated weight loss, decreased plasma glucose concentration, promoted glucose tolerance, secured immune organs and repaired dyslipidemia to moderate diabetes in STZ-induced diabetic mice [72]. An alkaline-extracted polysaccharide (CM3II) (106) was purified from C. militaris with anti-atherosclerotic effects; it was mainly composed of mannose, glucose and galactose at a 1.4:1.0:1.2 molar ratio. In experimental mice with atherosclerosis induced by a high-fat diet, Oil Red O staining results showed that simvastatin and CM3II interference decreased the atherosclerotic lesion/lumen ratio by 6.1% and 17.8% (p < 0.05), respectively. Moreover, CM3II increasingly decreased the TC and TG standards [73]. Two new polysaccharides, SCP II-1 (107) and SCP II-2 (108), were purified from silkworm Cordyceps and demonstrated antioxidant and antitumor activity; the molecular weight of SCP II-1 was 35.2 kDa and SCP II-2 was 23.4 kDa; they consisted of ribose, mannose, glucose and galactose in a molar ratio of 1.0:27.38:8.52:17.99 and 1.0:21.21:1.95:14.28, respectively. In the DPPH radical scavenging activity assays, the DPPH radical removal degrees of SCP II-1 and SCP II-2 were 88.328% and 75.028%, respectively. The DPPH radical removal IC50 values were less than 0.5 mg/mL. In the antitumor activity experiment, SCP II-1 had an IC50 value of 119.34 ± 1.76 μg/mL against HepG2 cell proliferation [74]. A polysaccharide (CMP-III) (109) was isolated from C. militaris; its average molecular weight was 4.796 × 104 kDa; it was composed of glucose, mannose and galactose at an 8.09:1.00:0.25 molar ratio. Moreover, the studies of immunomodulatory functions showed that CMP-III could enhance macrophage phagocytosis and the release of NO, TNF-α and IL-6 at a concentration of 25–200 μg/mL [75]. An acidic exopolysaccharide (AESP-II) (110) was purified from C. militaris that demonstrated immunological activity. AESP-II consisted of mannose, glucuronic acid, rhamnose, galactose acid, N-acetyl-galactosamine, glucose, galactose and arabinose at a 1.07:5.38:1:3.14:2.23:15:6.09:4.04 molar ratio, and its molecular weight was 61.52 kDa. In addition, AESP-II considerably increased the proliferation of B lymphocytes in a dose-dependent manner and significantly increased the proliferation of T lymphocytes at a low dose (25 mg/kg body weight) in an animal experiment [76]. A poly-N-acetylhexosamine (polyhexNAc) (111) with an average molecular weight of about 6 kDa was isolated from C. sinensis Cs-HK1; its molecular structure is a [-4-β-D-ManNAc-(1 → 3)-β-D-GalNAc-(1 →] disaccharide repeating unit in the chief chain. It exhibited remarkable antioxidant activities (330 μmol Trolox/g and 45.7 μmol Fe(II)/g) and showed meaningful cytoprotective activity at a concentration of 10–200 mg/mL [77]. A novel polysaccharide (CMPA90-1) (112) with antioxidant and anti-tumor activity was isolated from C. militaris. CMPA90-1 consisted of arabinose, mannose and galactose at a 1.00:2.89:2.03 molar ratio; it exhibited inhibitory activity against A549 cells and its IC50 value was 39.08 μg/mL in the cytotoxicity assay [78].
Two novel polysaccharides, PSCK2-2 (113) and PSCK2-3 (114), which demonstrated good antioxidant properties and powerful protective effects against DNA damage, were isolated from C. kyushuensis. PSCK2-2 was composed of Fru-Man-Rha GalN and Ara at a 1.0:1.19:0.11:0.11:0.34 molar ratio, and PSCK2-3 was composed of Fru-Man-Rha Glu and Ara at a 1:1.29:0.14:0.07:0.32 molar ratio. The hydroxyl radical scavenging activities showed that the inhibition rate was 98.33 ± 3.29% (p < 0.05) and 55.83 ± 2.41% (p < 0.05), respectively, when the concentration of PSCK2-2 and PSCK2-3 was 6 mg/mL; their IC50 values were 1.5 mg/mL and 4.8 mg/mL, respectively [79]. Cordyceps sinensis polysaccharide 1 (115), with a molecular weight of 1.17 × 105 Da, was purified from C. sinensis. It consisted of (1 → 6)-linked α-D-Glc and α-D-Gal, with minor β-(1 → 4)-D-Xyl and β-(1 → 4)-D-Man residues presumably seated in the side chains with a trace quantity of α-(1 → 3)-L-Rha residue. In the assays of the restrained proliferation of sarcoma 180 cells, polysaccharide 1 showed significant activity and induced apoptosis in a dosage-dependent manner [80]. A novel polysaccharide CME-1 (116) was identified from C. sinensis, it was composed of mannose and galactose at a 4:6 molar ratio and its molecular weight was 27.6 kDa. For inhibiting human platelet aggregation, CME-1 with a concentration in the range of 2.3–7.6 μM was highly effective when fueled by collagen, thrombin and arachidonic acid except for U46619 [81].
Two polysaccharides, WIPS (117) and AIPS (118), were separated from C. sinensis Cs-HK1. The molecular weight of WIPS and AIPS were 1.18 × 103 kDa and 1.15 × 103 kDa, and they were elucidated as α-D-glucans with a backbone of (1 → 4)-linked α-D-Glcp (>60%). AIPS inhibited tumor growth by about 28%, and WIPS inhibited about 12% of melanoma tumor growth in mice. In addition, AIPS and WIPS augmented the impact on the T-cell proliferation and viability in the lymphocyte proliferative assay [82]. A novel acidic polysaccharide AEPS-1 (119), with immunomodulatory properties, fractionated from the exopolysaccharide produced by C. sinensis Cs-HK1, has an α-D-(1 → 3)-Glcp backbone structure. It consists of glucopyranose (Glcp) and pyrano-glucuronic acid (GlcUp) at an 8:1 molar ratio plus a trace amount of mannose; its average molecular weight was about 36 kDa. When the concentration of AEPS-1 was increased from 25 to 250 μg/mL, the cytokine IL-10 also increased but the other three cytokines, TNF-a, IL-1b and IL-6 in the culture medium of macrophage Raw264.7 cells, were decreased [83].
A polysaccharide MCMP (120) was purified from C. militaris and demonstrated anti-tumor activity. The molecular weight of MCMP was 8.1 kDa, and it consisted of mannose, rhamnose, galactose and glucose at a 59.36:1:8.31:39.50 molar ratio. At a concentration of 8 mg/mL, the inhibition rates of MCMP on HepG-2 cells, Hela cells and mesangial cells were 57.11%, 67.11% and 58.74% after 72 h incubation, respectively [84]. A novel polysaccharide, with a molecular weight of about 82 kDa, named cordysinocan (121), was obtained from Cordyceps. In cultured T-lymphocytes, cordysinocan induced cell proliferation and its EC50 was 6 μg/mL; it also induced IL-2 secretion with an EC50 of 8.5 μg/mL. Furthermore, in cultured macrophages, cordysinocan induced macrophage phagocytosis and its EC50 was 5 μg/mL; it also increased phagocytosis and the enzymatic efficacy of acid phosphatase [85]. A polysaccharide, namely CBP-1 (122), was separated from C. militaris. CBP-1 had a backbone of (1 → 4)-α-D-mannose residues, which sometimes branched at O-3. The branches consisted of (1 → 4)-α-D-glucose residues and (1 → 6)-β-D-galactose residues and ended with β-D-galactose residues. CBP-1 demonstrated hydroxyl radical-scavenging activity and its IC50 value was 0.638 mg/mL in in vitro antioxidant assays [86].
A novel polysaccharide PS-T80 (123) was collected from Ophiocordyceps sobolifera and demonstrated antioxidant activities; it was mainly composed of β-D-glucose and α-D-mannose at a 2:1 molar ratio and its average molecular weight was 74 kDa. The structure studies revealed that PS-T80 was a mannoglucan, owing to the repeating unit of [→ 3)-β-D-Glcp-(1 → 3)-α-D-Manp-(1 → 3)-β-D-Glcp-(1 →]. DPPH radical scavenging activity was improved in a dose-dependent manner from a PS-T80 (123) concentration in antioxidant assays, and the IC50 value was approximately 0.97 mg/mL [87].
A novel fungal polysaccharide (PS-T100) (124) was purified from Ophiocordyceps sobolifera; however, its biological activity was not tested. PS-T100 was mainly composed of [→ 3)-β-D-Glcp-(1 → 3)-α-D-Manp-(1 → 3)-β-D-Glcp-(1 → 3)-α-D-Manp-(1 → 3)-β-D-Glcp-(1 →] repeating units; its average molecular weight was 2.29 × 102 kDa [88]. Two novel polysaccharides, CM1 (125) and CMS (126), were purified from C. militaris and demonstrated lipid-lowering activity. CM1 was composed of mannose, glucose and galactose at a 1.4:1.0:1.2 molar ratio, and CMS consisted of glucose. Their molecular weights were 700 kDa and 18.2 kDa, respectively. CM1 mainly consisted of (1 → 4)-β-D-Glcp and (1 → 2)-α-D-Manp residue, and CMS was a homopolysaccharide with → 6)Glcp(1 → linkage. Both CM1 and CMS improved [3H]-cholesterol effusion from macrophages to the medium in a dose-dependent manner (0–100 μg/mL) [89]. A Se-polysaccharide (SeCPS-II) (127) was isolated from selenium-enriched C. gunnii. Its molecular weight was 4.12 × 103 kDa, and it consisted of α-L-rhamnose, α-D-mannose, α-D-glucose and β-D-galactose at a 4.33:12.62:27.50:18.99 molar ratio. The inhibition rates of SeCPS-II against SKOV-3 cells in the low-, medium- and high-dosage groups were 8.14% (p < 0.01), 19.75% (p < 0.01) and 36.40% (p < 0.01), respectively, in antitumor assays in vivo [90]. A new polysaccharide CMPB90-1 (128) with a molecular weight of 5.8 kDa was isolated from cultured C. militaris; it consisted of Gal, Glc and Man at a 3.04:1.00:1.45 molar ratio. Furthermore, CMPB90-1 considerably increased the proliferation of lymphocytes at a concentration of 31.2–500 µg/mL in splenic lymphocyte proliferation assays and enhanced the killing effect of NK cells on splenocytes in vitro. It also reinforced the phagocytosis influence of macrophages and induced the M1 polarization of the macrophages [91].
A polysaccharide CP2-S (129) was purified from C. militaris, with a molecular weight of 5.938 × 103 kDa and consisted of glucose. CP2-S significantly stimulated macrophages to take up neutral red, produce NO and increased the excretion of the cytokines IL-1β and IL-6 (50–500 μg/mL) [92]. A protein-polysaccharide HS002-II (130) was fractionated from Hirsutella sinensis and its average molecular weight was 44 kDa. HS002-II was composed of 57.9% polysaccharide and 42.1% protein and was linked by N-type carbohydrate–protein linkages. It consisted of (1 → 3)-linked α-D-ribofuranosyl units, (1 → 4)-linked α-D-xylopyr-anosyl units and (1 → 4)-linked β-D-glucopyranosyl units. Furthermore, HS002-II induced the expression of pro-inflammatory cytokines TNF-α in the upper clear liquid and IL-1β, NF-κB, TNF-α and iNOS in the transcription level in a concentration-dependent manner (0–2.2 μM) [93]. A new polysaccharide (CM-S) (131) was extracted from the fruiting bodies of C. militaris. Its molecular weight was 134,631 Da. CM-S consisted of galactose, glucose and xylose at a 3:2:1 molar ratio and its main chain was (1 → 6)-α-d-galactose. In addition, CM-S considerably increased the proliferation of T cells in contrast to the blank control group at 5, 10 and 20 μg/mL [94]. The information of polysaccharides from Cordyceps fungi was shown in Table 1.
9. Conclusions
Cordyceps sinensis, a famous and precious traditional Chinese medicine, has various activities, including antitussive, asthma-relieving, immune regulation, antibacterial and antitumor, and has a medicinal history of more than 200 years [1]. Cordycepin, isolated and purified from Cordyceps, has also become a focus of research because of its various biological activities. The present review summarizes compounds obtained from Cordyceps with new structures or new activities reported from 2007 to 2022. These compounds include nucleosides, non-ribosomal peptides and alkaloids, polyketides and polysaccharides, among others, which show antitumor, antioxidant, antibacterial, hypoglycemic and immune regulation enhancement activities. Furthermore, researchers have also obtained metabolites from Cordyceps fungi in different manners. For example, four novel isoflavone methy-lglycosides, daidzein 7-O-β-D-glucoside 4″-O-methylate (49), glycitein 7-O-β-D-glucoside 4″-O-methylate (50), genistein 7-O-β-D-glucoside 4″-O-methylate (51), and genistein 4′-O-β-D-glucoside 4″-O-methylate (52) were obtained from C. militaris grown on germinated soybeans [46]; two new polyketides cordypyrones A (68) and B (69) were obtained by the heterologous expression of the gene cluster in Aspergillus nidulans [53]; and an acidic exopolysaccharide AESP-II (110) was isolated on the basis of the immune activity of the fermentation broth of C. militaris [76]. In addition, the paper briefly concludes new activities or new mechanisms of cordycepin that have been reported in recent years. By summarizing the compounds with the new structures and new activities of the known metabolites obtained from the genus Cordyceps in the past 15 years, this paper provides a theoretical basis for research on the active compounds of the genus Cordyceps.
10. Prospects
Cordyceps fungi have always attracted scientific attention due to their various biological activities; however, effectively isolating active monomer compounds from them has been challenging. Under laboratory conditions, fungi produce far fewer compounds than they do under natural conditions [99], and through the analysis of fungal genome data and bioinformatics, fungi have a number of biosynthetic gene clusters of active natural products; however, more than 90% are silent [100]. Cordyceps A (68) and B (69) are novel compounds obtained by heterologously expressing the gene cluster of C. militaris in Aspergillus nidulans. Therefore, by the method of heterologous expression, the biosynthetic gene cluster of Cordyceps fungi can be activated and expressed in filamentous fungi, which can also be used as a potential method for obtaining active compounds in Cordyceps fungi. In addition, in recent years some important molecular technologies have been developed in the deep mining of fungal natural products, such as obtaining specific or non-specific target products through molecular genetic manipulation and directly reconstituting the biosynthesis pathway of target compounds in engineered strains to obtain target compounds. The progress of these methods provides technical support for the research of natural products of Cordyceps fungi [101]. The research on the metabolites and their activities of Cordyceps fungi has been ongoing, and the most in-depth research on the activity and action mechanism has been carried out on cordycepin. In particular, important progress has been made in research on the mechanism of the anti-tumor, immunosuppression and neuroprotective effects of cordycepin [21,22,23,28]. Cordycepin has been clinically studied in multiple clinical settings worldwide as a potential anti-leukemia/anti-cancer chemotherapeutic agent and has passed clinical phase 1 and 2 (clinical trials NCT00003005 (https://clinicaltrials.gov/ct2/show/NCT00003005, accessed on 3 September 2004) and NCT00709215 (https://clinicaltrials.gov/ct2/show/NCT00709215, accessed on 3 July 2008)). Therefore, there is still great research potential and mining value for other active compounds of Cordyceps fungi.
Author Contributions
Conceptualization, P.-J.Z.; writing—original draft preparation, S.-L.Q., S.-S.L., D.L. and P.-J.Z.; writing—review and editing, S.-L.Q. and P.-J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31970060, and 31860015) and the Department of Science and Technology of Yunnan Province (202001BB050061).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, S.P.; Yang, F.Q.; Tsim, K.W.K. Quality control of Cordyceps sinensis, a valued traditional Chinese medicine. J. Pharm. Biomed. 2006, 41, 1571–1584. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Zhang, L.; Wang, J.N.; Wang, W.W.; Niyati, N.; Guo, Y.L.; Wang, X.F. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef]
- Yue, K.; Ye, M.; Zhou, Z.J.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2013, 65, 474–493. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.L.; Xiao, G.H.; Xiong, C.H.; Hu, X.; Zhang, S.W.; Zheng, H.J.; Huang, Y.; Zhou, Y.; Wang, S.Y.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, Y.; Moriyama, M.; Lukasik, P.; Vanderpool, D.; Tanahashi, M.; Meng, X.Y.; McCutcheon, J.P.; Fukatsu, T. Recurrent symbiont recruitment from fungal parasites in cicadas. Proc. Natl. Acad. Sci. USA 2018, 115, E5970–E5979. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.W.; Gong, Z.H.; Su, Y.; Lin, J.; Tang, K.X. Cordyceps fungi: Natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Tang, J.; Tola, A.; Auberon, F.; Oluwaniyi, O.; Ouyang, Z. The genus Cordyceps: An extensive review of its traditional uses, phytochemistry and pharmacology. Fitoterapia 2018, 129, 293–316. [Google Scholar] [CrossRef]
- Chen, B.; Sun, Y.L.; Luo, F.F.; Wang, C.S. Bioactive metabolites and potential mycotoxins produced by Cordyceps fungi: A review of safety. Toxins 2020, 12, 410. [Google Scholar] [CrossRef]
- Yang, M.L.; Kuo, P.C.; Hwang, T.L.; Wu, T.S. Anti-inflammatory principles from Cordyceps sinensis. J. Nat. Prod. 2011, 74, 1996–2000. [Google Scholar] [CrossRef]
- Xue, Y.F.; Wu, L.L.; Ding, Y.L.; Cui, X.M.; Han, Z.Z.; Xu, H. A new nucleoside and two new pyrrole alkaloid derivatives from Cordyceps militaris. Nat. Prod. Res. 2020, 34, 341–350. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, J.; Choi, J.; Hirai, H.; Nishida, H.; Kawagishi, H. Cytotoxic compounds against cancer cells from Bombyx mori inoculated with Cordyceps militaris. Biosci. Biotechnol. Biochem. 2017, 81, 1224–1226. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Liu, C.; Xue, F.; Wu, W.; Wu, X.; Zhang, J.; Fu, J. Effects of cordycepin combined with doxorubicin on proliferation and metastasis of breast cancer cells. Mygosystema 2021, 40, 3012–3022. [Google Scholar] [CrossRef]
- Xiang, T.; Xia, C.; Liu, J.; Wang, C.; Shen, J. Separation, structural identification and anti-tumor effects of new compounds from Cordyceps militaris. Food Sci. 2021, 42, 235–242. [Google Scholar] [CrossRef]
- Sun, J.; Jin, M.; Zhou, W.; Diao, S.; Zhou, Y.; Li, S.; Wang, X.; Pan, S.; Jin, X.; Li, G. A new ribonucleotide from Cordyceps militaris. Nat. Prod. Res. 2017, 31, 2537–2543. [Google Scholar] [CrossRef]
- Kim, S.B.; Ahn, B.; Kim, M.; Ji, H.J.; Shin, S.K.; Hong, I.P.; Kim, C.Y.; Hwang, B.Y.; Lee, M.K. Effect of Cordyceps militaris extract and active constituents on metabolic parameters of obesity induced by high-fat diet in C58BL/6J mice. J. Ethnopharmacol. 2014, 151, 478–484. [Google Scholar] [CrossRef]
- Chen, L.H.; Yang, M.J.; Guan, Y.M.; Zhu, W.F.; Huang, H.L. Identification of nucleosides and nucleobases from cultured Cordyceps militaris. Nat. Prod. Commun. 2014, 9, 637–638. [Google Scholar] [CrossRef] [Green Version]
- Bibi, S.; Hasan, M.M.; Wang, Y.B.; Papadakos, S.P.; Yu, H. Cordycepin as a promising inhibitor of SARS-CoV-2 RNA dependent RNA polymerase (RdRp). Curr. Med. Chem. 2022, 29, 152–162. [Google Scholar] [CrossRef]
- Rabie, A.M. Potent inhibitory activities of the adenosine analogue cordycepin on SARS-CoV-2 replication. ACS Omega 2022, 7, 2960–2969. [Google Scholar] [CrossRef]
- Panya, A.; Songprakhon, P.; Panwong, S.; Jantakee, K.; Kaewkod, T.; Tragoolpua, Y.; Sawasdee, N.; Lee, V.S.; Nimmanpipug, P.; Yenchitsomanus, P.T. Cordycepin inhibits virus replication in dengue virus-infected vero cells. Molecules 2021, 26, 3118. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z. Cordycepin protects PC12 cells against 6-hydroxydopamine induced neurotoxicity via its antioxidant properties. Biomed. Pharmacother. 2016, 81, 7–14. [Google Scholar] [CrossRef]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Oh, J.I.; Lee, S.J.; Park, G. The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells. Neurotoxicology 2014, 41, 102–111. [Google Scholar] [CrossRef]
- Dong, Z.S.; Cao, Z.P.; Shang, Y.J.; Liu, Q.Y.; Wu, B.Y.; Liu, W.X.; Li, C.H. Neuroprotection of cordycepin in NMDA-induced excitotoxicity by modulating adenosine A1 receptors. Eur. J. Pharmacol. 2019, 853, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M. Cordycepin in anticancer research: Molecular mechanism of therapeutic effects. Curr. Med. Chem. 2020, 27, 983–996. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.Y.; Park, S.J.; Park, Y.J. The anticancer properties of cordycepin and their underlying mechanisms. Int. J. Mol. Sci. 2018, 19, 27. [Google Scholar] [CrossRef] [Green Version]
- Tania, M.; Shawon, J.; Saif, K.; Kiefer, R.; Khorram, M.S.; Halim, M.A.; Khan, M.A. Cordycepin downregulates Cdk-2 to interfere with cell cycle and increases apoptosis by generating ROS in cervical cancer cells: In vitro and in silico study. Curr. Cancer Drug Target 2019, 19, 152–159. [Google Scholar] [CrossRef]
- Chang, M.M.; Hong, S.Y.; Yang, S.H.; Wu, C.C.; Wang, C.Y.; Huang, B.M. Anti-cancer effect of cordycepin on FGF9-induced testicular tumorigenesis. Int. J. Mol. Sci. 2020, 21, 8336. [Google Scholar] [CrossRef]
- Liu, C.Y.; Qi, M.; Li, L.; Yuan, Y.; Wu, X.P.; Fu, J.S. Natural cordycepin induces apoptosis and suppresses metastasis in breast cancer cells by inhibiting the Hedgehog pathway. Food Funct. 2020, 11, 2107–2116. [Google Scholar] [CrossRef]
- Chang, M.M.; Pan, B.S.; Wang, C.Y.; Huang, B.M. Cordycepin-induced unfolded protein response-dependent cell death, and AKT/MAPK-mediated drug resistance in mouse testicular tumor cells. Cancer Med. 2019, 8, 3949–3964. [Google Scholar] [CrossRef] [Green Version]
- Hama, M.; Elshamy, A.I.; Yoneyama, T.; Kasai, Y.; Yamamoto, H.; Tanigawa, K.; Oshiro, A.; Noji, M.; Ban, S.; Imagawa, H.; et al. New alkaloidal metabolites from cultures of entomopathogenic fungus Cordyceps takaomontana NBRC 101754. Fitoterapia 2019, 139, 104364. [Google Scholar] [CrossRef]
- Fan, W.; Li, E.; Ren, J.; Wang, W.; Liu, X.; Zhang, Y. Cordycepamides A-E and cordyglycoside A, new alkaloidal and glycoside metabolites from the entomopathogenic fungus Cordyceps sp. Fitoterapia 2020, 142, 104525. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, J.; Wei, L.; Shi, M.; Wang, J.; Huang, J. Gunnilactams A-C, macrocyclic tetralactams from the mycelial culture of the entomogenous fungus Paecilomyces gunnii. J. Nat. Prod. 2017, 80, 1935–1938. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Chen, C.; Zou, Y.; Lou, H.; Zheng, Q.; Guo, L.; Lin, J.; Ye, Z.; Yun, F. Purification and structural characterization of a novel natural pigment: Cordycepene from edible and medicinal mushroom Cordyceps militaris. Appl. Microbiol. Biotechnol. 2019, 103, 7943–7952. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.M.; Tao, H.H.; Feng, B.M. Cordyceamides A and B from the culture liquid of Cordyceps sinensis (BERK.) SACC. Chem. Pharm. Bull. 2009, 57, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xu, J.; Wang, S.; Hou, Z.; Lu, X.; An, L.; Du, P. A new cerebroside from Cordyceps militaris with anti-PTP1B activity. Fitoterapia 2019, 138, 104342. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Chinthanom, P.; Rachtawee, P.; Somyong, W.; Luangsa-ard, J.J.; Hywel-Jones, N.L. Cordylactam, a new alkaloid from the spider pathogenic fungus Cordyceps sp BCC 12671. Phytochem. Lett. 2013, 6, 162–164. [Google Scholar] [CrossRef]
- Umeyama, A.; Takahashi, K.; Grudniewska, A.; Shimizu, M.; Hayashi, S.; Kato, M.; Okamoto, Y.; Suenaga, M.; Ban, S.; Kumada, T.; et al. In vitro antitrypanosomal activity of the cyclodepsipeptides, cardinalisamides A-C, from the insect pathogenic fungus Cordyceps cardinalis NBRC 103832. J. Antibiot. 2014, 67, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, D.M.; Jia, J.F.; Peng, Q.L.; Tian, H.Y.; Wang, L.; Ye, W.C. Cyclodepsipeptides from the ascocarps and insect-body portions of fungus Cordyceps cicadae. Fitoterapia 2014, 97, 23–27. [Google Scholar] [CrossRef]
- Haritakun, R.; Sappan, M.; Suvannakad, R.; Tasanathai, K.; Isaka, M. An antimycobacterial cyclodepsipeptide from the entomopathogenic fungus Ophiocordyceps communis BCC 16475. J. Nat. Prod. 2010, 73, 75–78. [Google Scholar] [CrossRef]
- Helaly, S.E.; Kuephadungphan, W.; Phainuphong, P.; Ibrahim, M.A.A.; Tasanathai, K.; Mongkolsamrit, S.; Luangsa-ard, J.J.; Phongpaichit, S.; Rukachaisirikul, V.; Stadler, M. Pigmentosins from Gibellula sp. as antibiofilm agents and a new glycosylated asperfuran from Cordyceps javanica. Beilstein J. Org. Chem. 2019, 15, 2968–2981. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L. Study on the Structure of Sedative and Hypnotic Active Substances of Paecilomyces gounisii. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2012. [Google Scholar]
- Yuan, G.; An, L.; Sun, Y.; Xu, G.; Du, P. Improvement of learning and memory induced by Cordyceps polypeptide treatment and the underlying mechanism. Evid. Based Complement. Alternat. Med. 2018, 2018, 9419264. [Google Scholar] [CrossRef]
- Lu, R.; Liu, X.; Gao, S.; Zhang, W.; Peng, F.; Hu, F.; Huang, B.; Chen, L.; Bao, G.; Li, C.; et al. New tyrosinase inhibitors from Paecilomyces gunnii. J. Agric. Food Chem. 2014, 62, 11917–11923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Xuan, L.J. Cyclopentenone and furan derivative from the mycelia of Cordyceps cicadae. J. Antibiot. 2008, 61, 43–45. [Google Scholar] [CrossRef]
- Lu, R.L.; Bao, G.H.; Hu, F.L.; Huang, B.; Li, C.R.; Li, Z.Z. Comparison of cytotoxic extracts from fruiting bodies, infected insects and cultured mycelia of Cordyceps formosana. Food Chem. 2014, 145, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Grudniewska, A.; Hayashi, S.; Shimizu, M.; Kato, M.; Suenaga, M.; Imagawa, H.; Ito, T.; Asakawa, Y.; Ban, S.; Kumada, T.; et al. Opaliferin, a new polyketide from cultures of entomopathogenic fungus Cordyceps sp. NBRC 106954. Org. Lett. 2014, 16, 4695–4697. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.N.; Kim, J.; Lee, M.Y.; Park, D.K.; Hong, Y.S.; Lee, C.H. Metabolomics revealed novel isoflavones and optimal cultivation time of Cordyceps militaris fermentation. J. Agric. Food Chem. 2010, 58, 4258–4267. [Google Scholar] [CrossRef]
- Wei, J.; Zhou, X.; Dong, M.; Yang, L.; Zhao, C.; Lu, R.; Bao, G.; Hu, F. Metabolites and novel compounds with anti-microbial or antiaging activities from Cordyceps fumosorosea. AMB Express 2022, 12, 40. [Google Scholar] [CrossRef]
- Yang, N.-N.; Jiang, N.; Ma, Q.-Y.; Kong, F.-D.; Xie, Q.-Y.; Zhou, L.-M.; Yu, Z.-F.; Zhao, Y.-X. Chemical study of the strain Cordyceps spp. from cell fusion between Cordyceps militaris and Cordyceps cicadae. J. Asian Nat. Prod. Res. 2019, 21, 449–455. [Google Scholar] [CrossRef]
- Ojima, K.-i.; Yangchum, A.; Laksanacharoen, P.; Tasanathai, K.; Thanakitpipattana, D.; Tokuyama, H.; Isaka, M. Cordybislactone, a stereoisomer of the 14-membered bislactone clonostachydiol, from the hopper pathogenic fungus Cordyceps sp BCC 49294: Revision of the absolute configuration of clonostachydiol. J. Antibiot. 2018, 71, 351–358. [Google Scholar] [CrossRef]
- Wang, M.; Kornsakulkarn, J.; Srichomthong, K.; Feng, T.; Liu, J.-K.; Isaka, M.; Thongpanchang, C. Antimicrobial anthraquinones from cultures of the ant pathogenic fungus Cordyceps morakotii BCC 56811. J. Antibiot. 2019, 72, 141–147. [Google Scholar] [CrossRef]
- Yoon, D.H.; Lim, M.-H.; Lee, Y.R.; Sung, G.-H.; Lee, T.-H.; Jeon, B.H.; Cho, J.Y.; Song, W.O.; Park, H.; Choi, S.; et al. A novel synthetic analog of militarin, MA-1 induces mitochondrial dependent apoptosis by ROS generation in human lung cancer cells. Toxicol. Appl. Pharmacol. 2013, 273, 659–671. [Google Scholar] [CrossRef]
- Jiang, Y.; Wong, J.H.; Fu, M.; Ng, T.B.; Liu, Z.K.; Wang, C.R.; Li, N.; Qiao, W.T.; Wen, T.Y.; Liu, F. Isolation of adenosine, iso-sinensetin and dimethylguanosine with antioxidant and HIV-1 protease inhibiting activities from fruiting bodies of Cordyceps militaris. Phytomedicine 2011, 18, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-L.; Yu, C.; Li, L. Heterologous expression of a natural product biosynthetic gene cluster from Cordyceps militaris. J. Antibiot. 2022, 75, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, Y.; Wu, Y.; Zhang, X.; Jiang, L.; Zhang, Y. Bioassay-guided separation and identification of a new anti-lung cancer compound from Cordyceps militaris by means of off-line two-dimensional preparative chromatography, real-time cell analysis, and X-ray single-crystal diffraction. Chromatographia 2015, 78, 495–506. [Google Scholar] [CrossRef]
- Dong, J.Z.; Wang, S.H.; Ai, X.R.; Yao, L.; Sun, Z.W.; Lei, C.; Wang, Y.; Wang, Q. Composition and characterization of cordyxanthins from Cordyceps militaris fruit bodies. J. Funct. Food. 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Zhang, S.W.; Xuan, L.J. Five aromatics bearing a 4-O-methylglucose unit from Cordyceps cicadae. Helv. Chim. Acta 2007, 90, 404–410. [Google Scholar] [CrossRef]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Intereya, K.; Kocharin, K. New diphenyl ethers from the insect pathogenic fungus Cordyceps sp. BCC 1861. Chem. Pharm. Bull. 2007, 55, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Asai, T.; Luo, D.; Obara, Y.; Taniguchi, T.; Monde, K.; Yamashita, K.; Oshima, Y. Dihydrobenzofurans as cannabinoid receptor ligands from Cordyceps annullata, an entomopathogenic fungus cultivated in the presence of an HDAC inhibitor. Tetrahedron Lett. 2012, 53, 2239–2243. [Google Scholar] [CrossRef]
- Yang, Q.; Yin, Y.; Yu, G.; Jin, Y.; Ye, X.; Shrestha, A.; Liu, W.; Yu, W.; Sun, H. A novel protein with anti-metastasis activity on 4T1 carcinoma from medicinal fungus Cordyceps militaris. Int. J. Biol. Macromol. 2015, 80, 385–391. [Google Scholar] [CrossRef]
- Bai, K.C.; Sheu, F. A novel protein from edible fungi Cordyceps militaris that induces apoptosis. J. Food Drug Anal. 2018, 26, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y.; Chen, X.; Siu, K.C. Isolation and structure characterization of an antioxidative glycopeptide from mycelial culture broth of a medicinal fungus. Int. J. Mol. Sci. 2014, 15, 17318–17332. [Google Scholar] [CrossRef] [Green Version]
- Park, B.T.; Na, K.H.; Jung, E.C.; Park, J.W.; Kim, H.H. Antifungal and anticancer activities of a protein from the mushroom Cordyceps militaris. Korean J. Physiol. Pharmacol. 2009, 13, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Wu, Q.; Wang, J.; Li, Y.; Chen, B.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Xie, Y.; et al. Novel selenium peptides obtained from selenium-enriched Cordyceps militaris alleviate neuroinflammation and gut microbiota dysbacteriosis in LPS-injured mice. J. Agric. Food Chem. 2022, 70, 3194–3206. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.W.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine 2011, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Bi, B.; Wang, X.; Wu, H.; Wei, Q. Purification and characterisation of a novel protease from Cordyceps sinensis and determination of the cleavage site motifs using oriented peptide library mixtures. Food Chem. 2011, 126, 46–53. [Google Scholar] [CrossRef]
- Cui, L.; Dong, M.S.; Chen, X.H.; Jiang, M.; Lv, X.; Yan, G. A novel fibrinolytic enzyme from Cordyceps militaris, a Chinese traditional medicinal mushroom. World J. Microbiol. Biotechnol. 2008, 24, 483–489. [Google Scholar] [CrossRef]
- FengLin, H.U.; ZengZhi, L.I. Secondary metabolites and their bioactivities of Cordyceps and its related fungi. Mygosystema 2007, 26, 607–632. [Google Scholar]
- Yan, J.K.; Wang, W.Q.; Ma, H.L.; Wu, J.Y. Sulfation and enhanced antioxidant capacity of an exopolysaccharide produced by the medicinal fungus Cordyceps sinensis. Molecules 2012, 18, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zeng, Y.; Cui, Y.; Liu, H.; Dong, C.; Sun, Y. Structural characterization, antioxidant and immunomodulatory activities of a neutral polysaccharide from Cordyceps militaris cultivated on hull-less barley. Carbohydr. Polym. 2020, 235, 115969. [Google Scholar] [CrossRef]
- Jing, Y.; Cui, X.; Chen, Z.; Huang, L.; Song, L.; Liu, T.; Lv, W.; Yu, R. Elucidation and biological activities of a new polysaccharide from cultured Cordyceps militaris. Carbohydr. Polym. 2014, 102, 288–296. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Yang, X.; Lin, P.; Liu, N.; Li, X.; Zhang, B.; Guo, S. Purification, structural characterization, and PCSK9 secretion inhibitory effect of the novel alkali-extracted polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2021, 179, 407–417. [Google Scholar] [CrossRef]
- Sun, H.; Yu, X.; Li, T.; Zhu, Z. Structure and hypoglycemic activity of a novel exopolysaccharide of Cordyceps militaris. Int. J. Biol. Macromol. 2021, 166, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lin, P.; Wang, J.; Liu, N.; Yin, F.; Shen, N.; Guo, S. Purification, characterization and anti-atherosclerotic effects of the polysaccharides from the fruiting body of Cordyceps militaris. Int. J. Biol. Macromol. 2021, 181, 890–904. [Google Scholar] [CrossRef]
- Shi, K.; Yang, G.; He, L.; Yang, B.; Li, Q.; Yi, S. Purification, characterization, antioxidant, and antitumor activity of polysaccharides isolated from silkworm Cordyceps. J. Food Biochem. 2020, 44, e13482. [Google Scholar] [CrossRef] [PubMed]
- He, B.L.; Zheng, Q.W.; Guo, L.Q.; Huang, J.Y.; Yun, F.; Huang, S.S.; Lin, J.F. Structural characterization and immune-enhancing activity of a novel high-molecular-weight polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2020, 145, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wen, Q.; Song, A.; Liu, Y.; Wang, F.; Jiang, B. Isolation and immune activity of a new acidic Cordyceps militaris exopolysaccharide. Int. J. Biol. Macromol. 2022, 194, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Siu, K.C.; Wang, W.Q.; Liu, X.X.; Wu, J.Y. Structure and antioxidant activity of a novel poly-N-acetylhexosamine produced by a medicinal fungus. Carbohydr. Polym. 2013, 94, 332–338. [Google Scholar] [CrossRef]
- Jing, Y.; Zhu, J.; Liu, T.; Bi, S.; Hu, X.; Chen, Z.; Song, L.; Lv, W.; Yu, R. Structural characterization and biological activities of a novel polysaccharide from cultured Cordyceps militaris and its sulfated derivative. J. Agric. Food Chem. 2015, 63, 3464–3471. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Yin, Q.S.; Han, T.; Zhao, Y.X.; Su, J.J.; Li, M.Z.; Ling, J.Y. Purification and antioxidant effect of novel fungal polysaccharides from the stroma of Cordyceps kyushuensis. Ind. Crops Prod. 2015, 69, 485–491. [Google Scholar] [CrossRef]
- Mei, Y.X.; Yang, W.; Zhu, P.X.; Peng, N.; Zhu, H.; Liang, Y.X. Isolation, characterization, and antitumor activity of a novel heteroglycan from cultured mycelia of Cordyceps sinensis. Planta Med. 2014, 80, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.J.; Chang, N.C.; Jayakumar, T.; Liao, J.C.; Lin, M.J.; Wang, S.H.; Chou, D.S.; Thomas, P.A.; Sheu, J.R. Ex vivo and in vivo studies of CME-1, a novel polysaccharide purified from the mycelia of Cordyceps sinensis that inhibits human platelet activation by activating adenylate cyclase/cyclic AMP. Thromb. Res. 2014, 134, 1301–1310. [Google Scholar] [CrossRef]
- Yan, J.-K.; Wang, W.-Q.; Li, L.; Wu, J.-Y. Physiochemical properties and antitumor activities of two α-glucans isolated from hot water and alkaline extracts of Cordyceps (Cs-HK1) fungal mycelia. Carbohydr. Polym. 2011, 85, 753–758. [Google Scholar] [CrossRef]
- Wang, Z.M.; Peng, X.A.; Lee, K.L.D.; Tang, J.C.O.; Cheung, P.C.K.; Wu, J.Y. Structural characterisation and immunomodulatory property of an acidic polysaccharide from mycelial culture of Cordyceps sinensis fungus Cs-HK1. Food Chem. 2011, 125, 637–643. [Google Scholar] [CrossRef]
- Zhang, A.L.; Lu, J.H.; Zhang, N.; Zheng, D.; Zhang, G.R.; Teng, L.R. Extraction, purification and anti-tumor activity of polysaccharide from mycelium of mutant Cordyceps militaris. Chem. Res. Chin. Univ. 2010, 26, 798–802. [Google Scholar]
- Cheung, J.K.H.; Li, J.; Cheung, A.W.H.; Zhu, Y.; Zheng, K.Y.Z.; Bi, C.W.C.; Duan, R.; Choi, R.C.Y.; Lau, D.T.W.; Dong, T.T.X.; et al. Cordysinocan, a polysaccharide isolated from cultured Cordyceps, activates immune responses in cultured T-lymphocytes and macrophages: Signaling cascade and induction of cytokines. J. Ethnopharmacol. 2009, 124, 61–68. [Google Scholar] [CrossRef]
- Yu, R.M.; Yin, Y.; Yang, W.; Ma, W.L.; Yang, L.; Chen, X.J.; Zhang, Z.; Ye, B.; Song, L.Y. Structural elucidation and biological activity of a novel polysaccharide by alkaline extraction from cultured Cordyceps militaris. Carbohydr. Polym. 2009, 75, 166–171. [Google Scholar] [CrossRef]
- Trung Hieu, L.; Thi Van Thi, T.; Van Khoa, T.; Xuan Anh Vu, H.; Thanh Minh, T.; Dang Giang Chau, N.; Thi Hong Chuong, N.; Varma, R.S.; Tam Kiet, T.; Thanh-Tam, H.; et al. Structural characterization of mannoglucan isolated from Ophiocordyceps sobolifera and its antioxidant activities. ACS Omega 2022, 7, 9397–9405. [Google Scholar] [CrossRef]
- Tran Thi Van, T.; Tran Van, K.; Ho Xuan Anh, V.; Le Lam, S.; Le Trung, H.; Nguyen Thi Hong, C.; Nguyen Chinh, C.; Kim, S.Y.; Le Quyet, V. Chemical structure of a novel heteroglycan polysaccharide isolated from the biomass of Ophiocordyceps Sobolifera. J. Mol. Struct. 2021, 1232, 129986. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Li, F.; Hou, P.; Yin, J.; Yang, Z.; Yang, X.; Li, T.; Xia, B.; Zhou, G.; et al. Structural characterisation and cholesterol efflux improving capacity of the novel polysaccharides from Cordyceps militaris. Int. J. Biol. Macromol. 2019, 131, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhu, Z.; Tang, Y.; Ren, Y.; Song, Q.; Tang, Y.; Zhang, Y. Structural characterization and antitumor activity of a novel Se-polysaccharide from seleniumenriched Cordyceps gunnii. Food Funct. 2018, 9, 2744–2754. [Google Scholar] [CrossRef]
- Bi, S.; Jing, Y.; Zhou, Q.; Hu, X.; Zhu, J.; Guo, Z.; Song, L.; Yu, R. Structural elucidation and immunostimulatory activity of a new polysaccharide from Cordyceps militaris. Food Funct. 2018, 9, 279–293. [Google Scholar] [CrossRef]
- Zhu, L.; Tang, Q.; Zhou, S.; Liu, Y.; Zhang, Z.; Gao, X.; Wang, S.; Wang, Z. Isolation and purification of a polysaccharide from the caterpillar medicinal mushroom Cordyceps militaris (Ascomycetes) fruit bodies and its immunomodulation of RAW 264.7 macrophages. Int. J. Med. Mushrooms 2014, 16, 247–257. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Ji, P.; Cheng, J.; Wang, Y.; Qian, H.; Li, W.; Gong, X.; Wang, Z. Structural characterization and immunostimulatory activity of a novel protein-bound polysaccharide produced by Hirsutella sinensis Liu, Guo, Yu & Zeng. Food Chem. 2013, 141, 946–953. [Google Scholar] [CrossRef]
- Ding, X.; Song, Z.; Hou, Y. Comparative study on the structure characterization and immune activity of Lactarius vellereus Fr. polysaccharide (LV-1) and Cordyceps militaris (L. ex Fr.) Link. polysaccharide (CM-S). J. Food Meas. Charact. 2022, 16, 901–919. [Google Scholar] [CrossRef]
- Li, X.G.; Pan, W.D.; Lou, H.Y.; Liu, R.M.; Xiao, J.H.; Zhong, J.J. New cytochalasins from medicinal macrofungus Crodyceps taii and their inhibitory activities against human cancer cells. Bioorg. Med. Chem. Lett. 2015, 25, 1823–1826. [Google Scholar] [CrossRef]
- Wei, P.Y.; Liu, L.X.; Liu, T.; Chen, C.; Luo, D.Q.; Shi, B.Z. Three new pigment protein tyrosine phosphatases inhibitors from the insect parasite fungus Cordyceps gracilioides: Terreusinone A, pinophilin C and cryptosporioptide A. Molecules 2015, 20, 5825–5834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, T.; Yamamoto, T.; Chung, Y.-M.; Chang, F.-R.; Wu, Y.-C.; Yamashita, K.; Oshima, Y. Aromatic polyketide glycosides from an entomopathogenic fungus, Cordyceps indigotica. Tetrahedron Lett. 2012, 53, 277–280. [Google Scholar] [CrossRef]
- Asai, T.; Yamamoto, T.; Oshima, Y. Aromatic polyketide production in Cordyceps indigotica, an entomopathogenic fungus, induced by exposure to a histone deacetylase inhibitor. Org. Lett. 2012, 14, 2006–2009. [Google Scholar] [CrossRef]
- Barkal, L.J.; Theberge, A.B.; Guo, C.J.; Spraker, J.; Rappert, L.; Berthier, J.; Brakke, K.A.; Wang, C.C.C.; Beebe, D.J.; Keller, N.P.; et al. Microbial metabolomics in open microscale platforms. Nat. Commun. 2016, 7, 10610. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, H. Reconstitution of biosynthetic machinery of fungal natural products in heterologous hosts. Biosci. Biotech. Bioch. 2020, 84, 433–444. [Google Scholar] [CrossRef]
- Skellam, E. Strategies for engineering natural product biosynthesis in fungi. Trends Biotechnol. 2019, 37, 416–427. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Nucleosides (1–10) from Cordyceps.
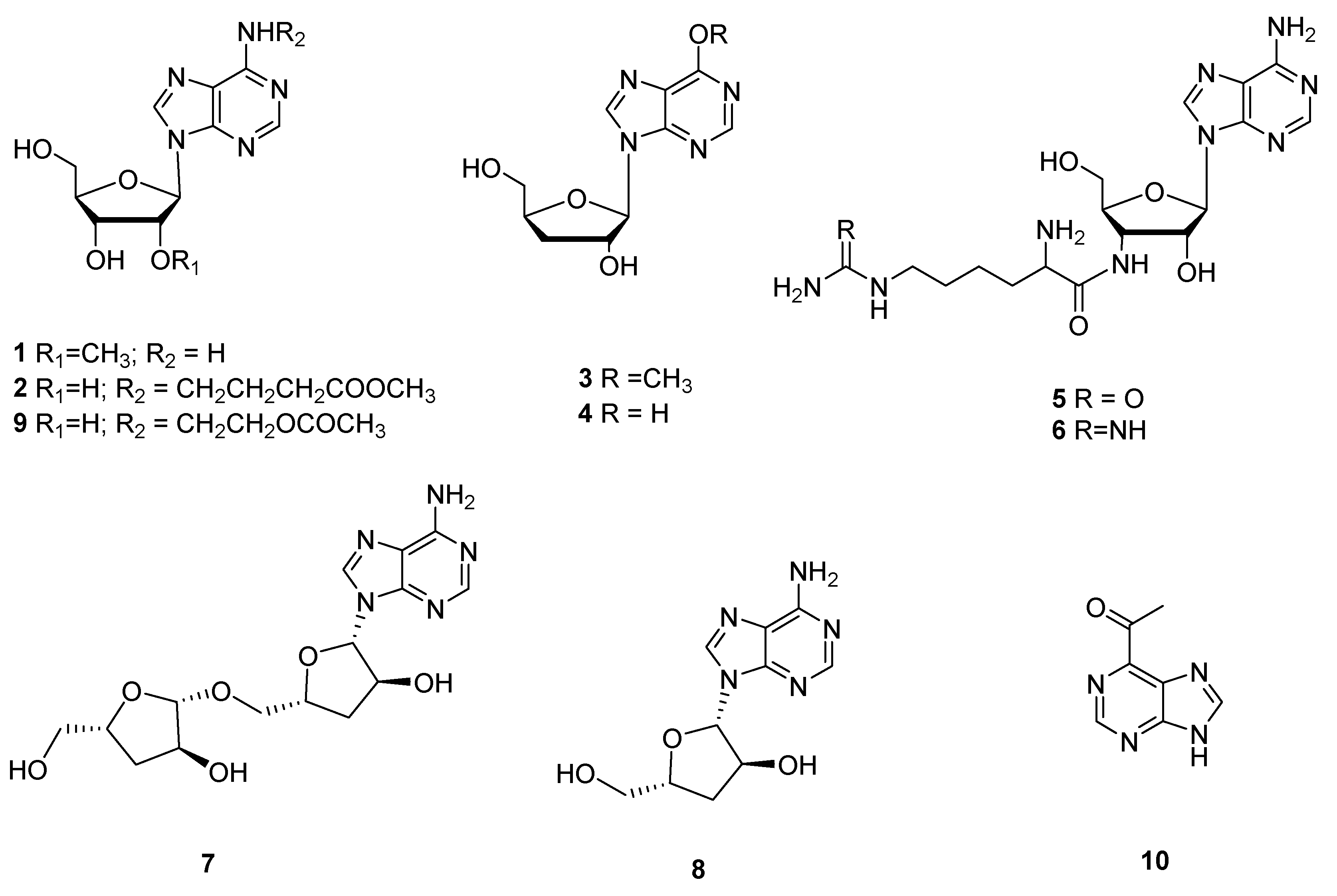
Figure 2.
Non-ribosomal peptides and alkaloids (11–31) from Cordyceps.
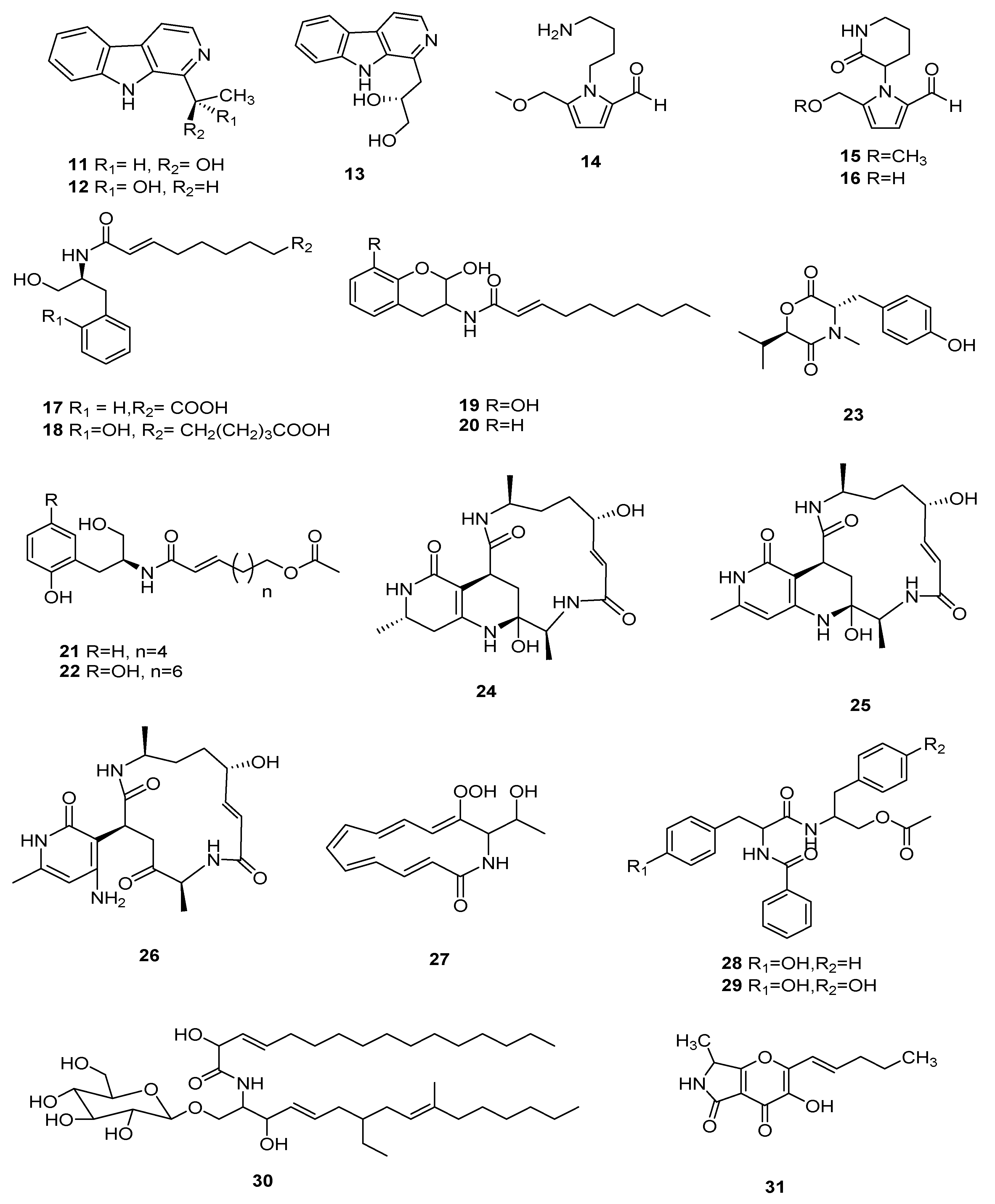
Figure 3.
Non-ribosomal peptides (32–40) from Cordyceps.
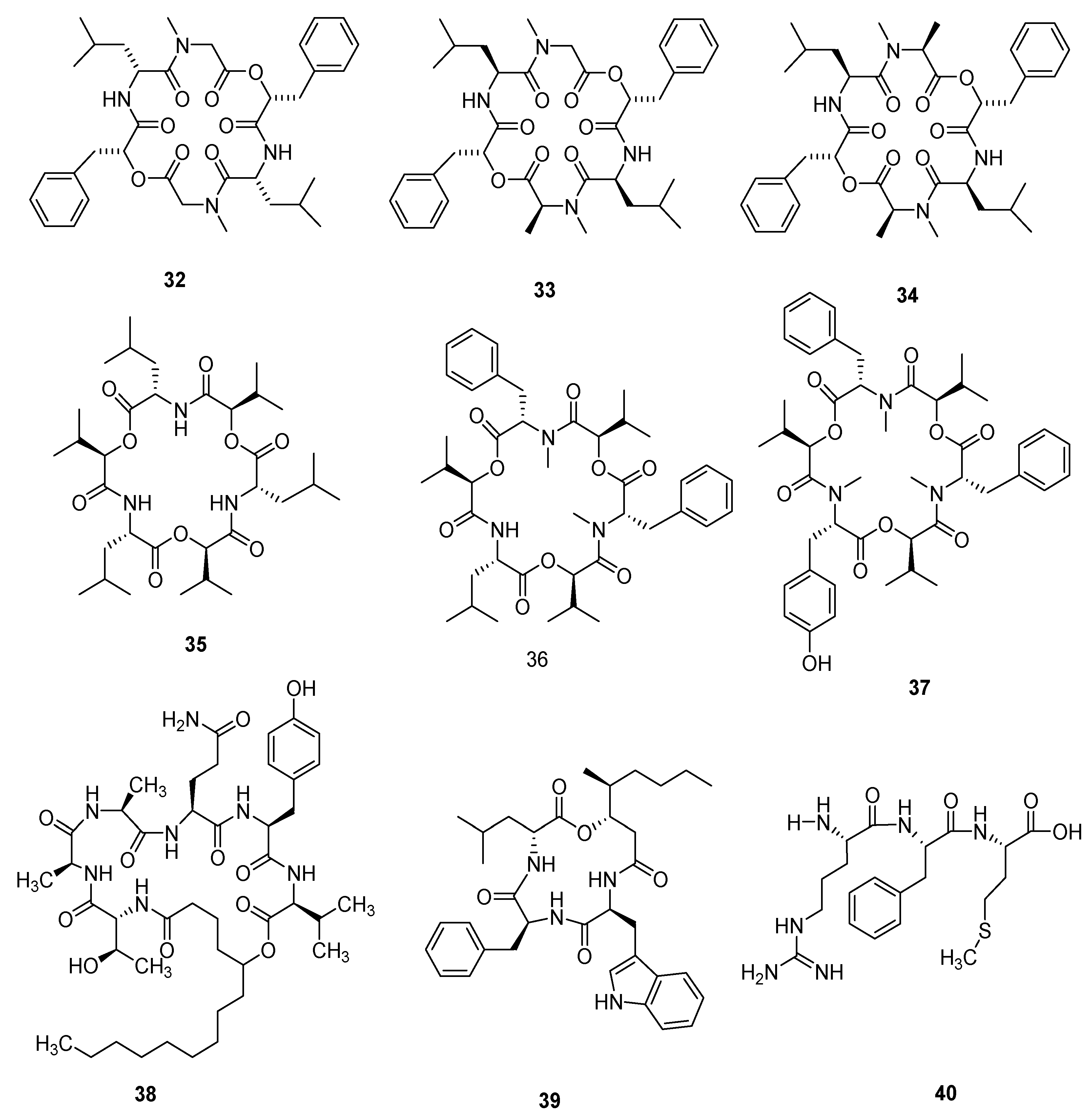
Figure 4.
Polyketides (41–56) from Cordyceps.

Figure 5.
Polyketides (57–69) from Cordyceps.

Figure 6.
Sterols and terpenoids (70–78) from Cordyceps.
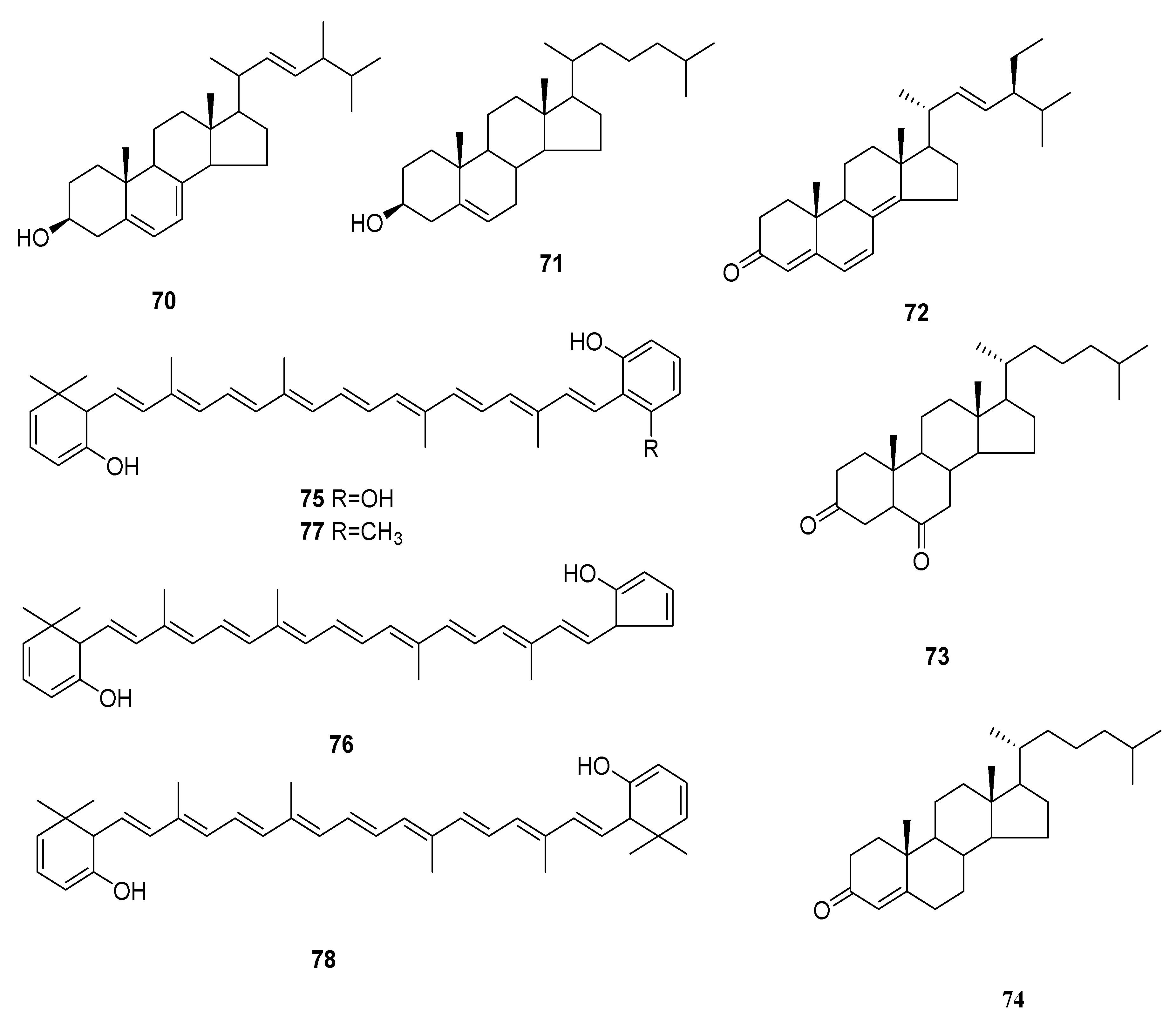
Figure 7.
Aromatics and their derivatives (79–91) from Cordyceps.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Polysaccharides originated from Cordyceps fungi.
| Name | Organism Species | Extraction, Medium | M. W. | Bioactivities | References |
|---|---|---|---|---|---|
| SDQCP-1 | C. militaris | Hot water | 19.3 kDa | Antioxidant and immunomodulatory | [69] |
| CMP-1 | C. militaris | Ultrasound, hot water | 4.3 kDa | Antioxidant, immune-stimulatory and anti-tumor | [70] |
| CM3-SII | C. militaris | Alkaline | - | Potential hypolipidemic | [71] |
| EPS-III | C. militaris | Ethanol, Sevag method | 1.56 × 103 kDa | Hypoglycemic | [72] |
| CM3 II | C. militaris | Alkaline | - | Anti-atherosclerotic | [73] |
| SCP II-1 | Silkworm Cordyceps sp. | Hot water | 35.2 kDa | Antioxidant and antitumor | [74] |
| SCP II-2 | Silkworm Cordyceps sp. | Hot water | 23.4 kDa | Antioxidant | [74] |
| CMP-III | C. militaris | Ultrasound, hot water | 4.796× 104 kDa | Immunomodulatory | [75] |
| AESP-II | C. militaris | Ethanol, Sevag method | 61.52 kDa | Immunomodulatory | [76] |
| PolyhexNAc | C. sinensis | Ethanol | 6 kDa | Antioxidant and cytoprotective | [77] |
| CMPA90-1 | C. militaris | Simulated gastric juice, ethanol | - | Antioxidant and anti-tumor | [78] |
| PSCK2-2 | C. kyushuensis | Methanol, hot water, ethanol | - | Antioxidant activity and protective effects against DNA damage | [79] |
| PSCK2-3 | C. kyushuensis | Methanol, hot water, ethanol | - | Antioxidant and protective effects against DNA damage | [79] |
| Cordyceps sinensis polysaccharide 1 | C. sinensis | Hot water, ethanol, Sevag method | 1.17 × 102 kDa | Antitumor | [80] |
| CME-1 | C. sinensis | Double-distilled H2O | 27.6 kDa | Inhibiting human platelet aggregation | [81] |
| WIPS | C. sinensis | Hot water, alkaline | 1.18 × 103 kDa | Antitumor and immunomodulatory | [82] |
| AIPS | C. sinensis | Hot water, alkaline | 1.15 × 103 kDa | Antitumor and immunomodulatory | [82] |
| AEPS-1 | C. sinensis | Ethanol, Sevag method | 36 kDa | Immunomodulatory | [83] |
| MCMP | C. militaris | Hot water, ethanol, Sevag method | 8.1 kDa | Antitumor | [84] |
| Cordysinocan | Cordyceps sp. | Ethanol | 82 kDa | Immunomodulatory | [85] |
| CBP-1 | C. militaris | Ethanol, hot water, alkaline | - | Antioxidant | [86] |
| PS-T80 | O. sobolifera | Ethanol, hot water | 74 kDa | Antioxidant | [87] |
| PS-T100 | O. sobolifera | Hot water, ethanol | 2.29 × 102 kDa | - | [88] |
| CM1 | C. militaris | Ethanol, hot water | 700 kDa | Lipid-lowering | [89] |
| CMS | C. militaris | Ethanol, hot water | 18.2 kDa | Lipid-lowering | [89] |
| SeCPS-II | C. gunnii | Hot water, ethanol, Sevag method | 4.12 × 103 kDa | Antitumor | [90] |
| CMPB90-1 | C. militaris | Water, ethanol, alkaline | 5.8 kDa | Immunomodulatory | [91] |
| CP2-S | C. militaris | Hot water, ethanol | 5.938 × 103 kDa | Immunomodulatory | [92] |
| HS002-II | Hirsutella sinensis | Papain enzymolysis, Sevag method, ethanol | 44 kDa | Immunostimulatory | [93] |
| CM-S | C. militaris | Hot water, ethanol | 134631 Da | Immune | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qu, S.-L.; Li, S.-S.; Li, D.; Zhao, P.-J. Metabolites and Their Bioactivities from the Genus Cordyceps. Microorganisms 2022, 10, 1489. https://doi.org/10.3390/microorganisms10081489
AMA Style
Qu S-L, Li S-S, Li D, Zhao P-J. Metabolites and Their Bioactivities from the Genus Cordyceps. Microorganisms. 2022; 10(8):1489. https://doi.org/10.3390/microorganisms10081489
Chicago/Turabian StyleQu, Shuai-Ling, Su-Su Li, Dong Li, and Pei-Ji Zhao. 2022. "Metabolites and Their Bioactivities from the Genus Cordyceps" Microorganisms 10, no. 8: 1489. https://doi.org/10.3390/microorganisms10081489
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.