

Effects of Dietary Supplementation of Solubles from Shredded, Steam-Exploded Pine Particles on the Performance and Cecum Microbiota of Acute Heat-Stressed Broilers
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Solubles from Shredded, Steam-Exploded Pine Particles (SSPPs)
2.2. Experimental Birds and Housing
2.3. Rectal Temperature
2.4. Sampling and Measurements
2.5. Plasma Analysis
2.6. Cecum DNA Extraction and Metagenome Investigation
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-De-Paula, V.; Pinheiro, M.L.; Sakai, M.; Sá, L.R.; Ferreira, A.J.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Awad, E.A.; Najaa, M.; Zulaikha, Z.A.; Zulkifli, I.; Soleimani, A.F. Effects of heat stress on growth performance, selected physiological and immunological parameters, caecal microflora, and meat quality in two broiler strains. Asian-Australas. J. Anim. Sci. 2020, 33, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Ncho, C.M.; Jeong, C.-M.; Choi, Y.-H. Embryonic Thermal Manipulation and in ovo Gamma-Aminobutyric Acid Supplementation Regulating the Chick Weight and Stress-Related Genes at Hatch. Front. Vet. Sci. 2021, 8, 807450. [Google Scholar] [CrossRef]
- Goo, D.; Kim, J.H.; Park, G.H.; Delos Reyes, J.B.; Kil, D.Y. Effect of Heat Stress and Stocking Density on Growth Performance, Breast Meat Quality, and Intestinal Barrier Function in Broiler Chickens. Animals 2019, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Kucuk, O. Zinc Supplementation Alleviates Heat Stress in Laying Japanese Quail. J. Nutr. 2003, 133, 2808–2811. [Google Scholar] [CrossRef] [PubMed]
- Ncho, C.-M.; Goel, A.; Jeong, C.-M.; Youssouf, M.; Choi, Y.-H. In Ovo Injection of GABA Can Help Body Weight Gain at Hatch, Increase Chick Weight to Egg Weight Ratio, and Improve Broiler Heat Resistance. Animals 2021, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Farnell, Y.Z.; Peebles, E.D.; Kiess, A.S.; Wamsley, K.G.S.; Zhai, W. Effects of prebiotics, probiotics, and their combination on growth performance, small intestine morphology, and resident Lactobacillus of male broilers1. Poult. Sci. 2016, 95, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Okrathok, S.; Khempaka, S. Modified-dietary fiber from cassava pulp reduces abdominal fat and meat cholesterol contents without affecting growth performance of broiler chickens. J. Appl. Poult. Res. 2020, 29, 229–239. [Google Scholar] [CrossRef]
- Urso, U.R.A.; Dahlke, F.; Maiorka, A.; Bueno, I.J.M.; Schneider, A.F.; Surek, D.; Rocha, C. Vitamin E and selenium in broiler breeder diets: Effect on live performance, hatching process, and chick quality. Poult. Sci. 2015, 94, 976–983. [Google Scholar] [CrossRef]
- Li, J.; Henriksson, G.; Gellerstedt, G. Lignin depolymerization/repolymerization and its critical role for delignification of aspen wood by steam explosion. Bioresour. Technol. 2007, 98, 3061–3068. [Google Scholar] [CrossRef] [PubMed]
- Aarum, I.; Devle, H.; Ekeberg, D.; Horn, S.J.; Stenstrøm, Y. Characterization of Pseudo-Lignin from Steam Exploded Birch. ACS Omega 2018, 3, 4924–4931. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ren, X.; Wu, H.; Meng, Q.; Zhou, Z. Steam Explosion Treatment of Byproduct Feedstuffs for Potential Use as Ruminant Feed. Animals 2019, 9, 688. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Heo, J.M.; Yang, J.-K. Effects of steam-exploded wood as an insoluble dietary fiber source on the performance characteristics of broilers. BioResources 2019, 14, 1512–1524. [Google Scholar] [CrossRef]
- Svihus, B.; Choct, M.; Classen, H.L. Function and nutritional roles of the avian caeca: A review. World’s Poult. Sci. J. 2013, 69, 249–264. [Google Scholar] [CrossRef]
- Baurhoo, B.; Phillip, L.; Ruiz-Feria, C.A. Effects of Purified Lignin and Mannan Oligosaccharides on Intestinal Integrity and Microbial Populations in the Ceca and Litter of Broiler Chickens. Poult. Sci. 2007, 86, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Baurhoo, B.; Letellier, A.; Zhao, X.; Ruiz-Feria, C.A. Cecal Populations of Lactobacilli and Bifidobacteria and Escherichia coli Populations After In Vivo Escherichia coli Challenge in Birds Fed Diets with Purified Lignin or Mannanoligosaccharides. Poult. Sci. 2007, 86, 2509–2516. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kim, B.-J.; Ncho, C.-M.; Jeong, C.-M.; Gupta, V.; Jung, J.-Y.; Ha, S.-Y.; Lee, D.-H.; Yang, J.-K.; Choi, Y.-H. Dietary Supplementation of Shredded, Steam-Exploded Pine Particles Decreases Pathogenic Microbes in the Cecum of Acute Heat-Stressed Broilers. Animals 2021, 11, 2252. [Google Scholar] [CrossRef]
- Goel, A.; Kim, B.-J.; Ncho, C.-M.; Jeong, C.-M.; Gupta, V.; Jung, J.-Y.; Ha, S.-Y.; Lee, D.-H.; Yang, J.-K.; Choi, Y.-H. Utilization of Shredded Steam-Exploded Pine Particles as a Dietary Ingredient to Modify Cecal Microbiota in Broilers. Agriculture 2021, 11, 1196. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Kim, D.-H.; Lee, S.; Lee, T.; Lee, K.-W.; Chang, H.-H.; Moon, B.; Ayasan, T.; Choi, Y.-H. An Acute, Rather Than Progressive, Increase in Temperature-Humidity Index Has Severe Effects on Mortality in Laying Hens. Front. Vet. Sci. 2020, 7, 568093. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Kim, J.; Yoon, H.-S.; Choi, Y.-H. Effects of Dietary Corticosterone on Yolk Colors and Eggshell Quality in Laying Hens. Asian-Australas. J. Anim. Sci. 2015, 28, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast clustering algorithms for metagenomic sequence analysis. Brief. Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, J.; Zhong, H.; Li, N.; Xu, H.; Zhu, Q.; Liu, Y. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 2017, 7, 6400. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. [Google Scholar] [CrossRef] [PubMed]
- Mohd Shaufi, M.A.; Sieo, C.C.; Chong, C.W.; Gan, H.M.; Ho, Y.W. Deciphering chicken gut microbial dynamics based on high-throughput 16S rRNA metagenomics analyses. Gut Pathog. 2015, 7, 4. [Google Scholar] [CrossRef]
- Yuan, X.; Chen, R.; McCormick, K.L.; Zhang, Y.; Lin, X.; Yang, X. The role of the gut microbiota on the metabolic status of obese children. Microb. Cell Factories 2021, 20, 53. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Vogel, H.; Jonas, W.; Woting, A.; Blaut, M.; Schürmann, A.; Cedernaes, J. Gut microbiota and glucometabolic alterations in response to recurrent partial sleep deprivation in normal-weight young individuals. Mol. Metab. 2016, 5, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Tao, Y.; Zhang, X.; Pan, J.; Zhu, X.; Wang, H.; Du, P.; Zhu, Y.; Huang, Y.; Chen, W. Dynamic changes of blood glucose, serum biochemical parameters and gene expression in response to exogenous insulin in Arbor Acres broilers and Silky fowls. Sci. Rep. 2020, 10, 6697. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Fang, S.; Yang, H.; Gao, J.; He, M.; Ke, S.; Zhao, Y.; Chen, C.; Huang, L. Evaluating the contribution of gut microbiome to the variance of porcine serum glucose and lipid concentration. Sci. Rep. 2017, 7, 14928. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Bai, L.; Qu, Q.; Zhou, S.; Yang, M.; Guo, S.; Li, Q.; Liu, C. Impact of gut microbiota structure in heat-stressed broilers. Poult. Sci. 2019, 98, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gao, H.; Li, X.; Cao, Z.; Li, M.; Liu, J.; Qiao, Y.; Ma, L.; Zhao, Z.; Pan, H. Correlation analysis of muscle amino acid deposition and gut microbiota profile of broilers reared at different ambient temperatures. Anim. Biosci. 2021, 34, 93–101. [Google Scholar] [CrossRef]
- Shi, R.; Yang, X.; Chen, L.; Chang, H.-T.; Liu, H.-Y.; Zhao, J.; Wang, X.-W.; Wang, C.-Q. Pathogenicity of Shigella in Chickens. PLoS ONE 2014, 9, e100264. [Google Scholar] [CrossRef]
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.M.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Velge, P.; Bottreau, E.; Fievez, V.; Haesebrouck, F.; Ducatelle, R. Invasion of Salmonella enteritidis in avian intestinal epithelial cells in vitro is influenced by short-chain fatty acids. Int. J. Food Microbiol. 2003, 85, 237–248. [Google Scholar] [CrossRef]
- Faden, H. The Role of Faecalibacterium, Roseburia, and Butyrate in Inflammatory Bowel Disease. Dig. Dis. 2022, 27, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Wrzosek, L.; Miquel, S.; Noordine, M.-L.; Bouet, S.; Joncquel Chevalier-Curt, M.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitziiinfluence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Nanto-Hara, F.; Kikusato, M.; Ohwada, S.; Toyomizu, M. Heat Stress Directly Affects Intestinal Integrity in Broiler Chickens. J. Poult. Sci. 2020, 57, 284–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 185–227. [Google Scholar] [CrossRef]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Genet. 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Dolka, B.; Gołębiewska–Kosakowska, M.; Krajewski, K.; Kwieciński, P.; Nowak, T.; Szubstarski, J.; Wilczyński, J.; Szeleszczuk, P. Occurrence of Enterococcus spp. in poultry in Poland based on 2014–2015 data. Med. Weter. 2017, 73, 220–224. [Google Scholar] [CrossRef]
- Xiang, S.; Ye, K.; Li, M.; Ying, J.; Wang, H.; Han, J.; Shi, L.; Xiao, J.; Shen, Y.; Feng, X.; et al. Xylitol enhances synthesis of propionate in the colon via cross-feeding of gut microbiota. Microbiome 2021, 9, 62. [Google Scholar] [CrossRef]
- Shetty, S.A.; Zuffa, S.; Bui, T.P.N.; Aalvink, S.; Smidt, H.; De Vos, W.M. Reclassification of Eubacterium hallii as Anaerobutyricum hallii gen. nov., comb. nov., and description of Anaerobutyricum soehngenii sp. nov., a butyrate and propionate-producing bacterium from infant faeces. Int. J. Syst. Evol. Microbiol. 2018, 68, 3741–3746. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Shi, Y.; Tang, L.; Chen, L.; Fan, H.; Wang, H.; Wang, J.; Jia, X.; Chen, S.; Lai, S. Heat Stress Affects Faecal Microbial and Metabolic Alterations of Rabbits. Front. Microbiol. 2021, 12, 817615. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Guo, H.; Zheng, W.; Xue, Y.; Zhao, R.; Yao, W. Heat stress affects fecal microbial and metabolic alterations of primiparous sows during late gestation. J. Anim. Sci. Biotechnol. 2019, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Yang, Y.; Kong, Y.; Guo, Q.; Xu, Y.; Xing, P.; Sun, Y.; Qian, J.; Xu, R.; Xie, L.; et al. Gut Bacteria Erysipelatoclostridium and Its Related Metabolite Ptilosteroid A Could Predict Radiation-Induced Intestinal Injury. Front. Public Health 2022, 10, 862598. [Google Scholar] [CrossRef] [PubMed]
- Quinteiro-Filho, W.M.; Rodrigues, M.V.; Ribeiro, A.; Ferraz-De-Paula, V.; Pinheiro, M.L.; Sá, L.R.; Ferreira, A.J.; Palermo-Neto, J. Acute heat stress impairs performance parameters and induces mild intestinal enteritis in broiler chickens: Role of acute hypothalamic-pituitary-adrenal axis activation. J. Anim. Sci. 2012, 90, 1986–1994. [Google Scholar] [CrossRef] [PubMed]
- Deeb, N.; Cahaner, A. The effects of naked neck genotypes, ambient temperature, and feeding status and their interactions on body temperature and performance of broilers. Poult. Sci. 1999, 78, 1341–1346. [Google Scholar] [CrossRef]
- Nascimento, S.T.; Silva, I.J.O.d.; Mourão, G.B.; Castro, A.C.d. Bands of respiratory rate and cloacal temperature for different broiler chicken strains. Rev. Bras. Zootec. 2012, 41, 1318–1324. [Google Scholar] [CrossRef]
- El Hadi, H.; Sykes, A.H. Thermal panting and respiratory alkalosis in the laying hen. Br. Poult. Sci. 1982, 23, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Emami, N.K.; Jung, U.; Voy, B.; Dridi, S. Radical Response: Effects of Heat Stress-Induced Oxidative Stress on Lipid Metabolism in the Avian Liver. Antioxidants 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Dietary taurine supplementation decreases fat synthesis by suppressing the liver X receptor α pathway and alleviates lipid accumulation in the liver of chronic heat-stressed broilers. J. Sci. Food Agric. 2019, 99, 5631–5637. [Google Scholar] [CrossRef]
- Shim, K.S.; Hwang, K.T.; Son, M.W.; Park, G.H. Lipid Metabolism and Peroxidation in Broiler Chicks under Chronic Heat Stress. Asian-Australasian J. Anim. Sci. 2006, 19, 1206–1211. [Google Scholar] [CrossRef]
- Hirakawa, R.; Nurjanah, S.; Furukawa, K.; Murai, A.; Kikusato, M.; Nochi, T.; Toyomizu, M. Heat Stress Causes Immune Abnormalities via Massive Damage to Effect Proliferation and Differentiation of Lymphocytes in Broiler Chickens. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Xie, J.; Tang, L.; Lu, L.; Zhang, L.; Lin, X.; Liu, H.-C.; Odle, J.; Luo, X. Effects of acute and chronic heat stress on plasma metabolites, hormones and oxidant status in restrictedly fed broiler breeders. Poult. Sci. 2015, 94, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Garriga, C.; Hunter, R.R.; Amat, C.; Planas, J.M.; Mitchell, M.A.; Moretó, M. Heat stress increases apical glucose transport in the chicken jejunum. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R195–R201. [Google Scholar] [CrossRef]
- He, J.; He, Y.; Pan, D.; Cao, J.; Sun, Y.; Zeng, X. Associations of Gut Microbiota With Heat Stress-Induced Changes of Growth, Fat Deposition, Intestinal Morphology, and Antioxidant Capacity in Ducks. Front. Microbiol. 2019, 10, 903. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Final BW (g) | % Difference in BW | Feed Intake (g/Bird) | ||
|---|---|---|---|---|
| Treatments | 0% NT | 2535 b ± 13 | 1.79 b ± 0.53 | 62 ± 5.6 |
| 0% HS | 2454 a ± 13 | −1.46 a ± 0.53 | 44 ± 5.6 | |
| 0.1% HS | 2466 a ± 13 | −0.95 a ± 0.53 | 46 ± 5.6 | |
| 0.4% HS | 2476 a ± 13 | −0.56 a ± 0.53 | 47 ± 5.6 | |
| p value | 0.003 | 0.003 | 0.128 | |
| Initial BW | Est | 0.956 | −0.002 | 0.025 |
| SE | 0.109 | 0.004 | 0.045 | |
| p value | 0.001 | 0.679 | 0.586 | |
| Contrast p values | 0% NT vs. 0% HS | 0.068 | 0.001 | 0.028 |
| 0% NT vs. 0.1% HS | 0.305 | 0.002 | 0.070 | |
| 0% NT vs. 0.4% HS | 0.181 | 0.005 | 0.066 | |
| 0% HS vs. 0.1% HS | 0.385 | 0.541 | 0.647 | |
| 0% HS vs. 0.4% HS | 0.586 | 0.238 | 0.665 | |
| 0% HS vs. 0.1% HS, and 0.4% HS | 0.415 | 0.301 | 0.607 | |
| 0% NT vs. 0% HS, 0.1% HS, and 0.4% HS | 0.091 | 0.001 | 0.020 |
| Treatments | Rectal Temperature (°C) | ||
|---|---|---|---|
| Before | After | ΔT | |
| 0% NT | 40.9 ± 0.15 | 41.3 a ± 0.10 | 0.4 a ± 0.15 |
| 0% HS | 41.2 ± 0.18 | 43.6 bc ± 0.23 | 2.4 bc ± 0.29 |
| 0.1% HS | 41.1 ± 0.13 | 44.0 c ± 0.45 | 2.9 c ± 0.36 |
| 0.4% HS | 41.1 ± 0.10 | 42.9 b ± 0.13 | 1.7 b ± 0.17 |
| p value | 0.565 | 0.001 | 0.001 |
| Contrast p values | |||
| 0% NT vs. 0% HS | 0.178 | 0.001 | 0.001 |
| 0% NT vs. 0.1% HS | 0.433 | 0.001 | 0.001 |
| 0% NT vs. 0.4% HS | 0.330 | 0.001 | 0.002 |
| 0% HS vs. 0.1% HS | 0.555 | 0.302 | 0.173 |
| 0% HS vs. 0.4% HS | 0.693 | 0.066 | 0.089 |
| 0% HS vs. 0.1% HS and 0.4% HS | 0.570 | 0.608 | 0.827 |
| 0% NT vs. 0% HS, 0.1% HS, and 0.4% HS | 0.208 | 0.001 | 0.001 |
| Relative Organ Weight (%) | |||
|---|---|---|---|
| Treatment | Liver | Spleen | Bursa |
| 0% NT | 2.83 ± 0.06 | 0.12 ± 0.01 | 0.10 ± 0.02 |
| 0% HS | 2.60 ± 0.17 | 0.14 ± 0.01 | 0.09 ± 0.01 |
| 0.1% HS | 2.60 ± 0.16 | 0.15 ± 0.01 | 0.10 ± 0.01 |
| 0.4% HS | 2.51 ± 0.19 | 0.13 ± 0.01 | 0.12 ± 0.01 |
| p value | 0.493 | 0.436 | 0.522 |
| Contrast p values | |||
| 0% NT vs. 0% HS | 0.291 | 0.247 | 0.595 |
| 0% NT vs. 0.1% HS | 0.294 | 0.139 | 0.905 |
| 0% NT vs. 0.4% HS | 0.147 | 0.610 | 0.346 |
| 0% HS vs. 0.1% HS | 0.995 | 0.732 | 0.517 |
| 0% HS vs. 0.4% HS | 0.676 | 0.507 | 0.148 |
| 0% HS vs. 0.1% HS and 0.4% HS | 0.812 | 0.852 | 0.226 |
| 0% NT vs. 0% HS, 0.1% HS, and 0.4% HS | 0.149 | 0.199 | 0.826 |
| Blood Biochemicals (mg/dl) | ||||
|---|---|---|---|---|
| Treatment | Glucose | Total Protein | Triglyceride | Cholesterol |
| 0% NT | 295.5 ± 24.9 | 3.3 ± 0.18 | 47.5 ± 11.5 | 139.3 ± 8.5 |
| 0% HS | 326 ± 16.4 | 3.5 ± 0.17 | 24 ± 3.6 | 135.5 ± 8.0 |
| 0.1% HS | 370.3 ± 45.8 | 3.8 ± 0.27 | 31 ± 6.2 | 129.3 ± 7.9 |
| 0.4% HS | 305.3 ± 29.7 | 3.9 ± 0.41 | 25.2 ± 5.4 | 152 ± 22.4 |
| p value | 0.357 | 0.408 | 0.121 | 0.672 |
| Contrast p values | ||||
| 0% NT vs. 0% HS | 0.496 | 0.704 | 0.034 | 0.840 |
| 0% NT vs. 0.1% HS | 0.104 | 0.279 | 0.127 | 0.599 |
| 0% NT vs. 0.4% HS | 0.825 | 0.129 | 0.043 | 0.507 |
| 0% HS vs. 0.1% HS | 0.325 | 0.476 | 0.507 | 0.745 |
| 0% HS vs. 0.4% HS | 0.643 | 0.245 | 0.911 | 0.389 |
| 0% HS vs. 0.1% HS and 0.4% HS | 0.759 | 0.280 | 0.654 | 0.753 |
| 0% NT vs. 0% HS, 0.1% HS, and 0.4% HS | 0.298 | 0.223 | 0.023 | 0.980 |
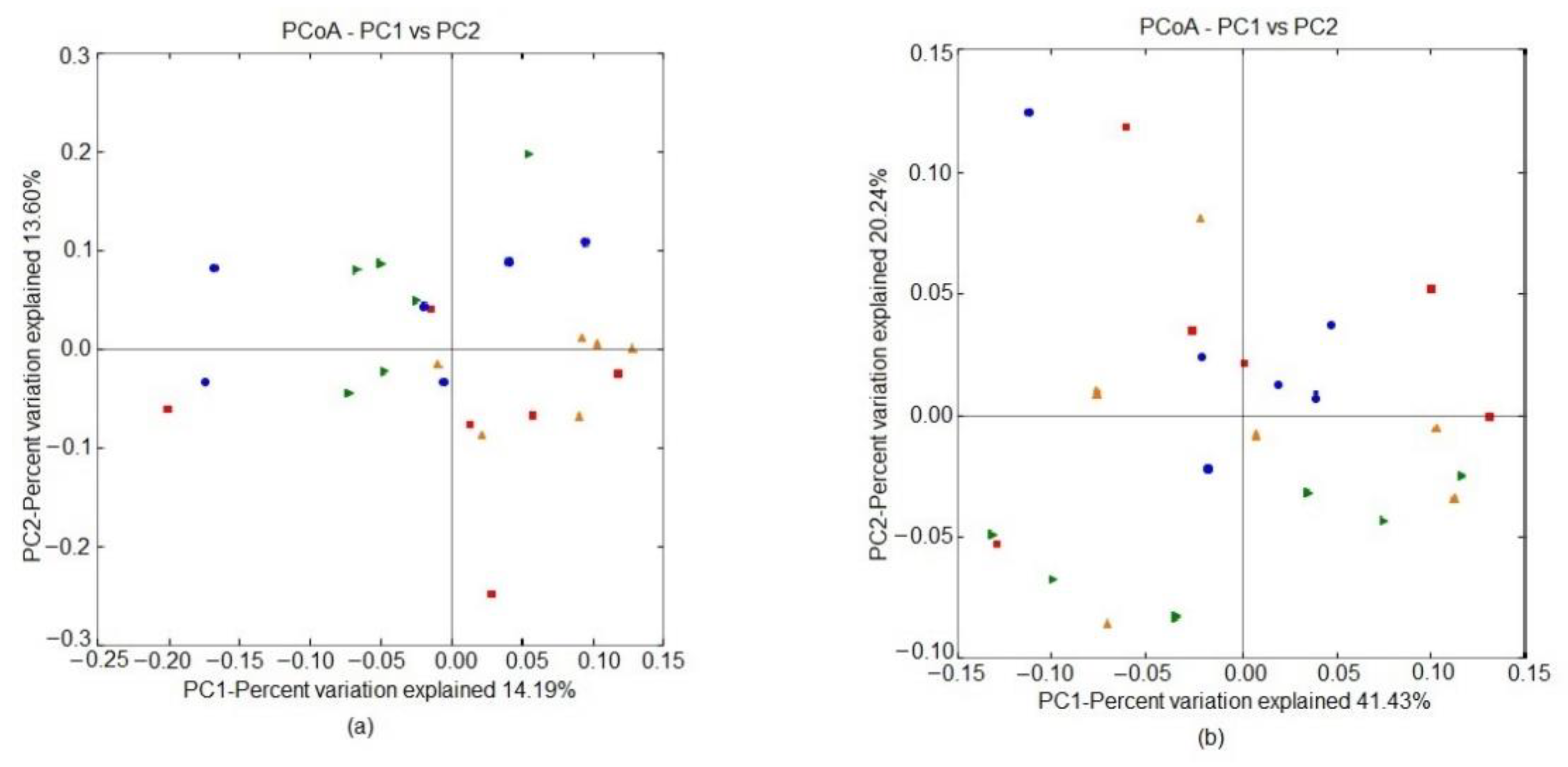
| PERMANOVA | ||
|---|---|---|
| Unweighted Unifrac Distances | Weighted Unifrac Distances | |
| Test statistic | 1.352 | 0.926 |
| p value | 0.02 | 0.469 |
| Number of permutations | 999 | 999 |
| Pairwise comparison | ||
| 0% NT vs. 0% HS | 0.183 | 0.983 |
| 0% NT vs. 0.1% HS | 0.007 | 0.397 |
| 0% NT vs. 0.4% HS | 0.610 | 0.122 |
| 0% HS vs. 0.1% HS | 0.295 | 0.576 |
| 0% HS vs. 0.4% HS | 0.063 | 0.273 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goel, A.; Ncho, C.-M.; Jeong, C.-M.; Gupta, V.; Jung, J.-Y.; Ha, S.-Y.; Yang, J.-K.; Choi, Y.-H. Effects of Dietary Supplementation of Solubles from Shredded, Steam-Exploded Pine Particles on the Performance and Cecum Microbiota of Acute Heat-Stressed Broilers. Microorganisms 2022, 10, 1795. https://doi.org/10.3390/microorganisms10091795
Goel A, Ncho C-M, Jeong C-M, Gupta V, Jung J-Y, Ha S-Y, Yang J-K, Choi Y-H. Effects of Dietary Supplementation of Solubles from Shredded, Steam-Exploded Pine Particles on the Performance and Cecum Microbiota of Acute Heat-Stressed Broilers. Microorganisms. 2022; 10(9):1795. https://doi.org/10.3390/microorganisms10091795
Chicago/Turabian StyleGoel, Akshat, Chris-Major Ncho, Chae-Mi Jeong, Vaishali Gupta, Ji-Young Jung, Si-Young Ha, Jae-Kyung Yang, and Yang-Ho Choi. 2022. "Effects of Dietary Supplementation of Solubles from Shredded, Steam-Exploded Pine Particles on the Performance and Cecum Microbiota of Acute Heat-Stressed Broilers" Microorganisms 10, no. 9: 1795. https://doi.org/10.3390/microorganisms10091795