
Isolation of Novel Bacterial Strains Pseudomonas extremaustralis CSW01 and Stutzerimonas stutzeri CSW02 from Sewage Sludge for Paracetamol Biodegradation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Enrichment Cultures and Bacterial Identification
2.3. Inoculum Preparation
2.4. Inhibitory Concentration of APAP for Bacterial Growth
2.5. Biodegradation Assays
2.6. Models of Biodegradation Kinetics
2.7. Detection of APAP and Its Main Metabolites
3. Results and Discussion
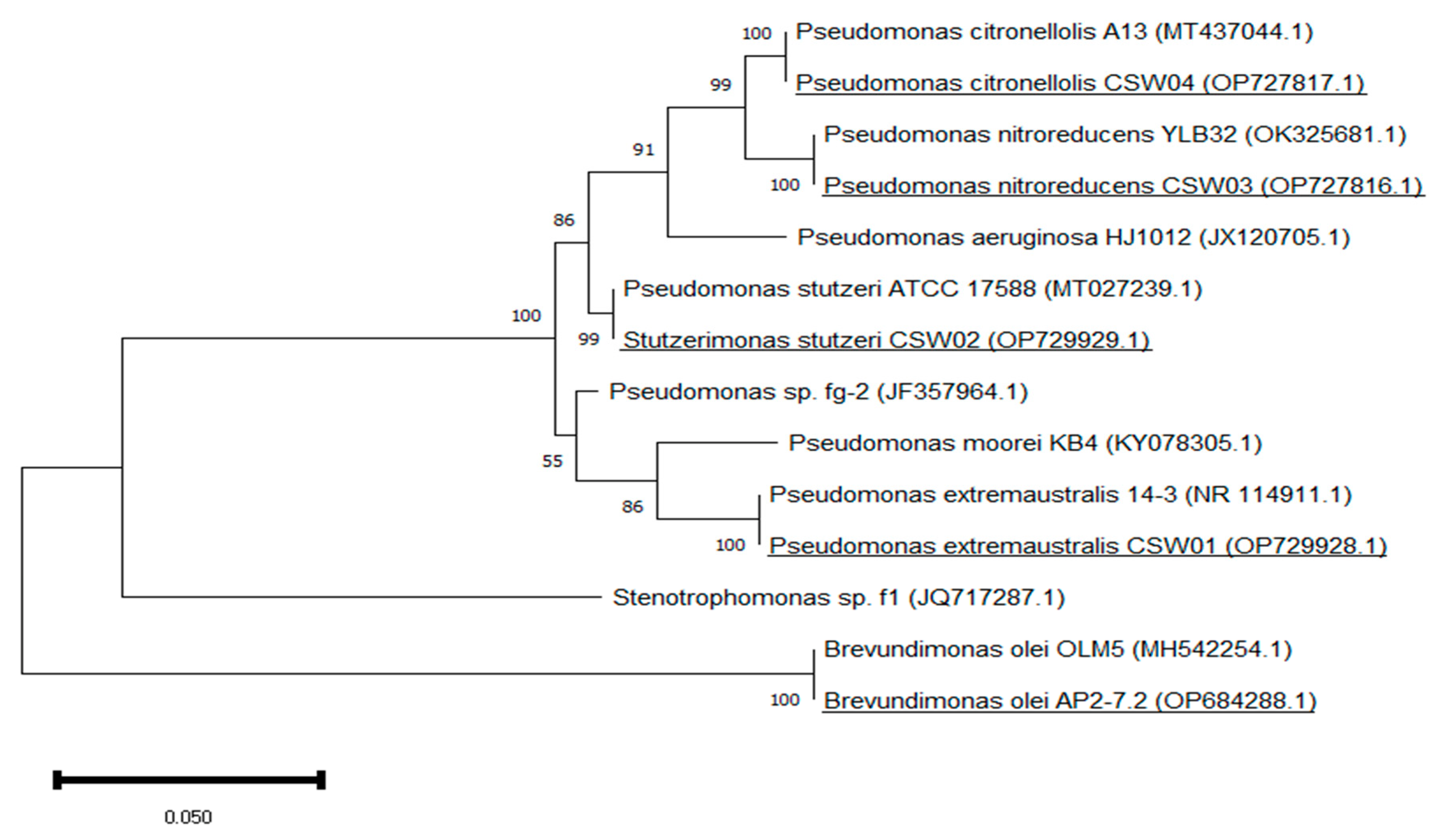
3.1. Enrichment Cultures for the Isolation and Identification of APAP-Degrading Bacterial Strains
3.2. APAP Degradation in Aqueous Solution
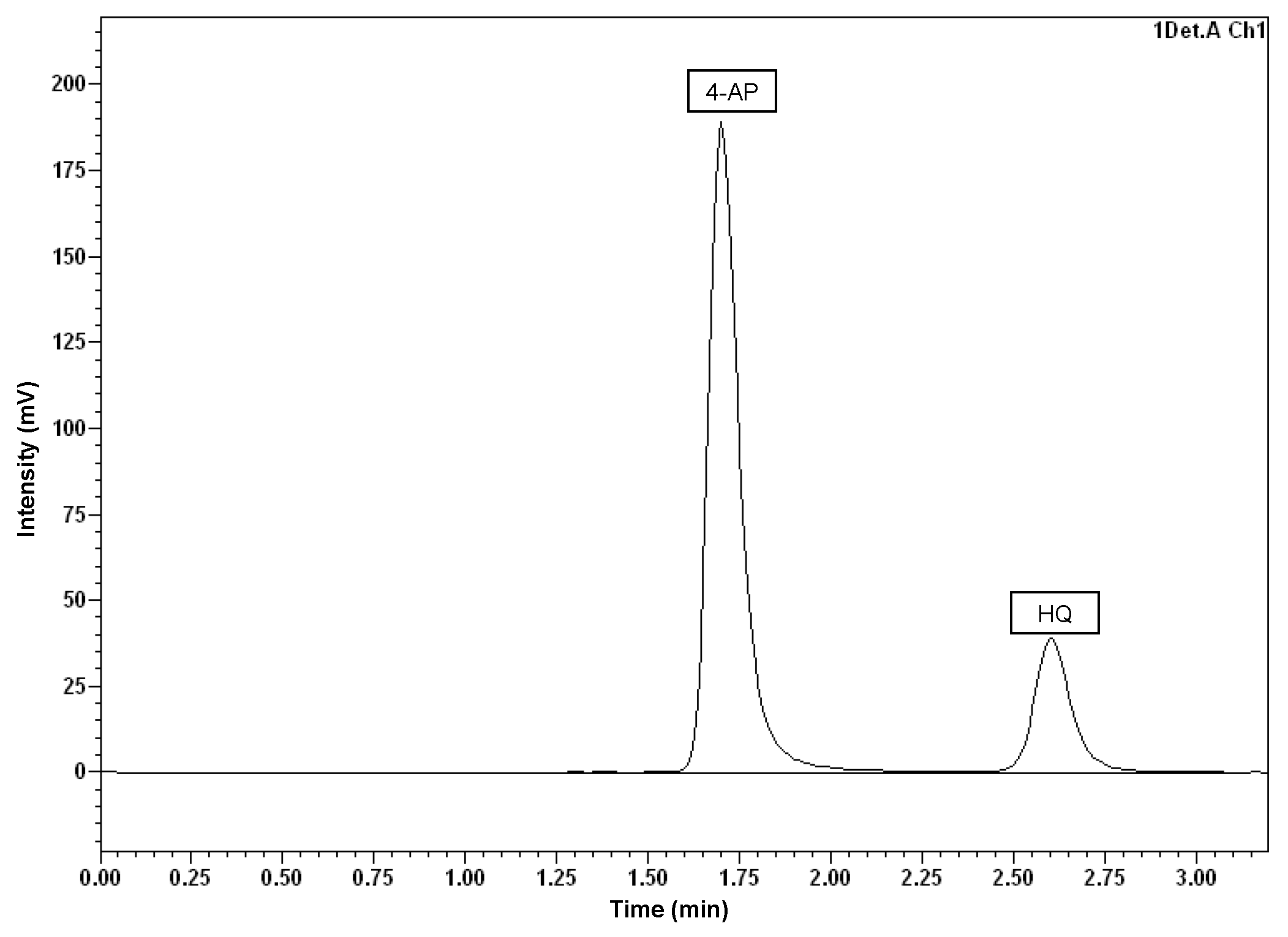
3.3. Detection and Quantification of Main Metabolites
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chaturvedi, P.; Shukla, P.; Giri, B.S.; Chowdhary, P.; Chandra, R.; Gupta, P.; Pandey, A. Prevalence and hazardous impact of pharmaceutical and personal care products and antibiotics in environment: A review on emerging contaminants. Environ. Res. 2021, 194, 110664. [Google Scholar] [CrossRef] [PubMed]
- Mejías, C.; Martín, J.; Santos, J.L.; Aparicio, I.; Alonso, E. Occurrence of pharmaceuticals and their metabolites in sewage sludge and soil: A review on their distribution and environmental risk assessment. Trends Environ. Anal. Chem. 2021, 30, e00125. [Google Scholar] [CrossRef]
- Lara-Moreno, A.; Aguilar-Romero, I.; Rubio-Bellido, M.; Madrid, F.; Villaverde, J.; Santos, J.L.; Alonso, E.; Morillo, E. Novel nonylphenol-degrading bacterial strains isolated from sewage sludge: Application in bioremediation of sludge. Sci. Total Environ. 2022, 847, 157647. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, M.; Bijlsma, L.; Pitarch, E.; López, F.J.; Hernández, F. Occurrence of pharmaceutical metabolites and transformation products in the aquatic environment of the Mediterranean area. Trends Environ. Anal. Chem. 2021, 29, 00118. [Google Scholar] [CrossRef]
- Bolesta, W.; Głodniok, M.; Styszko, K. From Sewage Sludge to the Soil—Transfer of Pharmaceuticals: A Review. Int. J. Environ. Res. Public Health 2022, 19, 10246. [Google Scholar] [CrossRef]
- Zhang, C.; Leon Barron, L.; Sturzenbaum, S. The transportation, transformation and (bio)accumulation of pharmaceuticals in the terrestrial ecosystem. Sci. Total Environ. 2021, 781, 146684. [Google Scholar] [CrossRef]
- Ziylan-Yavas, A.; Santos, D.; Moraes Flores, E.M.; Ince, N.H. Pharmaceuticals and personal care products (PPCPs): Environmental and public health risks. Environ. Prog. Sustain. Energy 2022, 41, e13821. [Google Scholar] [CrossRef]
- Hider-Mlynarz, K.; Cavalie, P.; Maison, P. Trends in analgesic consumption in France over the last 10 years and comparison of patterns across Europe. Br. J. Clin. Pharmacol. 2018, 84, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Żur, J.; Piński, A.; Marchlewicz, A.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic micropollutants paracetamol and ibuprofen—Toxicity, biodegradation, and genetic background of their utilization by bacteria. Environ. Sci. Pollut. Res. 2018, 25, 21498–21524. [Google Scholar] [CrossRef] [Green Version]
- Ebele, A.J.; Abdallah, M.A.E.; Harrad, S. Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerg. Contam. 2017, 3, 1–16. [Google Scholar] [CrossRef]
- Du, J.; Mei, C.F.; Ying, G.G.; Xu, M.Y. Toxicity thresholds for diclofenac, acetaminophen and ibuprofen in the water flea Daphnia magna. Bull. Environ. Contam. Toxicol. 2016, 97, 84–90. [Google Scholar] [CrossRef]
- Guiloski, I.C.; Ribas, J.L.C.; Piancini, L.D.S.; Dagostim, A.C.; Cirio, S.M.; Favaro, L.F.; Boschen, S.L.; Cestari, M.M.; da Cunha, C.; de Assis, H.C. Paracetamol causes endocrine disruption and hepatotoxicity in male fish Rhamdia quelen after subchronic exposure. Environ. Toxicol. Pharmacol. 2017, 53, 111–120. [Google Scholar] [CrossRef]
- Guerra, M.H.; Alberola, I.O.; Rodriguez, S.M.; Lopez, A.A.; Merino, A.A.; Lopera, A.E.C.; Alonso, J.Q. Oxidation mechanisms of amoxicillin and paracetamol in the photo-Fenton solar process. Water Res. 2019, 156, 232–240. [Google Scholar] [CrossRef]
- Bouissou-Schurtz, C.; Houeto, P.; Guerbet, M.; Bachelot, M.; Casellas, C.; Mauclaire, A.C.; Panetier, P.; Delval, C.; Masset, D. Ecological risk assessment of the presence of pharmaceutical residues in a French national water survey. Regul. Toxicol. Pharmacol. 2014, 69, 296–303. [Google Scholar] [CrossRef]
- Fekadu, S.; Alemayehu, E.; Dewil, R.; Van der Bruggen, B. Pharmaceuticals in freshwater aquatic environments: A comparison of the African and European challenge. Sci. Total Environ. 2019, 654, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Torun, M.; Gültekin, O.; Solpan, D.; Güven, O. Mineralization of paracetamol in aqueous solution with advanced oxidation processes. Environ. Technol. 2015, 36, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Villota, N.; Lomas, J.M.; Camarero, L.M. Study of the paracetamol degradation pathway that generates color and turbidity in oxidized wastewaters by photo-Fenton technology. J. Photochem. Photobiol. A Chem. 2016, 329, 113–119. [Google Scholar] [CrossRef]
- Gusseme, B.D.; Vanhaecke, L.; Verstraete, W.; Boon, N. Degradation of acetaminophen by Deftia tsuruhatensis and Pseudomonas aeruginosa in a membrane bioreactor. Water. Res. 2011, 45, 1829–1837. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, L.L.; Chen, J.M.; Liu, Y. Degradation of paracetamol by Pseudomonas aeruginosa strain HJ1012. J. Environ. Sci. Health A 2013, 48, 791–799. [Google Scholar] [CrossRef]
- Zhang, L.L.; Hu, J.; Zhu, R.Y.; Zhou, Q.W.; Chen, J.M. Degradation of paracetamol by pure bacterial cultures and their microbial consortium. Appl. Microbiol. Biotechnol. 2013, 97, 3687–3698. [Google Scholar] [CrossRef]
- Żur, J.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Marchlewicz, A.; Guzik, U. Paracetamol—Toxicity and microbial utilization. Pseudomonas moorei KB4 as a case study for exploring degradation pathway. Chemosphere 2018, 206, 192–202. [Google Scholar] [CrossRef]
- Palma, T.L.; Donaldben, M.N.; Costa, M.C.; Carlier, J.D. Putative role of flavobacterium, dokdonella and methylophilus strains in paracetamol biodegradation. Water Air Soil Pollut. 2018, 229, 200. [Google Scholar] [CrossRef]
- Wei, F.; Zhou, Q.W.; Leng, S.Q. Isolation, identification and biodegradation characteristics of a new bacterial strain degrading paracetamol. Chin. J. Environ. Sci. 2011, 32, 1812–1819. [Google Scholar]
- Park, S.; Oh, S. Detoxification and bioaugmentation potential for acetaminophen and its derivatives using Ensifer sp. isolated from activated sludge. Chemosphere 2020, 260, 127532. [Google Scholar] [CrossRef]
- Chen, R.; Liu, X.; Ma, Y. Isolation and identification of acetaminophen degrading strain Shinella sp. HZA2. J. Environ. Sci. Health Part B 2022, 57, 333–338. [Google Scholar] [CrossRef]
- Chopra, S.; Kumar, D. Biodegradation and Kinetic Analysis of Acetaminophen with Co-culture of Bacterial Strains Isolated from Sewage Wastewater. Curr. Microbiol. 2020, 77, 3147–3157. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.; Agarwala, R.; Schaffer, A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 2942. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.; Rubio-Bellido, M.; Merchán, F.; Morillo, E. Bioremediation of diuron contaminated soils by a novel degrading microbial consortium. J. Environ. Manag. 2017, 188, 379–386. [Google Scholar] [CrossRef] [PubMed]
- FOCUS. Guidance Document on Estimating Persistence and Degradation Kinetics from Environmental Fate Studies on Pesticides in EU Registration. Report of the FOCUS Work Group on Degradation Kinetics, EC Document Reference Sanco/10058/2005 Version 2.0. 2006, p. 434. Available online: https://esdac.jrc.ec.europa.eu/public_path/projects_data/focus/dk/docs/finalreportFOCDegKinetics.pdf (accessed on 10 December 2022).
- Gu, J.D. On enrichment culturing and transferring technique. Appl. Environ. Biotechnol. 2021, 6, 1–5. [Google Scholar] [CrossRef]
- Lalucat, J.; Gomila, M.; Mulet, M.; Zaruma, A.; García-Valdés, E. Past, present and future of the boundaries of the pseudomonas genus: Proposal of stutzerimonas gen. Nov. Syst. Appl. Microbiol. 2022, 45, 126289. [Google Scholar] [CrossRef]
- Gomila, M.; Mulet, M.; García-Valdés, E.; Lalucat, J. Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans. Microorganisms 2022, 10, 1363. [Google Scholar] [CrossRef]
- World Health Organization. Laboratory Biosafety Manual, 3rd ed.; World Health Organization: Geneva, Switzerland, 2004. Available online: https://apps.who.int/iris/handle/10665/42981 (accessed on 10 December 2022).
- Kaczorek, E.; Sałek, K.; Guzik, U.; Jesionowski, T.; Cybulski, Z. Biodegradation of alkyl derivatives of aromatic hydrocarbons and cell surface properties of a strain of pseudomonas stutzeri. Chemosphere 2013, 90, 471–478. [Google Scholar] [CrossRef]
- Parthipan, P.; Elumalai, P.; Sathishkumar, K.; Sabarinathan, D.; Murugan, K.; Benelli, G.; Rajasekar, A. Biosurfactant and enzyme mediated crude oil degradation by Pseudomonas stutzeri NA3 and Acinetobacter baumannii MN3. 3 Biotech 2017, 7, 278. [Google Scholar] [CrossRef]
- Bera, S.P.; Tank, S.K. Microbial degradation of Procion Red by pseudomonas stutzeri. Sci. Rep. 2021, 11, 3075. [Google Scholar] [CrossRef]
- Shahriarinour, M.; Divsar, F.; Dahka, F.K.; Chegini, S.N.; Mahani, M.; Moeini, A.; Cerruti, P. Biodegradation of Alprazolam in Pharmaceutical Wastewater Using Mesoporous Nanoparticles-Adhered Pseudomonas stutzeri. Molecules 2022, 27, 237. [Google Scholar] [CrossRef]
- López, N.I.; Pettinari, M.J.; Stackebrandt, E.; Tribelli, P.M.; Põtter, M.; Steinbüchel, A.; Méndez, B.S. Pseudomonas extremaustralis sp. nov., a poly(3-hydroxybutyrate) producer isolated from an antarctic environment. Curr. Microbiol. 2009, 59, 514–519. [Google Scholar] [CrossRef]
- Colonnella, M.A.; Lizarraga, L.; Rossi, L.; Díaz Peña, R.; Egoburo, D.; López, N.I.; Iustman, L.J. Effect of copper on diesel degradation in Pseudomonas extremaustralis. Extremophiles 2018, 23, 91–99. [Google Scholar] [CrossRef]
- López, G.; Diaz-Cárdenas, C.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; David Alzate, J.; González, L.N.; Restrepo, S.; Baena, S. Draft genome sequence of Pseudomonas extremaustralis strain USBA-GBX 515 isolated from Superparamo soil samples in Colombian Andes. Stand. Genom. Sci. 2017, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Finore, I.; Vigneron, A.; Vincent, W.F.; Leone, L.; Di Donato, P.; Schiano Moriello, A.; Nicolaus, B.; Poli, A. Novel Psychrophiles and Exopolymers from Permafrost Thaw Lake Sediments. Microorganisms 2020, 8, 1282. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, X.; Zhang, S.; Song, H.; Kong, Z. Insights into the bacterial species and communities of a full-scale anaerobic/anoxic/oxic wastewater treatment plant by using third-generation sequencing. J. Biosci. Bioeng. 2019, 128, 744–750. [Google Scholar] [CrossRef]
- Ahmed, S.; Javed, M.A.; Tanvir, S.; Hameed, A. Isolation and characterization of a pseudomonas strain that degrades 4-acetamidophenol and 4-aminophenol. Biodegradation 2001, 12, 303–309. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | IUPAC Name | Structure | Molecular Weight (g mol−1) | Water Solubility (20 °C) | pKa | Log KOW |
|---|---|---|---|---|---|---|
| Acetaminophen; paracetamol | N-(4-hydroxyphenyl)acetamide |  | 151.16 | 14 g L−1 | 9.38 | 0.46 |
| 4-aminophenol | 4-aminophenol |  | 109.13 | 16 g L−1 | pK1 = 5.48 pK2 = 10.46 | 0.04 |
| Hydroquinone | Benzene-1,4-diol |  | 110.11 | 6.7 g L−1 | 9.96 | 0.59 |
| Organic matter (%) | 48.0 | (7.2) |
| pH | 8.25 | (0.03) |
| Nitrogen (Kjeldahl) (%) | 5.05 | (0.76) |
| P2O5 (%) | 6.00 | (1.20) |
| K2O (%) | 0.42 | (0.07) |
| CaO (%) | 7.11 | (1.42) |
| MgO (%) | 0.94 | (0.17) |
| Fe (mg kg−1) | >50,000 | (8000) |
| Cd (mg kg−1) | <2 | (0.3) |
| Cu (mg kg−1) | 203 | (30) |
| Ni (mg kg−1) | 17 | (2) |
| Pb (mg kg−1) | 37 | (7) |
| Zn (mg kg−1) | 620 | (105) |
| Hg (mg kg−1) | 0.40 | (0.10) |
| Cr (mg kg−1) | 32 | (6) |
| Strain (Accession Number) | NCBI Affiliation (Accession Number) | Similarity | Class; Order; Family; Genus |
|---|---|---|---|
| CSW1(OP729928.1) | Pseudomonas extremaustralis 14-3 (NR_114911.1) | 100% | Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas |
| CSW2 (OP729929.1) | Pseudomonas stutzeri ATCC 17588 (MT027239.1) | 100% | Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Stutzerimonas |
| CSW3 (OP727816.1) | Pseudomonas nitroreducens YLB32 (OK325681.1) | 100% | Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas |
| CSW4 (OP727817.1) | Pseudomonas citronellolis A13 (MT437044.1) | 100% | Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas |
| AP2-7.2 (OP684288.1) | Brevundimonas olei OLM5 (MH542254.1) | 100% | Alphaproteobacteria; Caulobacterales; Caulobacteraceae; Brevundimonas |
| Bacterial Strain | APAP Concentration (mg L−1) | Model Kinetic | K (Day−1) | α (Day−1) | β (Day−1) | DT50 (h) | DT90 (h) | χ2 * | Scaled Error | R2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Pseudomonas extremaustralis CSW01 | 200 | SFO | 0.684 | - | - | 1.01 | 3.37 | 12.59 | 16.3 | 0.933 |
| 500 | SFO | 0.448 | - | - | 1.55 | 5.14 | 15.51 | 27.3 | 0.962 | |
| 2000 | FOMC | - | 151,924 | 1,318,434 | 6.02 | 20.0 | 22.36 | 168 | 0.878 | |
| 3000 | FOMC | - | 36,266 | 1,326,513 | 25.3 | 84.2 | 18.31 | 127 | 0.964 | |
| Stutzerimonas stutzeri CSW02 | 200 | SFO | 1.022 | - | - | 0.68 | 2.25 | 12.59 | 14.1 | 0.952 |
| 500 | SFO | 0.995 | - | - | 0.70 | 2.31 | 12.59 | 3.79 | 0.999 | |
| 2000 | FOMC | - | 218,321 | 1,313,894 | 4.17 | 13.9 | 18.31 | 160 | 0.912 | |
| 3000 | FOMC | - | 15,129 | 1,327,039 | 60.8 | 202 | 18.31 | 176 | 0.748 |
| Strain | Initial Concentration (mg L−1) | Complete Degradation (h) | Reference |
|---|---|---|---|
| Stutzerimonas stutzeri CSW02 | 500 | 4 | This study |
| Pseudomonas extremaustralis CSW01 | 500 | 6 | This study |
| Pseudomonas sp. ST1 | 777 | 72 | [49] |
| Cupriavidus necator F1 | 400 | 48 | [23] |
| Pseudomonas aeruginosa HJ1012 | 500 | 12 | [19] |
| Stenotrophomonas sp. f1 | 400 | 116 | [20] |
| Pseudomonas sp. fg-2 | 600 | 23 | [20] |
| Pseudomonas sp. f2 | 600 | 38 | [20] |
| Ensifer sp. POKHU | 400 | 65 | [24] |
| Shinella sp. HZA2 | 600 | 30 | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Ordóñez, A.; Aguilar-Romero, I.; Villaverde, J.; Madrid, F.; Morillo, E. Isolation of Novel Bacterial Strains Pseudomonas extremaustralis CSW01 and Stutzerimonas stutzeri CSW02 from Sewage Sludge for Paracetamol Biodegradation. Microorganisms 2023, 11, 196. https://doi.org/10.3390/microorganisms11010196
Vargas-Ordóñez A, Aguilar-Romero I, Villaverde J, Madrid F, Morillo E. Isolation of Novel Bacterial Strains Pseudomonas extremaustralis CSW01 and Stutzerimonas stutzeri CSW02 from Sewage Sludge for Paracetamol Biodegradation. Microorganisms. 2023; 11(1):196. https://doi.org/10.3390/microorganisms11010196
Chicago/Turabian StyleVargas-Ordóñez, Antonio, Inés Aguilar-Romero, Jaime Villaverde, Fernando Madrid, and Esmeralda Morillo. 2023. "Isolation of Novel Bacterial Strains Pseudomonas extremaustralis CSW01 and Stutzerimonas stutzeri CSW02 from Sewage Sludge for Paracetamol Biodegradation" Microorganisms 11, no. 1: 196. https://doi.org/10.3390/microorganisms11010196