
Cell-Free Culture Supernatant of Lactobacillus acidophilus AG01 and Bifidobacterium animalis subsp. lactis AG02 Reduces the Pathogenicity of NetB-Positive Clostridium perfringens in a Chicken Intestinal Epithelial Cell Line
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Cell Line, Bacterial Strains and Culture Conditions
2.3. Preparation of Cell-Free Supernatant (CFS)
2.4. Cell Adhesion Assay
2.5. Cell Permeability Assay
2.6. Cytotoxicity Assay
2.7. CHIC-8E11 Protein Expression without or with Probiotic CFS Pre-Treatment
2.8. Statistical Analysis
3. Results
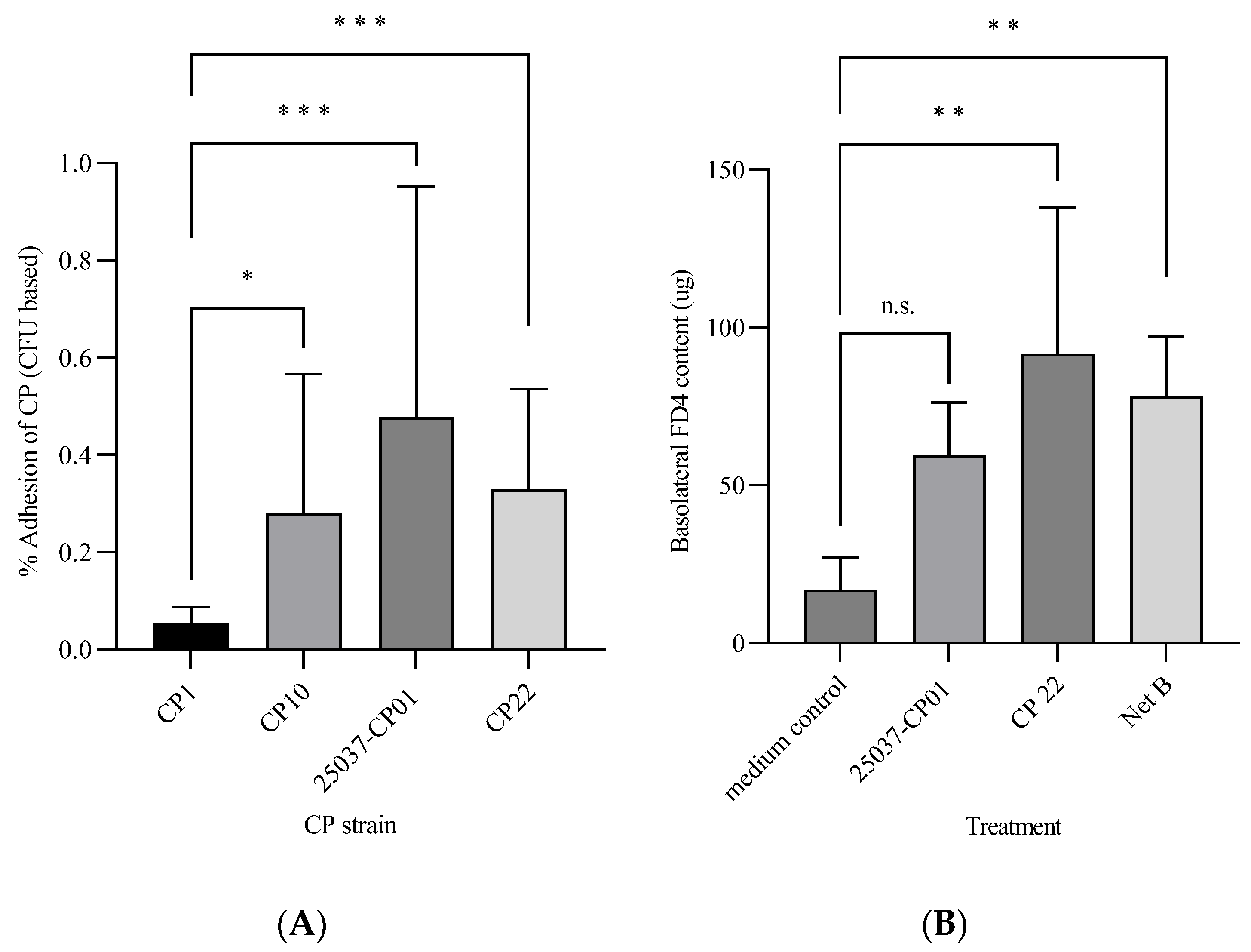
3.1. Pathogenic Effects of C. perfringens against CHIC-8E11 Cells
3.2. Effect of Pre-Treatment with Probiotics on C. perfringens Pathogenic Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| CFU | Colony-forming unit |
| CP | Clostridium perfringens |
| CFS | Cell-free supernatant |
| d | Day(s) |
| FBS | Fetal bovine serum |
| FD4 | FITC-dextran (4 KD) |
| LCA | Latent class model |
| MRS | Man–Rogosa–Sharpe broth |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OD | Optical density |
| PBS | Phosphate-buffered saline |
| TNF | Tumor necrosis factor |
| TSB | Tryptic soy broth |
References
- Kogut, M.H. Role of diet-microbiota interactions in precision nutrition of the chicken: Facts, gaps, and new concepts. Poult. Sci. 2022, 101, 101673. [Google Scholar] [CrossRef] [PubMed]
- Kairmi, S.H.; Taha-Abdelaziz, K.; Yitbarek, A.; Sargolzaei, M.; Spahany, H.; Astill, J.; Shojadoost, B.; Alizadeh, M.; Kulkarni, R.R.; Parkinson, J.; et al. Effects of therapeutic levels of dietary antibiotics on the cecal microbiome composition of broiler chickens. Poult. Sci. 2022, 101, 101864. [Google Scholar] [CrossRef] [PubMed]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. Poult. World 2015, 31, 16–17. [Google Scholar]
- Revitt-Mills, S.A.; Rood, J.I.; Adams, V. Clostridium perfringens extracellular toxins and enzymes: 20 and counting. Microbiol. Aust. 2015, 36, 114–117. [Google Scholar] [CrossRef]
- Emami, N.K.; Dalloul, R.A. Centennial Review: Recent developments in host-pathogen interactions during necrotic enteritis in poultry. Poult. Sci. 2021, 100, 101330. [Google Scholar] [CrossRef] [PubMed]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef] [PubMed]
- Bendary, M.M.; Abd El-Hamid, M.I.; El-Tarabili, R.M.; Hefny, A.A.; Algendy, R.M.; Elzohairy, N.A.; Ghoneim, M.M.; Al-Sanea, M.M.; Nahari, M.H.; Moustafa, W.H. Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology 2022, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef]
- Eid, S.; Atfehy, N.; Amer, F.; Tolba, H.; Hamed, R. Prevention of Necrotic Enteritis in Broiler Chickens by Prebiotics and Probiotics VS Control by Antibiotics, in Vivo Study. Alex. J. Vet. Sci. 2020, 64, 143–153. [Google Scholar] [CrossRef]
- Abd El-Ghany, W.A.; Abdel-Latif, M.A.; Hosny, F.; Alatfeehy, N.M.; Noreldin, A.E.; Quesnell, R.R.; Chapman, R.; Sakai, L.; Elbestawy, A.R. Comparative efficacy of postbiotic, probiotic, and antibiotic against necrotic enteritis in broiler chickens. Poult. Sci. 2022, 101, 101988. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Osorio, L.M.; Yepes-Medina, V.; Ballou, A.; Parini, M.; Angel, R. Short and Medium Chain Fatty Acids and Their Derivatives as a Natural Strategy in the Control of Necrotic Enteritis and Microbial Homeostasis in Broiler Chickens. Front. Vet. Sci. 2021, 8, 773372. [Google Scholar] [CrossRef] [PubMed]
- Diaz Carrasco, J.M.; Redondo, L.M.; Redondo, E.A.; Dominguez, J.E.; Chacana, A.P.; Fernandez Miyakawa, M.E. Use of Plant Extracts as an Effective Manner to Control Clostridium perfringens Induced Necrotic Enteritis in Poultry. Biomed. Res. Int. 2016, 3278359. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, M.H.; Khalifeh, M.S.; Nawasreh, A.N.; Hananeh, W.M.; Awawdeh, M.S. Assessment of Immune Response and Efficacy of Essential Oils Application on Controlling Necrotic Enteritis Induced by Clostridium perfringens in Broiler Chickens. Molecules 2021, 26, 4527. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacilliaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Coman, M.M.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Boyko, N.; Cresci, A. In vitro evaluation of antimicrobial activity of Lactobacillus rhamnosus IMC 501(®), Lactobacillus paracasei IMC 502(®) and SYNBIO(®) against pathogens. J. Appl. Microbiol. 2014, 117, 518–527. [Google Scholar] [CrossRef]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on the growth performance and intestinal health of broilers challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2018, 9, 25. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Q.; Yang, E.; Yan, L.; Li, T.; Zhuang, H. Antimicrobial Compounds Produced by Vaginal Lactobacillus crispatus Are Able to Strongly Inhibit Candida albicans Growth, Hyphal Formation and Regulate Virulence-related Gene Expressions. Front. Microbiol. 2017, 8, 564. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Ren, L.Q.; Zhou, Y.; Ye, B.C. Characterization of antimicrobial activity of three Lactobacillus plantarum strains isolated from Chinese traditional dairy food. Food. Sci. Nutr. 2019, 7, 1997–2005. [Google Scholar] [CrossRef]
- Yazhini, P.; Visha, P.; Selvaraj, P.; Vasanthakumar, P.; Chandran, V. Dietary encapsulated probiotic effect on broiler serum biochemical parameters. Vet. World 2018, 11, 1344–1348. [Google Scholar] [CrossRef]
- Khabirov, A.; Avzalov, R.; Tsapalova, G.; Andreeva, A.; Basharov, A. Effect of a probiotic containing lactobacilli and bifidobacterial on the metabolic processes, litter microbiocenosis, and production indicators of broiler Pekin ducklings. Vet. World 2022, 15, 998–1005. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Markazi, A.; Mortada, M.; Ng, T.T.; Applegate, T.J.; Bielke, L.R.; Syed, B.; Pender, C.M.; Curry, S.; Murugesan, G.R.; et al. Research Note: Effect of synbiotic supplementation on caecal Clostridium perfringens load in broiler chickens with different necrotic enteritis challenge models. Poult. Sci. 2020, 99, 2452–2458. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Tahoun, A.; Rizk, A.M.; Suzuki, T.; Elmonir, W.; Nassef, E.; Shukry, M.; Germoush, M.O.; Farrag, F.; Bin-Jumah, M.; et al. Evaluation of Bifidobacteria and Lactobacillus Probiotics as Alternative Therapy for Salmonella typhimurium Infection in Broiler Chickens. Animals 2020, 10, 1023. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The Gut Microbiota of Laying Hens and Its Manipulation with Prebiotics and Probiotics to Enhance Gut Health and Food Safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]
- Van der Klein, S.; Yibarek, A.; Gibbs, K. A comparison between two application strategies of a dual strain waterline probiotic during a milk dicrotic enteritis challenge. In Proceedings of the Poultry Science Association, 112th Annual Meeting Abstracts, Philadelphia, PA, USA, 10–13 July 2023; Abstract No. 173. pp. 84–85. [Google Scholar]
- Velge, P.; Bottreau, E.; Quere, P.; Pardon, P.; Nicolle, J.C.; Morisson, M.; Bout, D.; Dimier, I. Establishment and characterization of partially differentiated chicken enterocyte cell clones. Eur. J. Cell Biol. 2002, 81, 203–212. [Google Scholar] [CrossRef]
- Carboni, J.M.; Howe, C.L.; Barwick, K.W.; Mooseker, M.S.; Morrow, J.S. Characterization of intestinal brush border cytoskeletal proteins of normal and neoplastic human epithelial cells. Am. J. Pathol. 1987, 129, 589–600. [Google Scholar]
- Han, X.; Bertzbach, L.D.; Veit, M. Mimicking the passage of avian influenza viruses through the gastrointestinal tract of chickens. Vet. Microbiol. 2019, 239, 108462. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Kolenda, R.; Moman Khan, M.; Weinreich, J.; Li, G.; Wieler, L.H.; Tedin, K.; Roggenbuch, D.; Shchierack, P. Novel avian pathogenic Escherichia coli genes responsible for adhesion to chicken and human cell lines. Appl. Environ. Microbiol. 2020, 86, e01068-20. [Google Scholar] [CrossRef]
- Kolenda, R.; Burdukiewicz, M.; Wimonć, M.; Aleksandrowicz, A.; Ali, A.; Szabo, I.; Tedin, K.; Bartholdson Scott, J.; Pickard, D.; Schierack, P. Identification of natural mutations responsible for altered infection phenotypes of Salmonella enterica clinical isolates by using cell line infection screens. Appl. Environ. Microbiol. 2021, 87, e02177-20. [Google Scholar] [CrossRef]
- Paradowska, M.; Dunislawska, A.; Siwek, M.; Slawinska, A. Avian Cell Culture Models to Study Immunomodulatory Properties of Bioactive Products. Animals 2022, 12, 670. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, W.H.; Lee, S.J.; Lillehoj, H.S. Detection of chicken interleukin-10 production in intestinal epithelial cells and necrotic enteritis induced by Clostridium perfringens using capture ELISA. Vet. Immunol. Immunopathol. 2018, 204, 52–58. [Google Scholar] [CrossRef]
- Shen, C.; Christensen, L.G.; Bak, S.Y.; Christensen, N.; Kragh, K. Immunomodulatory effects of thymol and cinnamaldehyde in chicken cell lines. J. Appl. Anim. Nutr. 2020, 8, 21–30. [Google Scholar] [CrossRef]
- Bonis, V.; Rossell, C.; Gehart, H. The Intestinal Epithelium—Fluid Fate and Rigid Structure from Crypt Bottom to Villus Tip. Front. Cell Dev. Biol. 2021, 9, 661931. [Google Scholar] [CrossRef]
- Letourneau, J.; Levesque, C.; Berthiaume, F.; Jacques, M.; Mourez, M. In Vitro Assay of Bacterial Adhesion onto Mammalian Epithelial Cells. J. Vis. Exp. 2011, 51, e2783. [Google Scholar]
- Marks, H.; Grzeskowiak, L.; Martinez-Vallespin, B.; Dietz, H.; Zentek, J. Porcine and Chicken Intestinal Epithelial Cell Models for Screening Phytogenic Feed Additives-Chances and Limitations in Use as Alternatives to Feeding Trials. Microorganisms 2022, 10, 629. [Google Scholar] [CrossRef]
- Beaumont, M.; Blanc, F.; Cherbuy, C.; Egidy, G.; Giuffra, E.; Lacroix-Lamand, S.; Wiedemann, A. Intestinal organoids in farm animals. Vet. Res. 2021, 52, 33. [Google Scholar] [CrossRef]
- Nash, T.J.; Morris, K.M.; Mabbott, N.A.; Vervelde, L. Inside-out chicken enteroids with leukocyte component as a model to study host-pathogen interactions. Commun. Biol. 2021, 4, 377. [Google Scholar] [CrossRef] [PubMed]
- Llanco, L.A.; Nakano, V.; Moraes, C.T.P.; Piazza, R.M.F.; Avila-Campos, M.J. Adhesion and invasion of Clostridium perfringens type A into epithelial cells. Braz. J. Microbiol. 2017, 48, 764–768. [Google Scholar] [CrossRef]
- McClane, B.A. Clostridium perfringens type C isolates rapidly upregulate their toxin production upon contact with host cells. New insights into virulence? Virulence 2010, 1, 97–100. [Google Scholar] [CrossRef]
- Latorre, J.D.; Adhikari, B.; Park, S.H.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Kwon, Y.M.; Ricke, S.C.; et al. Evaluation of the Epithelial Barrier Function and Ileal Microbiome in an Established Necrotic Enteritis Challenge Model in Broiler Chickens. Front. Vet. Sci. 2018, 5, 199. [Google Scholar] [CrossRef] [PubMed]
- Trejo, F.M.; Minnaard, J.; Perez, P.F.; De Antoni, G.L. Inhibition of Clostridium difficile growth and adhesion to enterocytes by Bifidobacterium supernatants. Anaerobe 2006, 12, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Cerda, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef]
- Parker, C.T.; Sperandio, V. Cell-to-cell signalling during pathogenesis. Cell. Microbiol. 2009, 11, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Lepp, D.; Zhou, Y.; Ojha, S.; Mehdizadeh Gohari, I.; Carere, J.; Yang, C.; Prescott, J.F.; Gong, J. Clostridium perfringens produces an adhesive pilus required for the pathogenesis of necrotic enteritis in poultry. J. Bacteriol. 2021, 203, e00578-20. [Google Scholar] [CrossRef] [PubMed]
- Lepp, D.; Gong, J.; Songer, J.G.; Boerlin, P.; Parreira, V.R.; Prescott, J.F. Identification of accessory genome regions in poultry Clostridium perfringens isolates carrying the netB plasmid. J. Bacteriol. 2013, 195, 1152–1166. [Google Scholar] [CrossRef]
- Akerele, G.; Al Hakeem, W.G.; Lourenco, J.; Selvaraj, R.K. The Effect of Necrotic Enteritis Challenge on Production Performance, Cecal Microbiome, and Cecal Tonsil Transcriptome in Broilers. Pathogens 2022, 11, 839. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.A.; Koval, M. Specificity of interaction between Clostridium perfringens enterotoxin and claudin-family tight junction proteins. Toxins 2010, 2, 1595–1611. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.; Morris, W.E.; Loidl, C.F.; Tironi-Farinati, C.; McClane, B.A.; Uzal, F.A.; Fernandez Miyakawa, M.E. Clostridium perfringens epsilon toxin increases the small intestinal permeability in mice and rats. PLoS ONE 2009, 4, e7065. [Google Scholar] [CrossRef]
- Seike, S.; Takehara, M.; Kobayashi, K.; Nagahama, M. Clostridium perfringens Delta-Toxin Damages the Mouse Small Intestine. Toxins 2019, 11, 232. [Google Scholar] [CrossRef]
- Seike, S.; Takehara, M.; Takagishi, T.; Miyamoto, K.; Kobayashi, K.; Nagahama, M. Delta-toxin from Clostridium perfringens perturbs intestinal epithelial barrier function in caco-2 cell monolayers. Biochim. Biophys. Acta (BBA)—Biomembr. 2018, 1860, 428–433. [Google Scholar] [CrossRef]
- Morhardt, T.L.; Hayashi, A.; Ochi, T.; Quiros, M.; Kitamoto, S.; Nagao-Kitamoto, H.; Kuffa, P.; Atarashi, K.; Honda, K.; Kao, J.Y.; et al. IL-10 produced by macrophages regulates epithelial integrity in the small intestine. Sci. Rep. 2019, 9, 1223. [Google Scholar] [CrossRef]
- John, D.A.; Williams, L.K.; Kanamarlapudi, V.; Humphrey, T.J.; Wilkinson, T.S. The bacterial species Campylobacter jejuni induce diverse innate immune responses in human and avian intestinal epithelial cells. Front. Microbiol. 2017, 8, 1840. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.A.; McClane, B.A.; Uzal, F.A. Mechanisms of Action and Cell Death Associated with Clostridium perfringens Toxins. Toxins 2018, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Moe, P.C.; Heuck, A.P. Phospholipid hydrolysis caused by Clostridium perfringens alpha-toxin facilitates the targeting of perfringolysin O to membrane bilayers. Biochemistry 2010, 49, 9498–9507. [Google Scholar] [CrossRef] [PubMed]
- Khalique, A.; Zeng, D.; Shoaib, M.; Wang, H.; Qing, X.; Rajput, D.S.; Pan, K.; Ni, X. Probiotics mitigating subclinical necrotic enteritis (SNE) as potential alternatives to antibiotics in poultry. AMB Express 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Garde, S.; Gómez-Torres, N.; Hernández, M.; Ávila, M. Susceptibility of Clostridium perfringens to antimicrobials produced by lactic acid bacteria: Reuterin and nisin. Food Control 2014, 44, 22–25. [Google Scholar] [CrossRef]
- Meyer, M.M.; Fries-Craft, K.A.; Bobeck, E.A. Composition and inclusion of probiotics in broiler diets alter intestinal permeability and spleen immune cell profiles without negatively affecting performance. J. Anim. Sci. 2020, 98, skz383. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zeng, Y.; Zeng, D.; Wang, H.; Sun, N.; Xin, J.; Zhou, M.; Yang, H.; Lei, L.; Ling, H.; et al. Dietary probiotic supplementation suppresses subclinical necrotic enteritis in broiler chickens in a microbiota-dependent manner. Sec. Microb. Immunol. 2022, 13, 855426. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Dharmaprakash, V.; Nighot, P.; Guo, S.; Nighot, M.; Do, T.; Ma, T.Y. Bifidobacterium bifidum Enhances the Intestinal Epithelial Tight Junction Barrier and Protects against Intestinal Inflammation by Targeting the Toll-like Receptor-2 Pathway in an NF-kappaB-Independent Manner. Int. J. Mol. Sci. 2021, 22, 8070. [Google Scholar] [CrossRef]
- Rao, R.K.; Samak, G. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [PubMed]
- Emami, N.K.; Calik, A.; White, M.B.; Young, M.; Dalloul, R.A. Necrotic Enteritis in Broiler Chickens: The Role of Tight Junctions and Mucosal Immune Responses in Alleviating the Effect of the Disease. Microorganisms 2019, 7, 231. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin production: A probiotic trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Wei, Z.; Yang, W.; Huang, J.; Yang, Y.; Wang, J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front. Oncol. 2022, 12, 985363. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, T.; Takaesu, G.; Ninomiya-Tsuji, J.; Shibuya, H.; Gaynor, R.B.; Matsumoto, K. Role of the TAB2-related protein TAB3 in IL-1 and TNF signaling. EMBO J. 2003, 22, 6277–6288. [Google Scholar] [CrossRef] [PubMed]
- Fasina, Y.O.; Lillehoj, H.S. Characterization of intestinal immune response to Clostridium perfringens infection in broiler chickens. Poult. Sci. 2019, 98, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Liu, D.; Zhang, B.; Li, Z.; Li, Y.; Ding, B.; Guo, Y. Two Lactobacillus Species Inhibit the Growth and alpha-Toxin Production of Clostridium perfringens and Induced Proinflammatory Factors in Chicken Intestinal Epithelial Cells In Vitro. Front. Microbiol. 2017, 8, 2081. [Google Scholar] [CrossRef]
- Voloshin, N.; Tyurin-Kuzmin, P.; Karagyaur, M.; Akopyan, Z.; Kulebyakin, K. Practical Use of Immortalized Cells in Medicine: Current Advances and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 12716. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. perfringens Strain No. | Country of Origin and Year of Isolation | Expression of Gene Encoding Virulence Factor | |
|---|---|---|---|
| α Toxin | NetB Toxin | ||
| CP1 | UK—2017 | + | - |
| CP10 | SWE—2018 | + | - |
| 25037-CP01 | USA—2016 | + | + |
| CP22 | USA—2016 | + | + |
| Treatment No. | Treatment Details | Data Expressed as: C, % Cytotoxicity, or RC, % Reduction in Cytotoxicity |
|---|---|---|
| Baseline treatments; CHIC-8E11 cells treated with… | ||
| 1 | Medium control (at 10, 20, 30, 40 and 50 µL/mL) | C |
| 2 | CFS L. acidophilus AG01 (at 10, 20, 30, 40 and 50 µL/mL) | C |
| 3 | CFS B. animalis subsp. lactis AG02 (at 10, 20, 30, 40 and 50 µL/mL) | C |
| 4 | CFS 25037-CP01 (at 10, 20, 30, 40 and 50 µL/mL) | C |
| 5 | CFS CP22 (at 10, 20, 30, 40 and 50 µL/mL) | C |
| 6 | NetB toxin (at 1 µg/mL) | C |
| Experimental treatments; CHIC-8E11 cells treated with… | ||
| 7 | CFS L. acidophilus AG01 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment, followed by 4 h contact time with 30 µL/mL of 25037-CP01 CFS | RC vs. T4 (at 30 µL/mL) |
| 8 | CFS L. acidophilus AG01 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment, followed by 4 h contact time with 30 µL/mL of CP22 CFS | RC vs. T5 (at 30 µL/mL) |
| 9 | CFS B. animalis AG02 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment followed by 4 h contact time with 30 µL/mL of 25037-CP01 CFS | RC vs. T4 (at 30 µL/mL) |
| 10 | CFS B. animalis AG02 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment, followed by 4 h contact time with 30 µL/mL of CP22 CFS | RC vs. T5 (at 30 µL/mL) |
| 11 | CFS L. acidophilus AG01 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment, followed by 4 h contact time with 1 µL/mL of NetB toxin | RC vs. T6 (at 1 µL/mL) |
| 12 | CFS B. animalis AG02 (at 10, 20, 30, 40 and 50 µL/mL) overnight pre-treatment, followed by 4 h contact time with 31 µL/mL of NetB toxin | RC vs. T6 (at 1 µL/mL) |
| Response Measure | Treatment No. | Dose of Application to CHIC-8E11 Cells | |||||
|---|---|---|---|---|---|---|---|
| 1 µg/mL | 10 µL/mL | 20 µL/mL | 30 µL/mL | 40 µL/mL | 50 µL/mL | ||
| Cytotoxicity (%) of baseline treatments (±SD) | 1 | nd | 0 (0.04) | 0 (0.04) | 0 (0.04) | 0 (0.04) | 0 (0.06) |
| 2 | nd | 1.12 (0.05) | 6.65 (0.17) | 4.64 (0.06) | 10.3 (0.09) | 11.17 (0.28) | |
| 3 | nd | 7.78 (0.28) | 7 (0.41) | 9.52 (0.28) | 6.2 (0.32) | 7.26 (0.35) | |
| 4 | nd | 34.57 (0.06) | 44.73 (0.03) | 55.77 (0.04) | 51.71 (0.03) | 51.15 (0.02) | |
| 5 | nd | 45.22 (0.11) | 57.78 (0.02) | 63.04 (0.05) | 65.19 (0.12) | 61.70 (0.02) | |
| 6 | 74.60 (0.04) | nd | nd | nd | nd | nd | |
| Reduction in cytotoxicity (%) induced by probiotic pre-treatment vs. baseline treatment 1 (±SD) | 7 | nd | −7.75 (0.02) | −1.47 (0.16) | 13.31 (3.59) | 15.50 (2.02) | 24.39 (3.37) |
| 8 | nd | −3.71 (0.67) | 16.85 (2.50) | 9.01 (0.34) | 13.48 (2.54) | 24.42 (4.75) | |
| 9 | nd | −8.45 (0.69) | 2.15 (0.04) | 5.72 (0.79) | 13.84 (5.40) | 16.07 (2.40) | |
| 10 | nd | −19.92 (0.18) | 0.78 (0.37) | 10.78 (0.02) | 19.60 (0.3) | 21.23 (0.33) | |
| 11 | nd | 4.10 (0.25) | 5.21 (0.59) | 17.80 (1.36) | 10.56 (1.07) | 15.50 (0.005) | |
| 12 | nd | 20.24 (2.08) | 23.02 (1.02) | 26.70 (3.32) | 29.22 (0.20) | 33.76 (3.75) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadekar, D.; Udrea, A.C.; Bak, S.Y.; Christensen, N.; Gibbs, K.; Shen, C.; Bernardeau, M. Cell-Free Culture Supernatant of Lactobacillus acidophilus AG01 and Bifidobacterium animalis subsp. lactis AG02 Reduces the Pathogenicity of NetB-Positive Clostridium perfringens in a Chicken Intestinal Epithelial Cell Line. Microorganisms 2024, 12, 839. https://doi.org/10.3390/microorganisms12040839
Kadekar D, Udrea AC, Bak SY, Christensen N, Gibbs K, Shen C, Bernardeau M. Cell-Free Culture Supernatant of Lactobacillus acidophilus AG01 and Bifidobacterium animalis subsp. lactis AG02 Reduces the Pathogenicity of NetB-Positive Clostridium perfringens in a Chicken Intestinal Epithelial Cell Line. Microorganisms. 2024; 12(4):839. https://doi.org/10.3390/microorganisms12040839
Chicago/Turabian StyleKadekar, Darshana, Andreea Cornelia Udrea, Steffen Yde Bak, Niels Christensen, Kirsty Gibbs, Chong Shen, and Marion Bernardeau. 2024. "Cell-Free Culture Supernatant of Lactobacillus acidophilus AG01 and Bifidobacterium animalis subsp. lactis AG02 Reduces the Pathogenicity of NetB-Positive Clostridium perfringens in a Chicken Intestinal Epithelial Cell Line" Microorganisms 12, no. 4: 839. https://doi.org/10.3390/microorganisms12040839