

Exercise Affects Mucosa-Associated Microbiota and Colonic Tumor Formation Induced by Azoxymethane in High-Fat-Diet-Induced Obese Mice
 , , and
, , and
Abstract
:1. Introduction
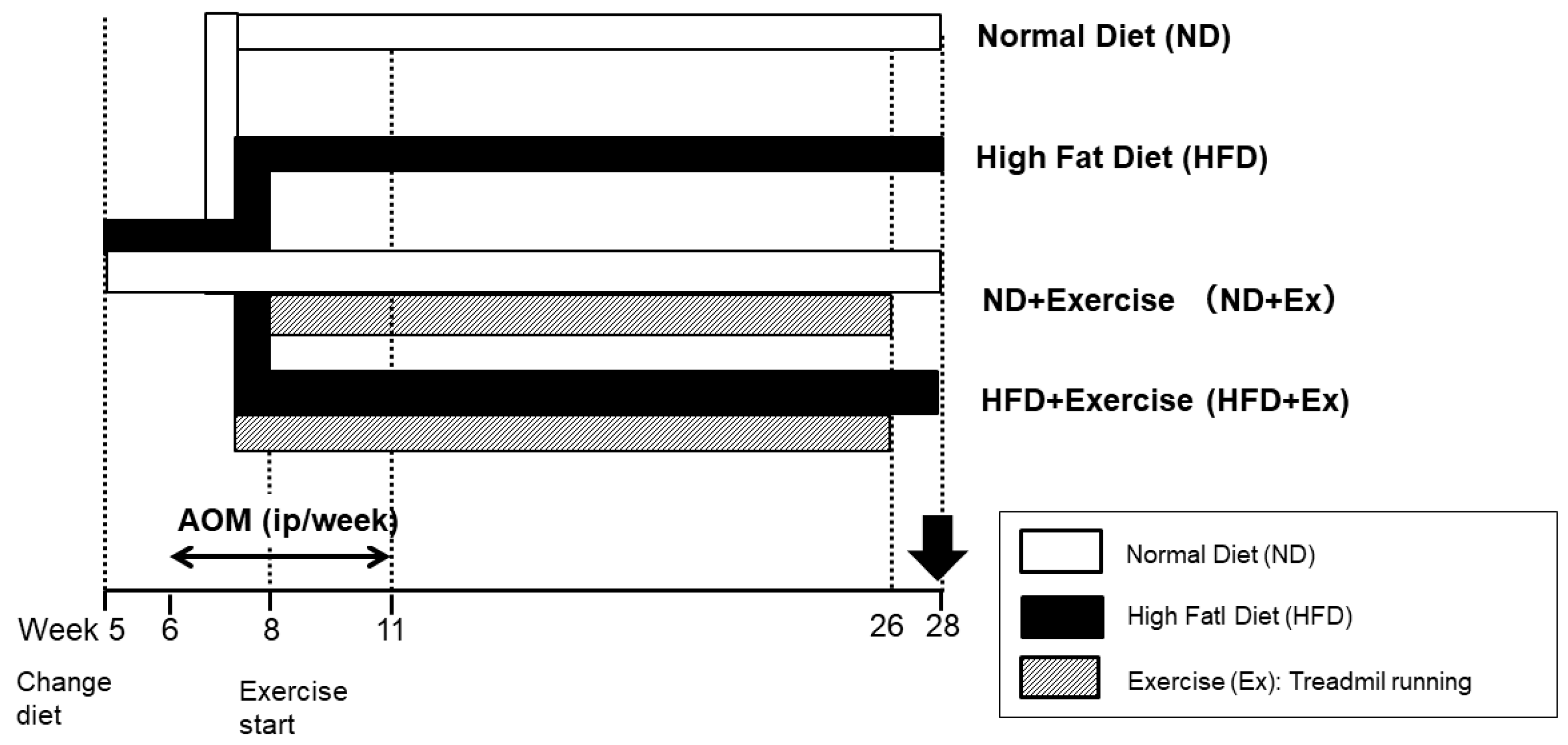
2. Materials and Methods
Statistical Analysis
3. Results
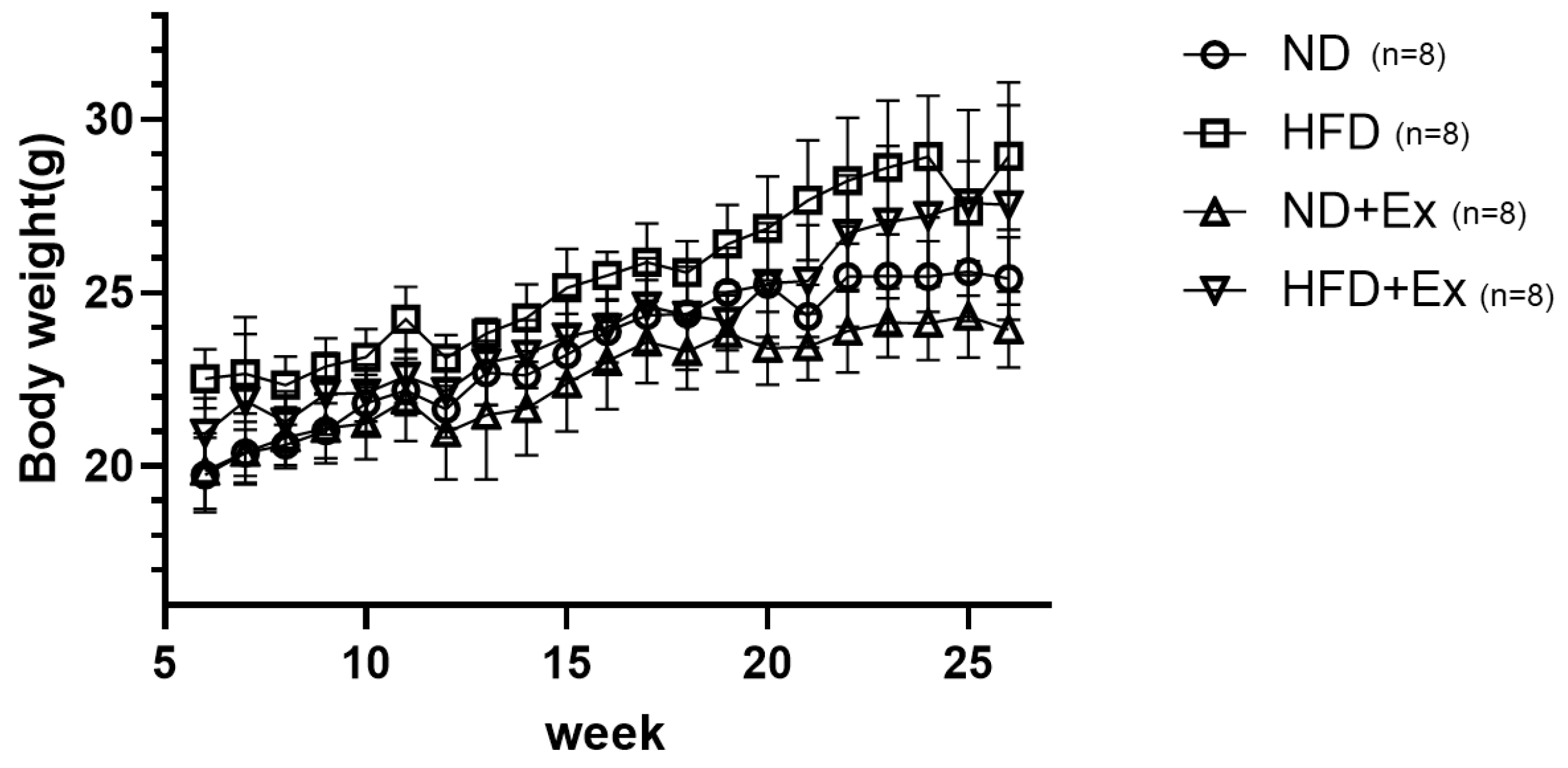
3.1. Mouse Body Weight Change by Diet and Exercise
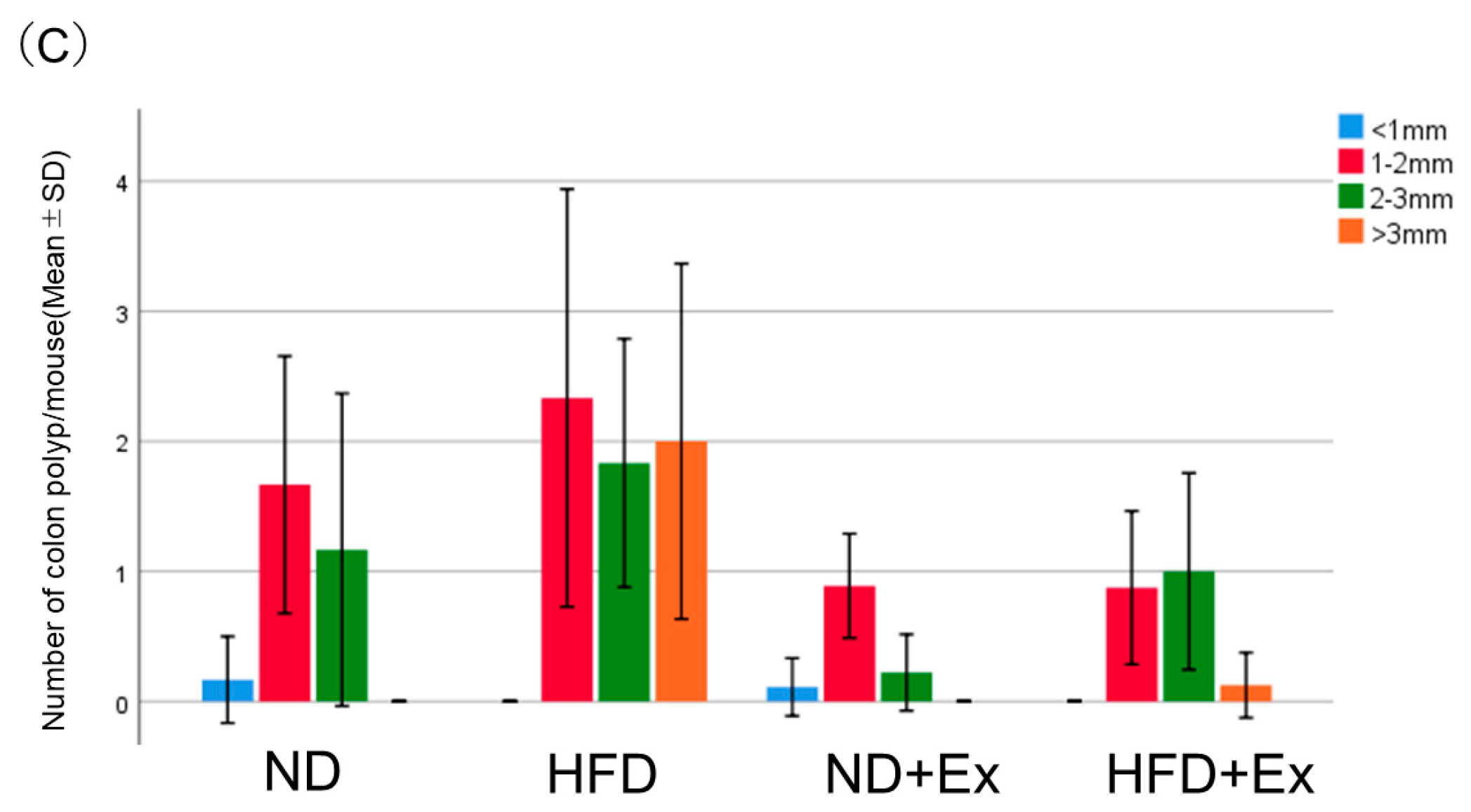
3.2. AOM-Induced Colorectal Tumor Count
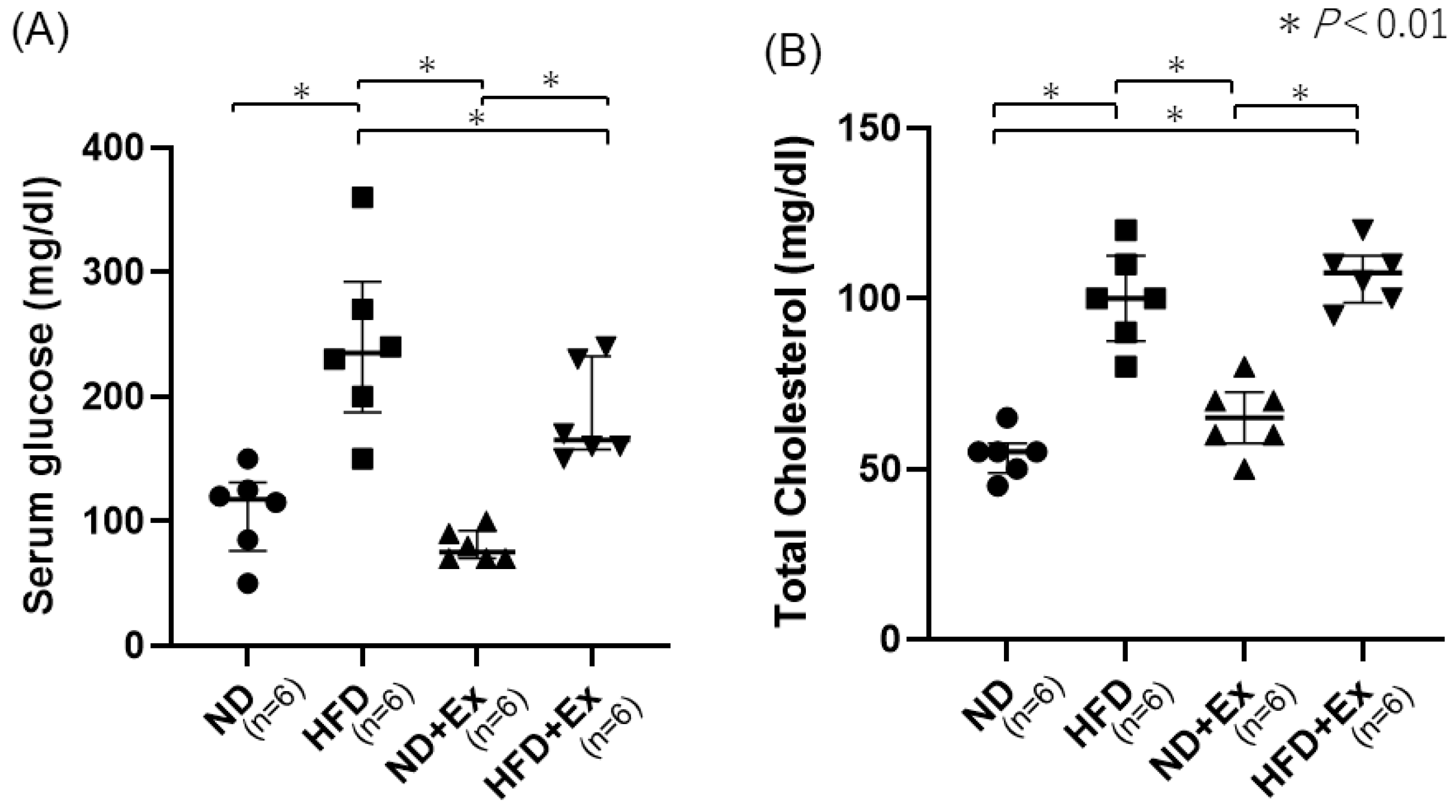
3.3. Blood Glucose (BS) and Total Cholesterol Levels
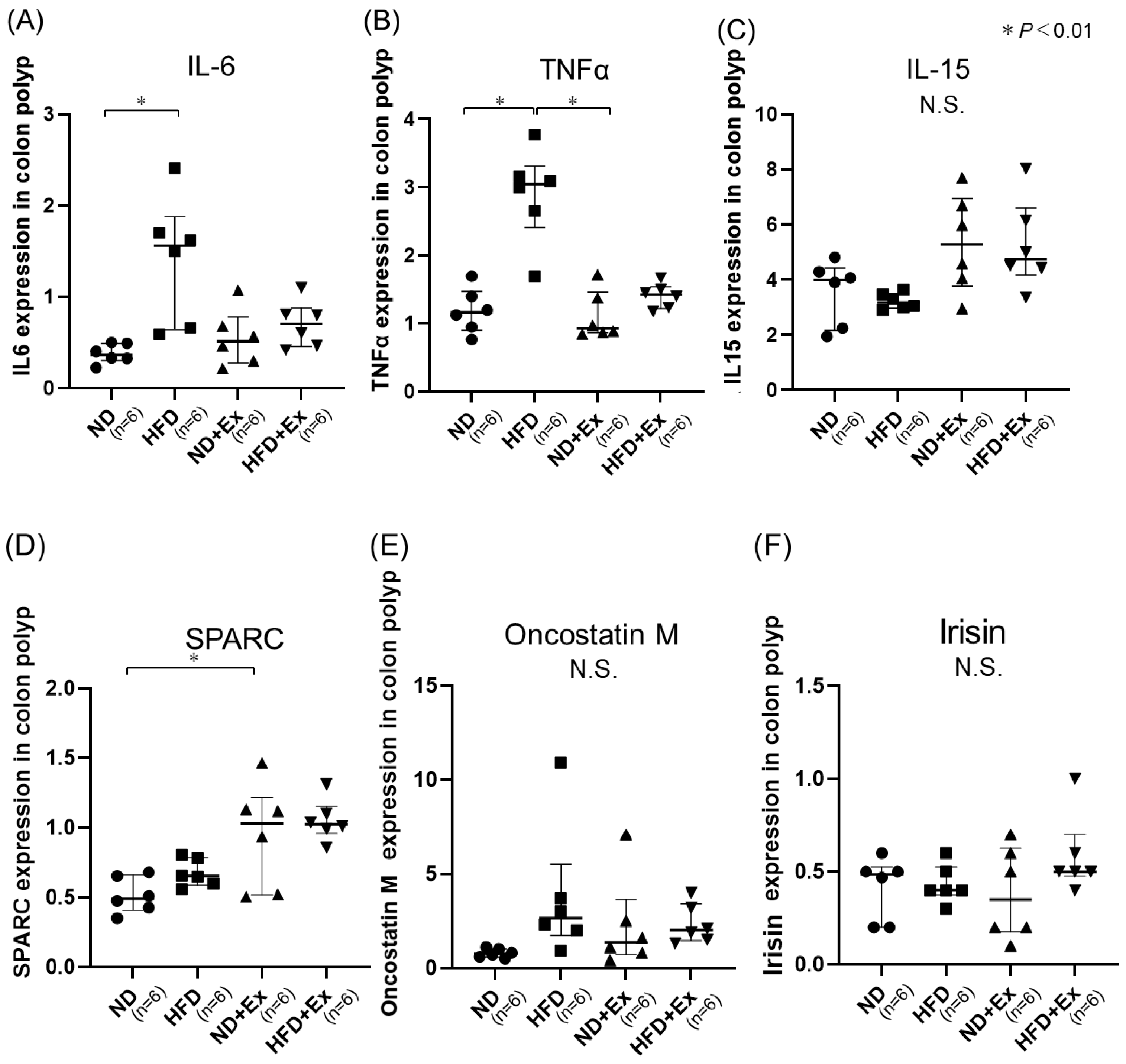
3.4. Cytokine and Myokine Expression in Colonic Tumors
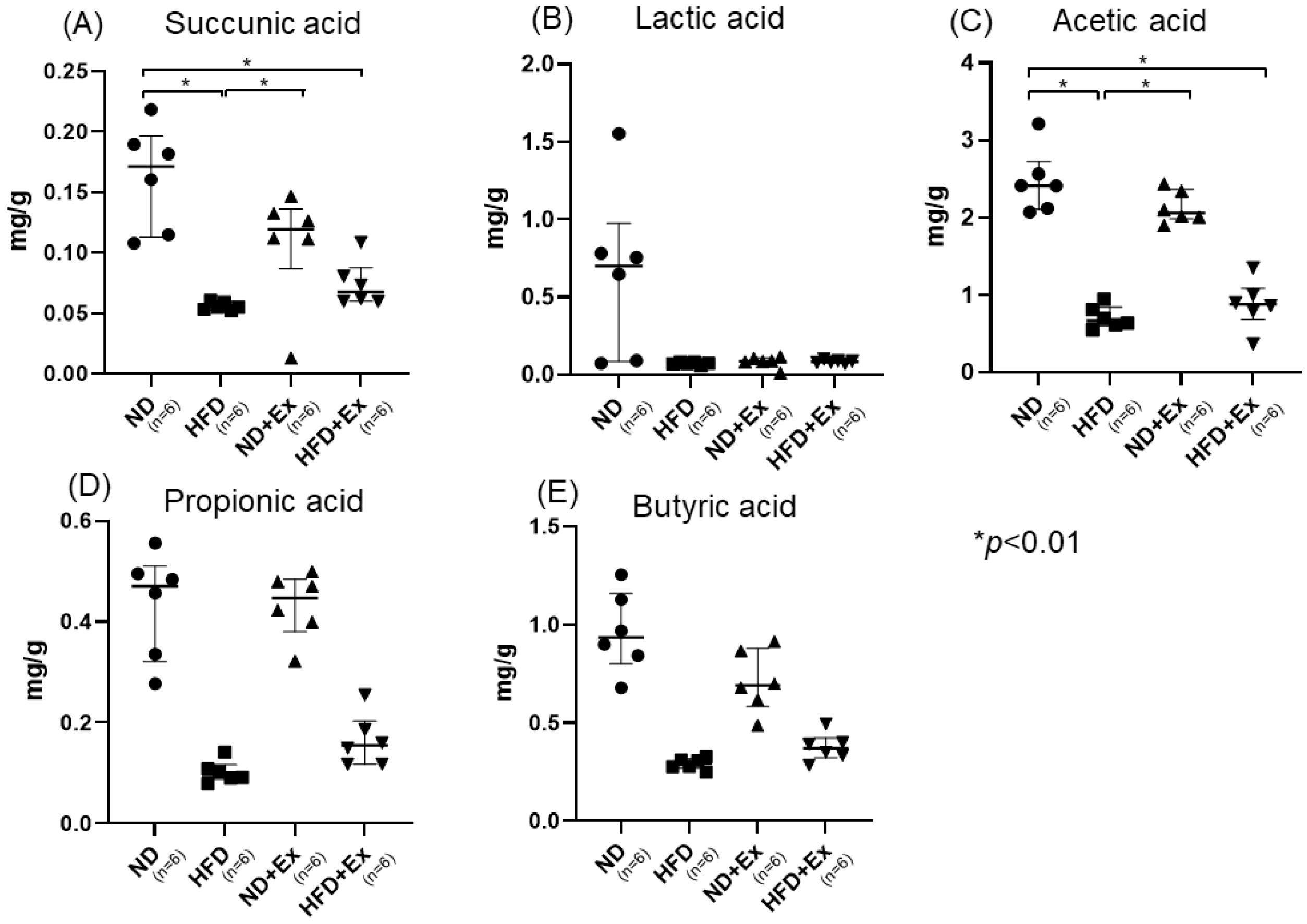
3.5. Short-Chain Fatty Acid (SFA) and Nonconjugated Bile Acids Analyses of Feces
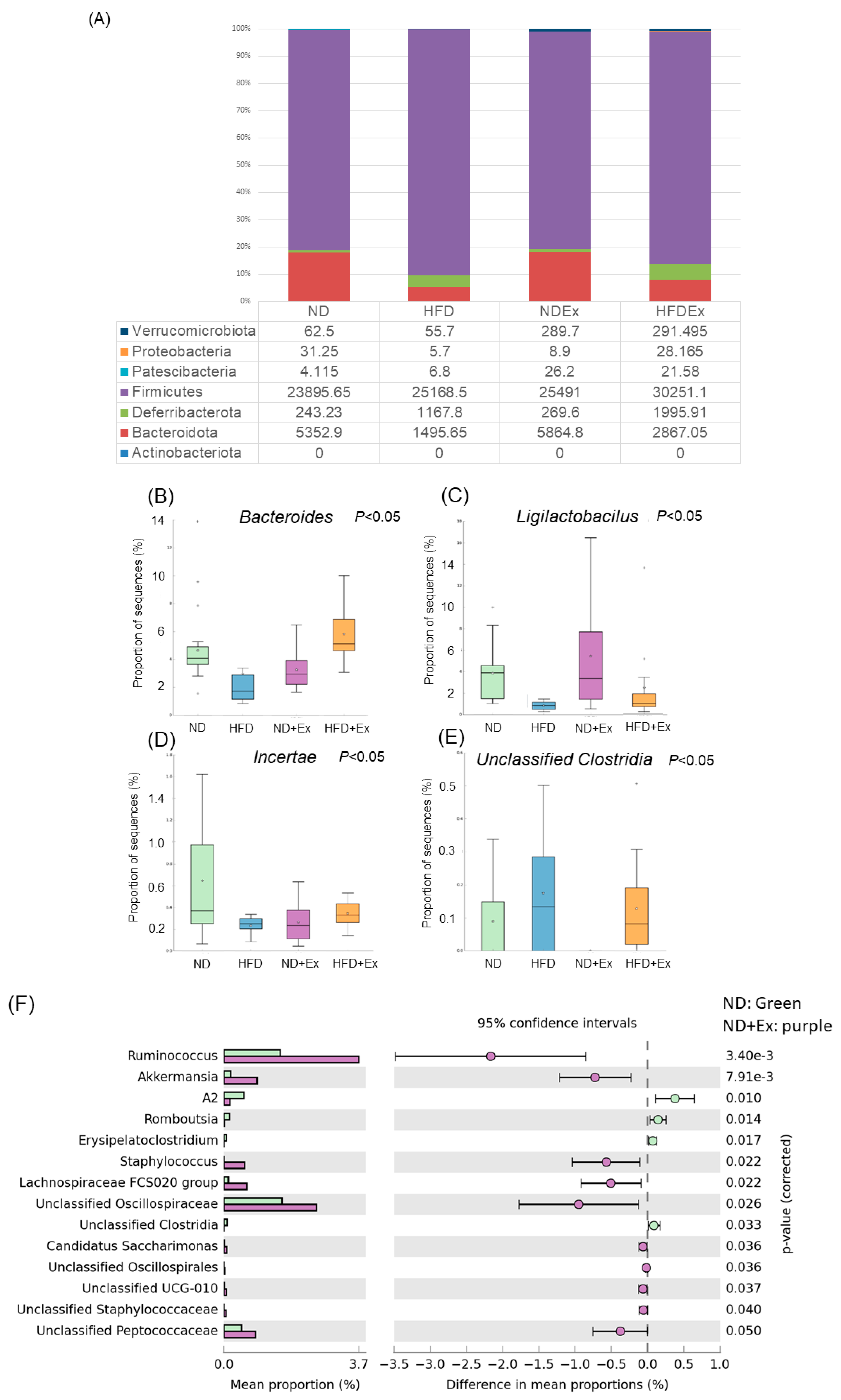
3.6. Changes in Fecal Microbiota
3.7. Changes in MAM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Keum, N.; Giovannucci, E. Global Burden of Colorectal Cancer: Emerging Trends, Risk Factors and Prevention Strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.H.; Yu, J. Gut Microbiota in Colorectal Cancer: Mechanisms of Action and Clinical Applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic Analysis of Faecal Microbiome as a Tool towards Targeted Non-Invasive Biomarkers for Colorectal Cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef]
- Nakatsu, G.; Li, X.; Zhou, H.; Sheng, J.; Wong, S.H.; Wu, W.K.K.; Ng, S.C.; Tsoi, H.; Dong, Y.; Zhang, N.; et al. Gut Mucosal Microbiome across Stages of Colorectal Carcinogenesis. Nat. Commun. 2015, 6, 8727. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Clevers, H. Gut Microbiota in Colorectal Cancer: Associations, Mechanisms, and Clinical Approaches. Annu. Rev. Cancer Biol. 2022, 6, 65–84. [Google Scholar] [CrossRef]
- Wong, C.C.; Yu, J. Gut Microbiota in Colorectal Cancer Development and Therapy. Nat. Rev. Clin. Oncol. 2023, 20, 429–452. [Google Scholar] [CrossRef]
- Zhang, Z.; Bahaji Azami, N.L.; Liu, N.; Sun, M. Research Progress of Intestinal Microecology in the Pathogenesis of Colorectal Adenoma and Carcinogenesis. Technol. Cancer Res. Treat. 2023, 22, 15330338221135938. [Google Scholar] [CrossRef]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Van Citters, G.W.; Lin, H.C. Management of Small Intestinal Bacterial Overgrowth. Curr. Gastroenterol. Rep. 2005, 7, 317–320. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Vieira, A.R.; Abar, L.; Chan, D.S.M.; Vingeliene, S.; Polemiti, E.; Stevens, C.; Greenwood, D.; Norat, T. Foods and Beverages and Colorectal Cancer Risk: A Systematic Review and Meta-Analysis of Cohort Studies, an Update of the Evidence of the WCRF-AICR Continuous Update Project. Ann. Oncol. 2017, 28, 1788–1802. [Google Scholar] [CrossRef]
- Papadimitriou, N.; Dimou, N.; Tsilidis, K.K.; Banbury, B.; Martin, R.M.; Lewis, S.J.; Kazmi, N.; Robinson, T.M.; Albanes, D.; Aleksandrova, K.; et al. Physical Activity and Risks of Breast and Colorectal Cancer: A Mendelian Randomisation Analysis. Nat. Commun. 2020, 11, 597. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Lee, I.-M.; Weiderpass, E.; Campbell, P.T.; Sampson, J.N.; Kitahara, C.M.; Keadle, S.K.; Arem, H.; Berrington de Gonzalez, A.; Hartge, P.; et al. Association of Leisure-Time Physical Activity With Risk of 26 Types of Cancer in 1.44 Million Adults. JAMA Intern. Med. 2016, 176, 816–825. [Google Scholar] [CrossRef]
- Morris, J.S.; Bradbury, K.E.; Cross, A.J.; Gunter, M.J.; Murphy, N. Physical Activity, Sedentary Behaviour and Colorectal Cancer Risk in the UK Biobank. Br. J. Cancer 2018, 118, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Kyu, H.H.; Bachman, V.F.; Alexander, L.T.; Mumford, J.E.; Afshin, A.; Estep, K.; Lennert Veerman, J.; Delwiche, K.; Iannarone, M.L.; Moyer, M.L.; et al. Physical Activity and Risk of Breast Cancer, Colon Cancer, Diabetes, Ischemic Heart Disease, and Ischemic Stroke Events: Systematic Review and Dose-Response Meta-Analysis for the Global Burden of Disease Study 2013. BMJ 2016, 354, i3857. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; You, Y.; Huang, J.; Guan, C.; Chen, Z.; Fang, M.; Yao, F.; Han, J. Association between Physical Activity and Digestive-System Cancer: An Updated Systematic Review and Meta-Analysis. J. Sport Health Sci. 2021, 10, 4–13. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, S.; Zhou, W.; Hu, D.; Xu, H.; Ji, G. Secondary Bile Acids and Tumorigenesis in Colorectal Cancer. Front. Oncol. 2022, 12, 813745. [Google Scholar] [CrossRef] [PubMed]
- Ocvirk, S.; O’Keefe, S.J.D. Dietary Fat, Bile Acid Metabolism and Colorectal Cancer. Semin. Cancer Biol. 2021, 73, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wei, H.; Zhou, Y.; Szeto, C.-H.; Li, C.; Lin, Y.; Coker, O.O.; Lau, H.C.H.; Chan, A.W.H.; Sung, J.J.Y.; et al. High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites. Gastroenterology 2022, 162, 135–149.e2. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Boytar, A.N.; Nitert, M.D.; Morrision, M.; Skinner, T.L.; Jenkins, D.G. Exercise-Induced Changes to the Human Gut Microbiota and Implications for Colorectal Cancer: A Narrative Review. J. Physiol. 2022, 600, 5189–5201. [Google Scholar] [CrossRef]
- Allen, J.M.; Mailing, L.J.; Cohrs, J.; Salmonson, C.; Fryer, J.D.; Nehra, V.; Hale, V.L.; Kashyap, P.; White, B.A.; Woods, J.A. Exercise Training-Induced Modification of the Gut Microbiota Persists after Microbiota Colonization and Attenuates the Response to Chemically-Induced Colitis in Gnotobiotic Mice. Gut Microbes 2018, 9, 115–130. [Google Scholar] [CrossRef]
- Queipo-Ortuño, M.I.; Seoane, L.M.; Murri, M.; Pardo, M.; Gomez-Zumaquero, J.M.; Cardona, F.; Casanueva, F.; Tinahones, F.J. Gut Microbiota Composition in Male Rat Models under Different Nutritional Status and Physical Activity and Its Association with Serum Leptin and Ghrelin Levels. PLoS ONE 2013, 8, e65465. [Google Scholar]
- Cheng, D.; Xie, M.Z. A Review of a Potential and Promising Probiotic candidate—Akkermansia Muciniphila. J. Appl. Microbiol. 2021, 130, 1813–1822. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-Inflammatory Properties of Two Akkermansia Muciniphila Strains on Chronic Colitis in Mice. Front. Cell Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Exercise Immunology: Practical Applications. Int. J. Sports Med. 1997, 18 (Suppl. 1), S91–S100. [Google Scholar] [CrossRef] [PubMed]
- Hagar, A.; Wang, Z.; Koyama, S.; Serrano, J.A.; Melo, L.; Vargas, S.; Carpenter, R.; Foley, J. Endurance Training Slows Breast Tumor Growth in Mice by Suppressing Treg Cells Recruitment to Tumors. BMC Cancer 2019, 19, 536. [Google Scholar] [CrossRef]
- Grosicki, G.J.; Langan, S.P.; Bagley, J.R.; Galpin, A.J.; Garner, D.; Hampton-Marcell, J.T.; Allen, J.M.; Robinson, A.T. Gut Check: Unveiling the Influence of Acute Exercise on the Gut Microbiota. Exp. Physiol. 2023, 108, 1466–1480. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Guo, B.; Sui, H.; Chen, J.; Zhang, L.; Lv, C.; Li, B. The Effects of Aerobic Exercise on the Intestinal Tumors and Flora of the ApcMin/+ Mouse. Clin. Transl. Oncol. 2022, 24, 305–318. [Google Scholar] [CrossRef]
- Lai, Z.-L.; Tseng, C.-H.; Ho, H.J.; Cheung, C.K.Y.; Lin, J.-Y.; Chen, Y.-J.; Cheng, F.-C.; Hsu, Y.-C.; Lin, J.-T.; El-Omar, E.M.; et al. Fecal Microbiota Transplantation Confers Beneficial Metabolic Effects of Diet and Exercise on Diet-Induced Obese Mice. Sci. Rep. 2018, 8, 15625. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Wang, M.; Song, Y.; Shi, Y. A Bibliometric Analysis on the Research Trend of Exercise and the Gut Microbiome. Microorganisms 2023, 11, 903. [Google Scholar] [CrossRef]
- Williams, E.A.; Coxhead, J.M.; Mathers, J.C. Anti-Cancer Effects of Butyrate: Use of Micro-Array Technology to Investigate Mechanisms. Proc. Nutr. Soc. 2003, 62, 107–115. [Google Scholar] [CrossRef]
- Wallace, K.; Baron, J.A.; Karagas, M.R.; Cole, B.F.; Byers, T.; Beach, M.A.; Pearson, L.H.; Burke, C.A.; Silverman, W.B.; Sandler, R.S. The Association of Physical Activity and Body Mass Index with the Risk of Large Bowel Polyps. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2082–2086. [Google Scholar] [CrossRef]
- Matsumoto, M.; Inoue, R.; Tsukahara, T.; Ushida, K.; Chiji, H.; Matsubara, N.; Hara, H. Voluntary Running Exercise Alters Microbiota Composition and Increases N-Butyrate Concentration in the Rat Cecum. Biosci. Biotechnol. Biochem. 2008, 72, 572–576. [Google Scholar] [CrossRef]
- Nierengarten, M.B. Association of Physical Exercise and a Healthy Gut Microbiome in Colorectal Cancer. Cancer 2023, 129, 655–656. [Google Scholar] [CrossRef]
- Seke Etet, P.F.; Vecchio, L.; Nwabo Kamdje, A.H.; Mimche, P.N.; Njamnshi, A.K.; Adem, A. Physiological and Environmental Factors Affecting Cancer Risk and Prognosis in Obesity. Semin. Cancer Biol. 2023, 94, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Beale, M.N.; Leach, H.J.; Baxter, B.A.; Smith, H.V.; Lyden, K.; Ryan, E.P. Physical Activity and Stool Metabolite Relationships Among Adults at High Risk for Colorectal Cancer. J. Phys. Act. Health 2021, 18, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.Y.; Sun, Y.; Wang, C. Diversity analysis of luminal and mucosa-associated microbial communities in obese mice. Zhonghua Nei Ke Za Zhi 2018, 57, 743–748. [Google Scholar] [PubMed]
- Xu, S.-S.; Wang, N.; Huang, L.; Zhang, X.-L.; Feng, S.-T.; Liu, S.-S.; Wang, Y.; Liu, Z.-G.; Wang, B.-Y.; Wu, T.-W.; et al. Changes in the Mucosa-Associated Microbiome and Transcriptome across Gut Segments Are Associated with Obesity in a Metabolic Syndrome Porcine Model. Microbiol. Spectr. 2022, 10, e0071722. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Xie, G.; Jia, W. Bile Acid-Microbiota Crosstalk in Gastrointestinal Inflammation and Carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence | Length | |
|---|---|---|---|
| IL-6 | FP | CGGCCTTCCCTACTTCACAAGTCCG | 66 |
| RP | CAGGTCTGTTGGGAGTGGTATCC | ||
| TNF-alpha | FP | CCACCATCAAGGACTCAAATGG | 74 |
| RP | CCTTTGCAGAACTCAGGAATGGACATTCG | ||
| IL-15 | FP | CATCCATCTCGTGCTACTTGTG | 112 |
| RP | GCCTCTGTTTTAGGGAGACCT | ||
| SPARC | FP | CCACACGTTTCTTTGAGACC | 95 |
| RP | GATGTCCTGCTCCTTGATGC | ||
| Oncostain M | FP | GTGGCTGCTCCAACTCTTCC | 81 |
| RP | AGAGTGATTCTGTGTTCCCCGT | ||
| Irisin | FP | GAGCCCAATAACAACAAGG | 242 |
| RP | GAGGATAATAAGCCCGATG | ||
| Actb | FP | CACTGTCGAGTCGCGTCC | 102 |
| RP | CGCAGCGATATCGTCATCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yo, S.; Matsumoto, H.; Gu, T.; Sasahira, M.; Oosawa, M.; Handa, O.; Umegaki, E.; Shiotani, A. Exercise Affects Mucosa-Associated Microbiota and Colonic Tumor Formation Induced by Azoxymethane in High-Fat-Diet-Induced Obese Mice. Microorganisms 2024, 12, 957. https://doi.org/10.3390/microorganisms12050957
Yo S, Matsumoto H, Gu T, Sasahira M, Oosawa M, Handa O, Umegaki E, Shiotani A. Exercise Affects Mucosa-Associated Microbiota and Colonic Tumor Formation Induced by Azoxymethane in High-Fat-Diet-Induced Obese Mice. Microorganisms. 2024; 12(5):957. https://doi.org/10.3390/microorganisms12050957
Chicago/Turabian StyleYo, Shogen, Hiroshi Matsumoto, Tingting Gu, Momoyo Sasahira, Motoyasu Oosawa, Osamu Handa, Eiji Umegaki, and Akiko Shiotani. 2024. "Exercise Affects Mucosa-Associated Microbiota and Colonic Tumor Formation Induced by Azoxymethane in High-Fat-Diet-Induced Obese Mice" Microorganisms 12, no. 5: 957. https://doi.org/10.3390/microorganisms12050957