Cecal Microbiome Analyses on Wild Japanese Rock Ptarmigans (Lagopus muta japonica) Reveals High Level of Coexistence of Lactic Acid Bacteria and Lactate-Utilizing Bacteria

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
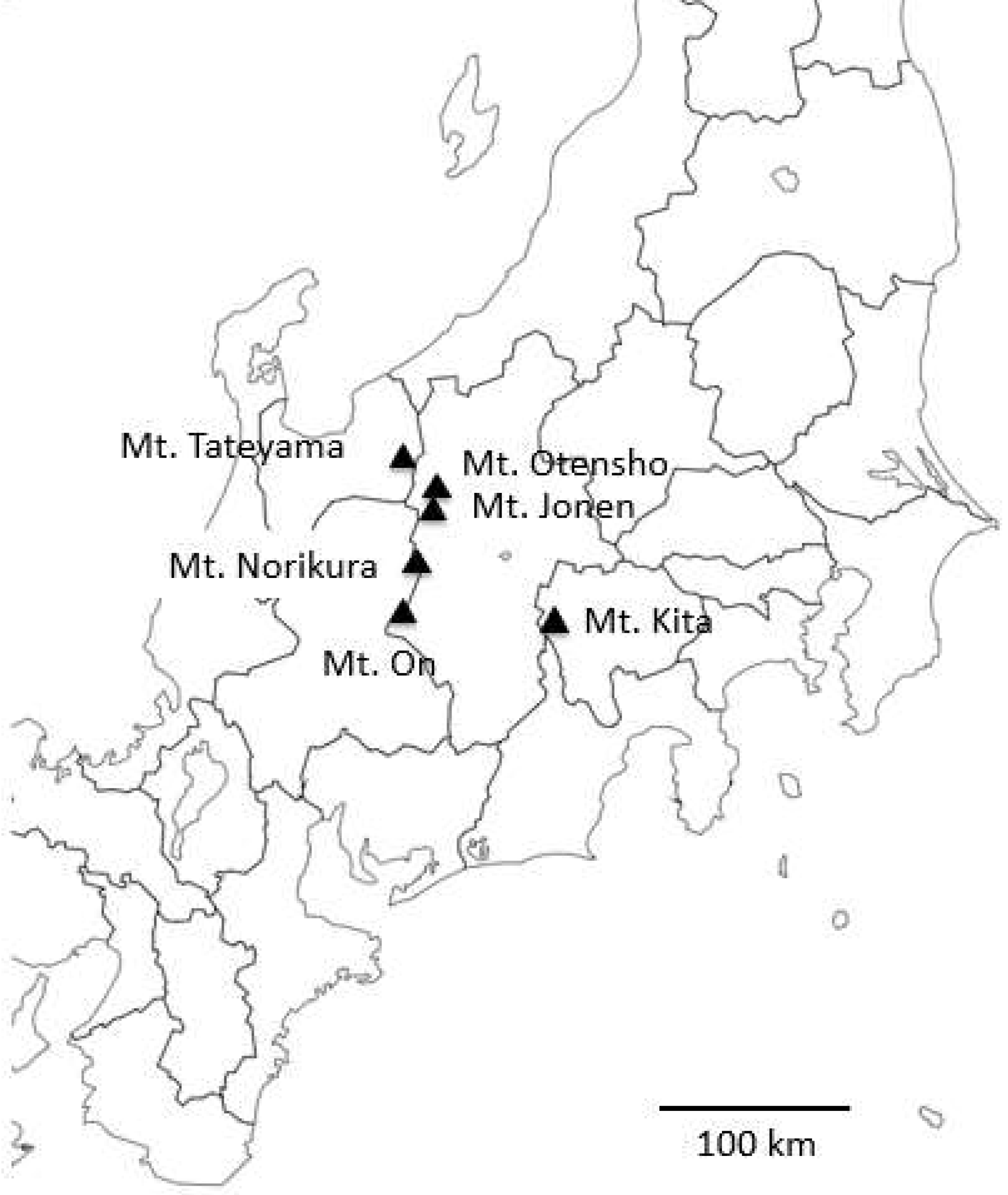
2.1. Sample Collections
2.2. DNA Extractions and High-Throughput Sequencing of 16s rRNA Gene
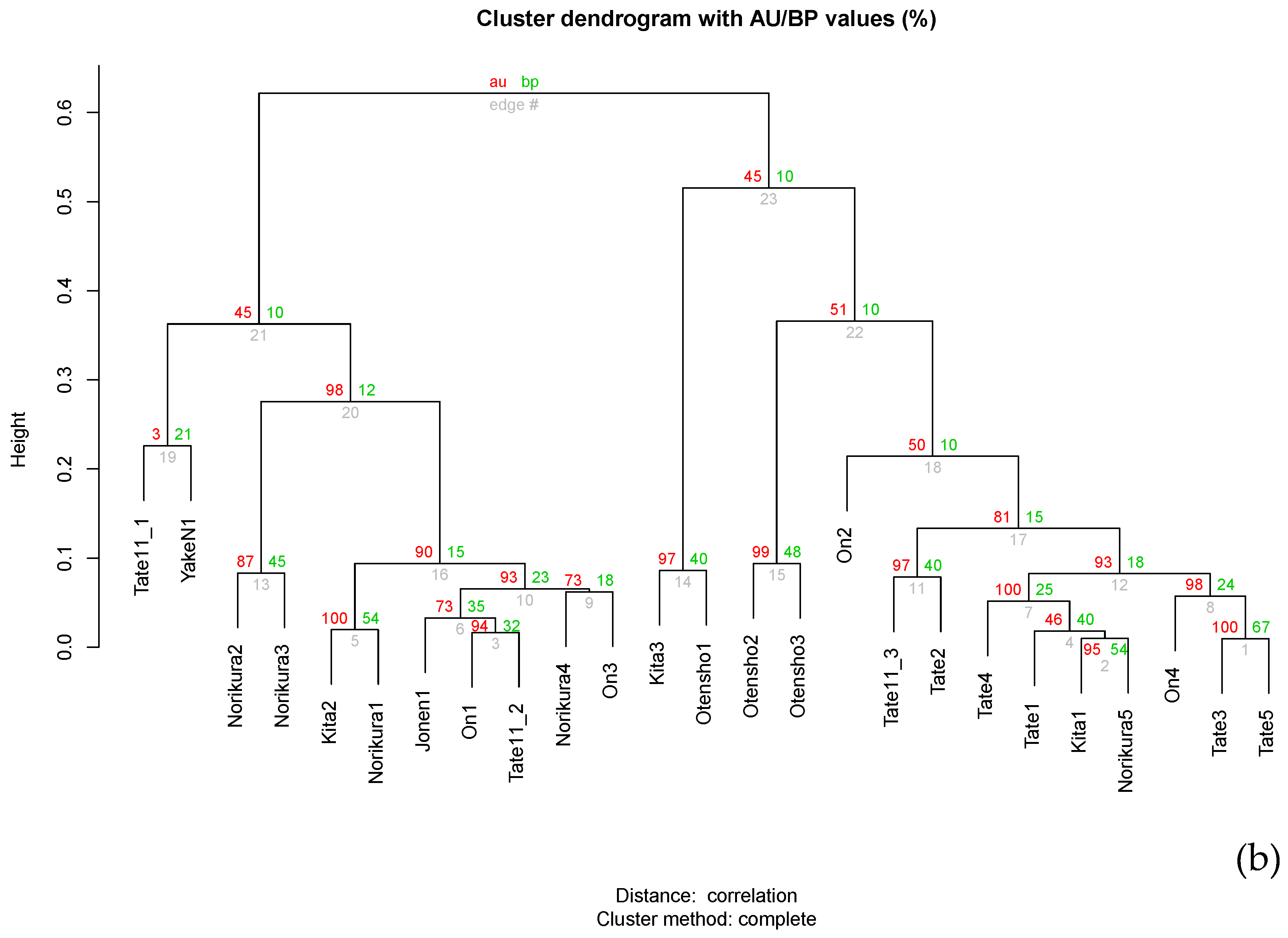
2.3. Phylogenetic Analyses and Community Comparisons
2.4. Ethics
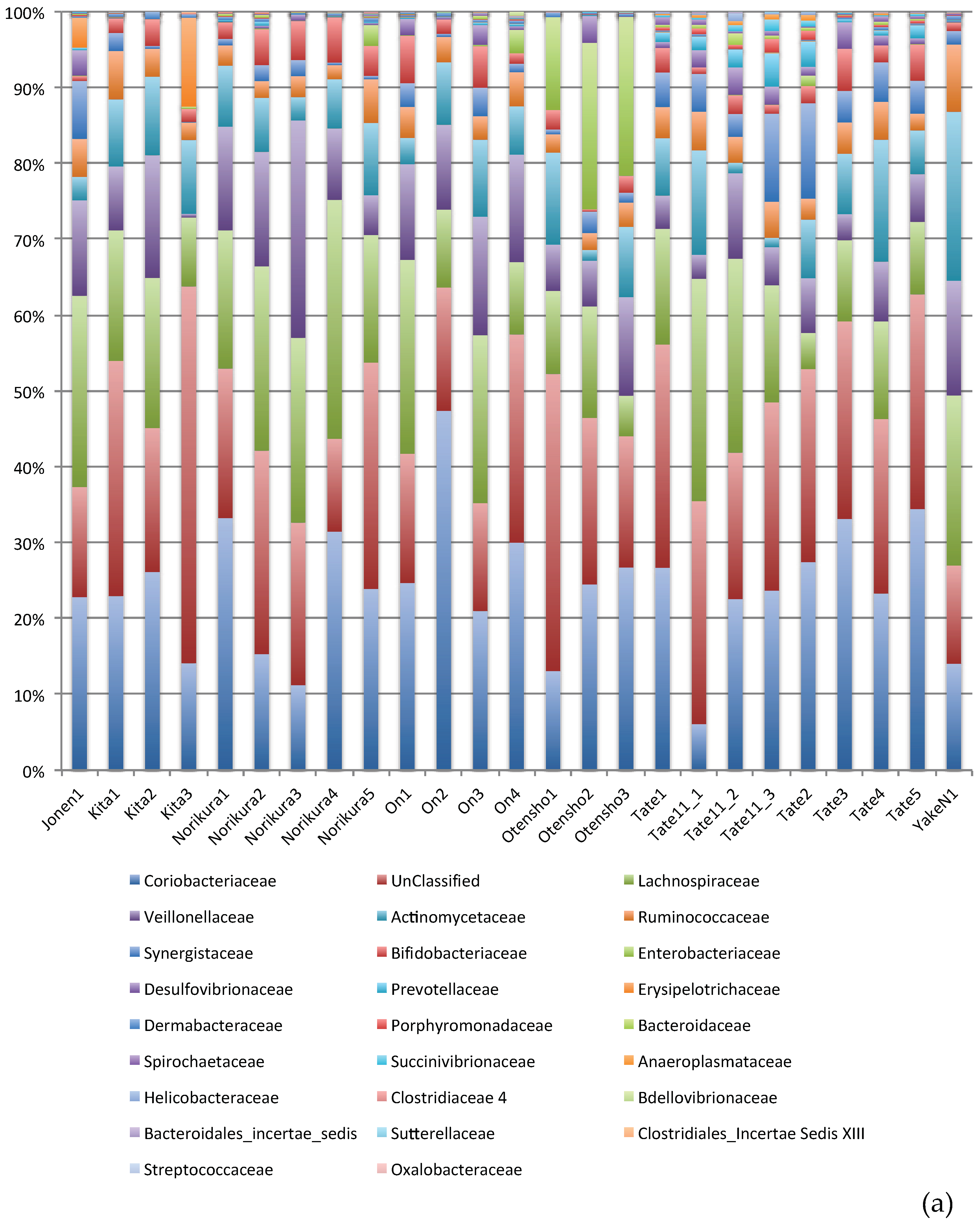
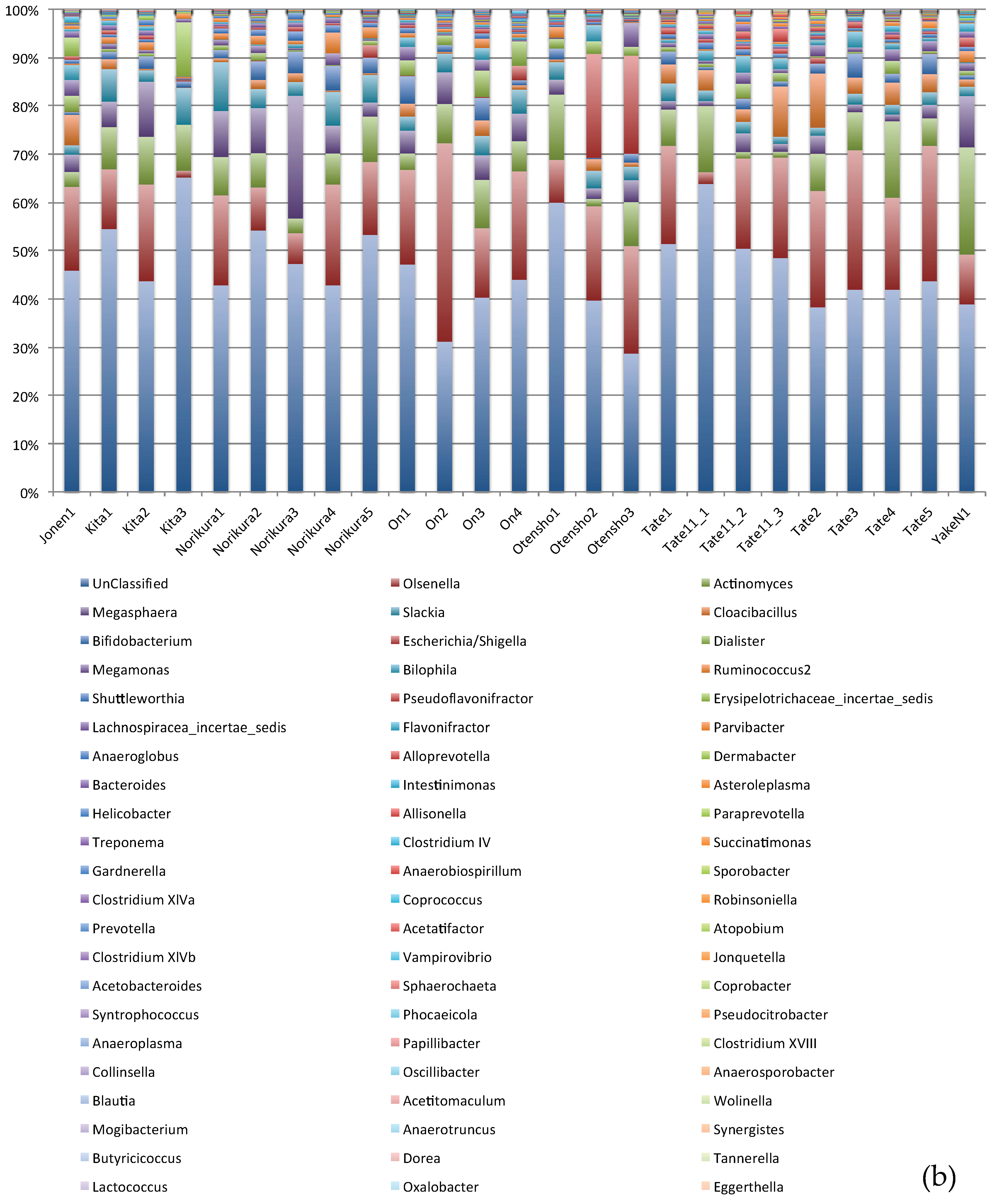
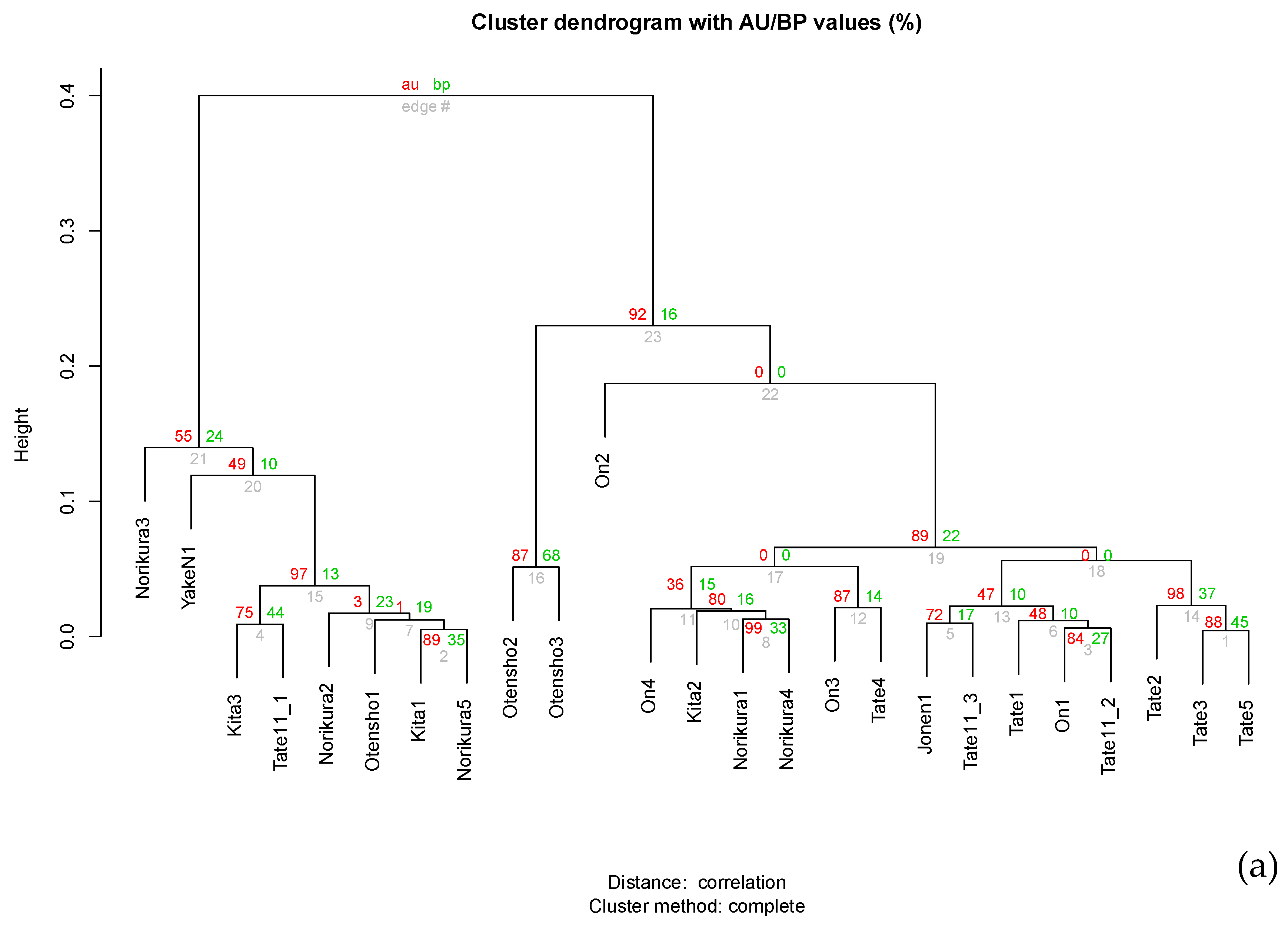
3. Result and Discussions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suzuki, A.; Kobayashi, A.; Nakamura, H.; Takasu, F. Population viability analysis of the Japanese rock ptarmigan Lagopus muta japonica in Japan. Wildl. Biol. 2013, 19, 339–346. [Google Scholar] [CrossRef]
- Nakamura, H. Rock Ptarmigan Lagopus mutus japonicus. Jpn. J. Ornithol. 2007, 56, 93–114. [Google Scholar] [CrossRef]
- Ministry of the Environment, Government of Japan 2012. National Conservation Program for Japanese Rock Ptarmigans. Available online: https://www.env.go.jp/nature/kisho/hogozoushoku/pdf/jigyoukeikaku/raicho.pdf (accessed on 25 July 2018).
- Ushida, K.; Segawa, T.; Tsuchida, S.; Murata, K. Cecal bacterial communities in wild Japanese rock ptarmigans and captive Svalbard rock ptarmigans. J. Vet. Med. Sci. 2016, 78, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchida, S.; Murata, K.; Ohkuma, M.; Ushida, K. Isolation of Streptococcus gallolyticus with very high degradability of condensed tannins from feces of the wild Japanese rock ptarmigans on Mt. Tateyama. J. Gen. Appl. Microbiol. 2017, 63, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Tuschida, S.; Ohara, Y.; Kuramochi, K.; Murata, K.; Ushida, K. Effective degradation of phenolic glycoside rhododendrin and its aglycone ghododendrol by cecal feces of wild Japanese rock ptarmigans. Jpn. J. Zoo Wildl. Med. 2017, 22, 41–45. [Google Scholar]
- Hird, S.M.; Carstens, B.C.; Cardiff, S.W.; Dittmann, D.L.; Brumfield, R.T. Sampling locality is more detectable than taxonomy or ecology in the gut microbiota of the brood-parasitic Brown-headed Cowbird (Molothrus ater). PeerJ 2014, 2, e321. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Fujimaki, Y.; Yoshii, R.; Koike, H. Genetic variability in the mitochondrial control region of the Japanese Rock Ptarmigan Lagopus mutus japonicas. Jpn. J. Ornithol. 2001, 50, 53–64. [Google Scholar] [CrossRef]
- Kobayashi, A.; Nakamura, H. Seasonal change of food items of the Japanese rock ptarmigan. Jpn. J. Ornithol. 2011, 60, 200–215. [Google Scholar] [CrossRef]
- Li, T.H.; Qin, Y.; Sham, P.C.; Lau, K.S.; Chu, K.M.; Leung, W.K. Alterations in Gastric Microbiota After H. Pylori Eradication and in Different Histological Stages of Gastric Carcinogenesis. Sci. Rep. 2017, 7, 44935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okai, S.; Usui, F.; Yokota, S.; Hori-I, Y.; Hasegawa, M.; Nakamura, T.; Kurosawa, M.; Okada, S.; Yamamoto, K.; Nishiyama, E.; et al. High-affinity monoclonal IgA regulates gut microbiota and prevents colitis in mice. Nat. Microbiol. 2016, 1, 16103. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Goto, S.; Mori, H.; Higashi, K.; Hosomichi, K.; Aizawa, N.; Takahashi, N.; Tsuchida, M.; Suzuki, Y.; Yamada, T.; et al. Comprehensive microbiome analysis of tonsillar crypts in IgA nephropathy. Nephrol. Dial. Transplant. 2016, 32, 2072–2079. [Google Scholar] [CrossRef] [PubMed]
- Orla-Jensen, S. La classification des bactéries lactiques. Le Lait 1924, 4, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Dewhirst, F.E.; Paster, B.J.; Tzellas, N.; Coleman, B.; Downes, J.; Spratt, D.A.; Wade, W.G. Characterization of novel human oral isolates and cloned 16S rDNA sequences that fall in the family Coriobacteriaceae: Description of Olsenella gen. nov., reclassification of Lactobacillus uli as Olsenella uli comb. nov. and description of Olsenella profusa sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 1797–1804. [Google Scholar] [PubMed]
- Gutierrez, J.; Davis, R.E.; Lindahl, I.L.; Warwick, E.J. Bacterial Changes in the Rumen During the Onset of Feed-lot Bloat of Cattle and Characteristics of Peptostreptococcus elsdenii n. sp. Appl. Microbiol. 1959, 7, 16–22. [Google Scholar] [PubMed]
- Hashizume, K.; Tsukahara, T.; Yamada, K.; Koyama, H.; Ushida, K. Megasphaera elsdenii JCM1772T normalizes hyperlactate production in the large intestine of fructooligosaccharide-fed rats by stimulating butyrate production. J. Nutr. 2003, 133, 3187–3190. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, T.; Koyama, H.; Okada, M.; Ushida, K. Stimulation of butyrate production by gluconic acid in batch culture of pig cecal digesta and identification of butyrate-producing bacteria. J. Nutr. 2002, 132, 2229–2234. [Google Scholar] [CrossRef] [PubMed]
- Ushida, K.; Hoshi, S.; Ajisaka, K. 13C-NMR studies on lactate metabolism in a porcine gut microbial ecosystem. Microb. Ecol. Health Dis. 2002, 14, 241–246. [Google Scholar]
- Umesaki, Y.; Yajima, T.; Yokokura, T.; Mutai, M. Effect of organic acid absorption on bicarbonate transport in rat colon. Pflüg. Arch. 1979, 379, 43–47. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU Code | Domain | Phylum | Class | Order | Family | Genus | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FCAYKGM7 | Bacteria | 1 | Actinobacteria | 1 | Actinobacteria | 1 | Coriobacteriales | 1 | Coriobacteriaceae | 1 | Olsenella | 0.96 |
| FCAYKGM14 | Bacteria | 1 | Firmicutes | 1 | Clostridia | 1 | Clostridiales | 1 | Lachnospiraceae | 1 | Shuttleworthia | 0.38 |
| FCAYKGM12 | Bacteria | 1 | Actinobacteria | 1 | Actinobacteria | 1 | Actinomycetales | 1 | Actinomycetaceae | 0.92 | Actinomyces | 0.9 |
| FCAYKGM22 | Bacteria | 1 | Firmicutes | 1 | Negativicutes | 1 | Selenomonadales | 1 | Veillonellaceae | 1 | Megasphaera | 0.82 |
| FCAYKGM20 | Bacteria | 1 | Firmicutes | 0.96 | Clostridia | 0.87 | Clostridiales | 0.86 | Eubacteriaceae | 0.2 | Alkalibaculum | 0.18 |
| FCAYKGM29 | Bacteria | 1 | Actinobacteria | 0.96 | Actinobacteria | 0.96 | Coriobacteriales | 0.94 | Coriobacteriaceae | 0.94 | Slackia | 0.67 |
| FCAYKGM30 | Bacteria | 1 | Firmicutes | 0.99 | Clostridia | 0.98 | Clostridiales | 0.98 | Lachnospiraceae | 0.44 | Robinsoniella | 0.23 |
| FCAYKGM50 | Bacteria | 1 | Actinobacteria | 1 | Actinobacteria | 1 | Bifidobacteriales | 1 | Bifidobacteriaceae | 1 | Bifidobacterium | 0.91 |
| FCAYKGM90 | Bacteria | 1 | Firmicutes | 0.91 | Clostridia | 0.87 | Clostridiales | 0.86 | Eubacteriaceae | 0.22 | Alkalibaculum | 0.21 |
| FCAYKGM67 | Bacteria | 1 | Bacteroidetes | 1 | Bacteroidia | 1 | Bacteroidales | 1 | Prevotellaceae | 0.39 | Paraprevotella | 0.32 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, A.; Kobayashi, A.; Tsuchida, S.; Yamada, T.; Murata, K.; Nakamura, H.; Ushida, K. Cecal Microbiome Analyses on Wild Japanese Rock Ptarmigans (Lagopus muta japonica) Reveals High Level of Coexistence of Lactic Acid Bacteria and Lactate-Utilizing Bacteria. Microorganisms 2018, 6, 77. https://doi.org/10.3390/microorganisms6030077
Ueda A, Kobayashi A, Tsuchida S, Yamada T, Murata K, Nakamura H, Ushida K. Cecal Microbiome Analyses on Wild Japanese Rock Ptarmigans (Lagopus muta japonica) Reveals High Level of Coexistence of Lactic Acid Bacteria and Lactate-Utilizing Bacteria. Microorganisms. 2018; 6(3):77. https://doi.org/10.3390/microorganisms6030077
Chicago/Turabian StyleUeda, Atsushi, Atsushi Kobayashi, Sayaka Tsuchida, Takuji Yamada, Koichi Murata, Hiroshi Nakamura, and Kazunari Ushida. 2018. "Cecal Microbiome Analyses on Wild Japanese Rock Ptarmigans (Lagopus muta japonica) Reveals High Level of Coexistence of Lactic Acid Bacteria and Lactate-Utilizing Bacteria" Microorganisms 6, no. 3: 77. https://doi.org/10.3390/microorganisms6030077