Influence of Season and Diet on Fiber Digestion and Bacterial Community Structure in the Rumen of Muskoxen (Ovibos moschatus)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Trial and Feed and Fecal Sampling
2.2. In Situ Digestibility
2.3. Ruminal pH and Sampling
2.4. Chemical Analysis
2.5. Overall Tract Apparent Digestibility Calculation
2.6. DNA Extraction from Ruminal Solids
2.7. Sequence Analysis
2.8. Statistical Analysis
3. Results
3.1. Feed Composition
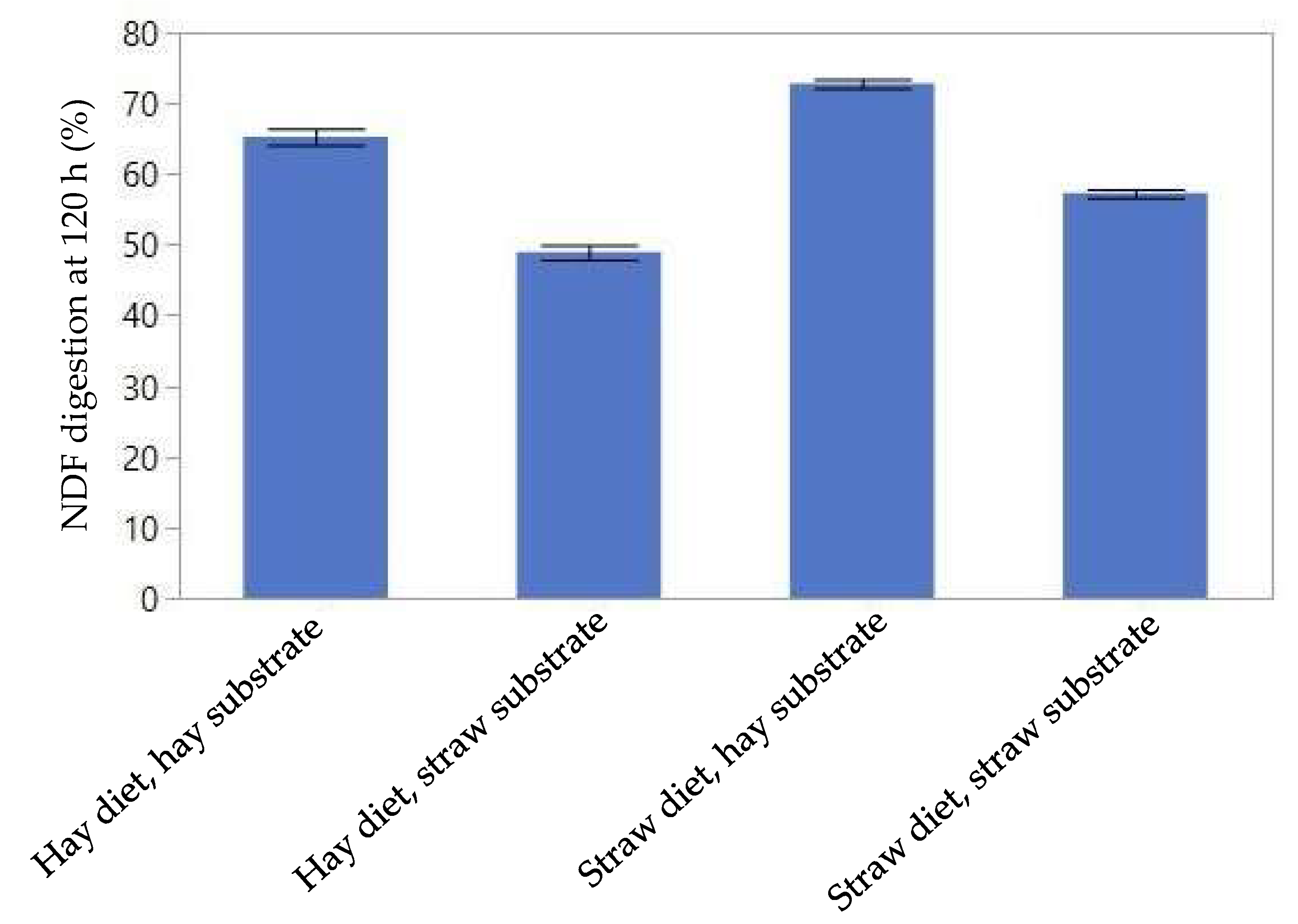
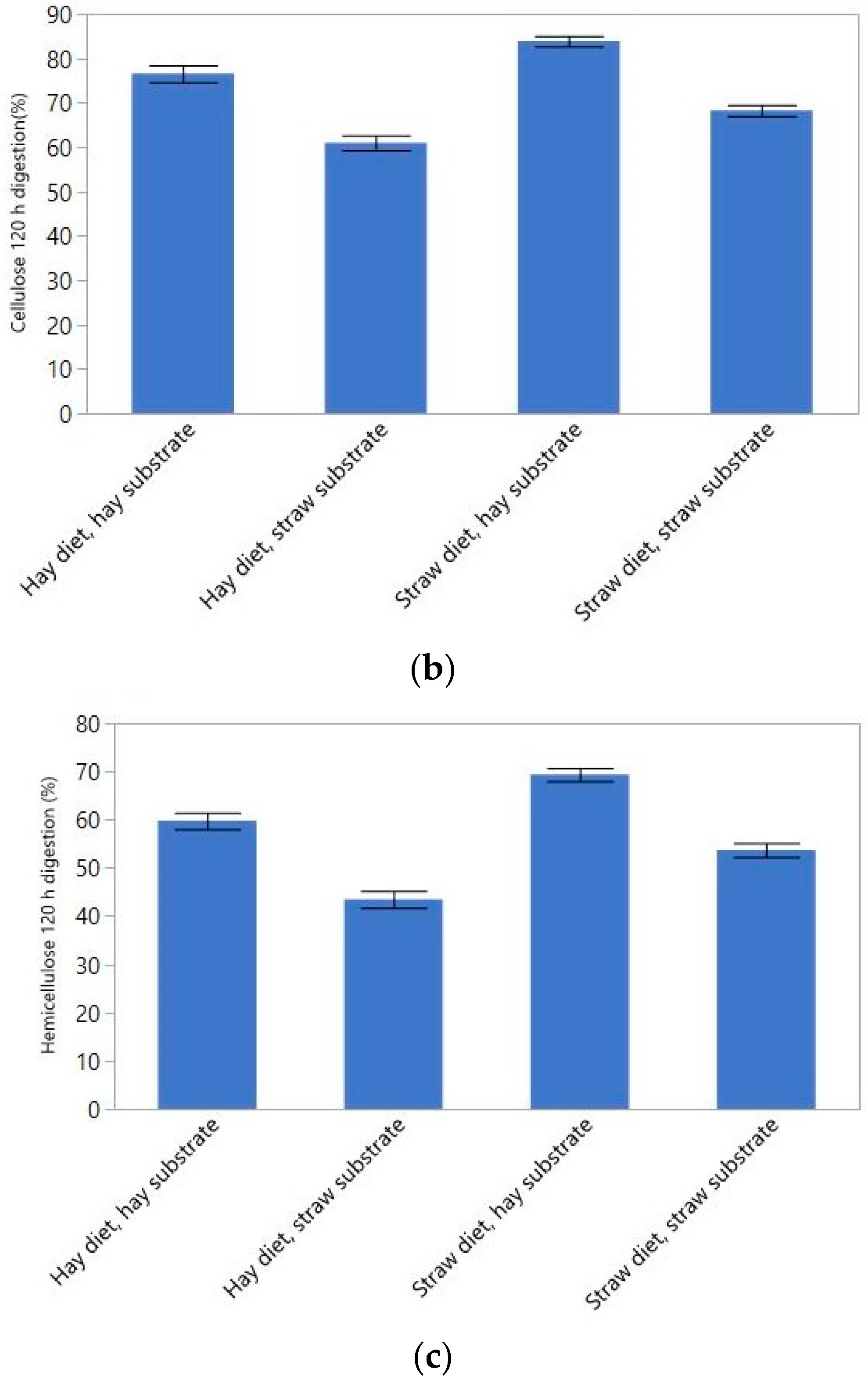
3.2. Ruminal Fiber Digestion Kinetics
3.3. Ruminal pH
3.4. Bacterial Community Analyses
3.5. Association between Fiber Digestion Kinetics and the Proportion of Bacterial Sequences
4. Discussion
4.1. Intake, Digestion and Body Mass Changes
4.2. Bacterial Community
4.3. Implications for Muskoxen Biology
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Attwood, G.; Kelly, W.; Altermann, E.; Moon, C.; Leahy, S.; Cookson, A. Application of rumen microbial genome information to livestock systems in the postgenomic era. Aust. J. Exp. Agric. 2008, 48, 695–700. [Google Scholar] [CrossRef]
- Wallace, R.J. Gut microbiology—Broad genetic diversity, yet specific metabolic niches. Animal 2008, 2, 661–668. [Google Scholar] [CrossRef]
- Barboza, P.S.; Bennett, A.; Lignot, J.-H.; Mackie, R.I.; McWhorter, T.J.; Secor, S.M.; Skovgaard, N.; Sundset, M.A.; Wang, T. Digestive Challenges for Vertebrate Animals: Microbial Diversity, Cardiorespiratory Coupling, and Dietary Specialization. Physiol. Biochem. Zool. 2010, 83, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Hume, I.D.; Hummel, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 2010, 4, 979–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, A.J.; Poppi, D.P. Nutritional Ecology of Grazing and Browsing Ruminants. In The Ecology of Browsing and Grazing; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2008; pp. 89–116. ISBN 978-3-540-72421-6. [Google Scholar]
- Hackmann, T.J.; Spain, J.N. Invited review: Ruminant ecology and evolution: Perspectives useful to ruminant livestock research and production. J. Dairy Sci. 2010, 93, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Larter, N.C.; Nagy, J.A. Seasonal and annual variability in the quality of important forage plants on Banks Island, Canadian High Arctic. Appl. Veg. Sci. 2001, 4, 115–128. [Google Scholar] [CrossRef]
- Barboza, P.S.; Reynolds, P.E. Monitoring nutrition of a large grazer: Muskoxen on the Arctic Wildlife Refuge. Int. Congr. Ser. 2004, 1275, 327–333. [Google Scholar] [CrossRef]
- Ihl, C.; Klein, D.R. Habitat and Diet Selection by Muskoxen and Reindeer in Western Alaska. J. Wildl. Manag. 2001, 65, 964–972. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Svoboda, J.; Henry, G.; Raillard, M.; Reuss, R. Tundra grazing systems and climatic change. In Arctic Ecosystems in a Changing Climate: An Ecophysiological Perspective; Academic Press: San Diego, CA, USA, 1992; pp. 391–412. [Google Scholar]
- Kazmin, V.D.; Kholod, S.S.; Rozenfeld, S.B.; Abaturov, B.D. Current state of forage resources and feeding of reindeer (Rangifer tarandus) and musk oxen (Ovibos moschatus) in the arctic tundras of Wrangel Island. Biol. Bull. 2011, 38, 747–753. [Google Scholar] [CrossRef]
- Larter, N.C.; Nagy, J.A. Peary caribou, muskoxen and Banks Island forage: Assessing seasonal diet similarities. Rangifer 1997, 17, 9–16. [Google Scholar] [CrossRef]
- Gustine, D.D.; Barboza, P.S.; Lawler, J.P.; Arthur, S.M.; Shults, B.S.; Persons, K.; Adams, L.G. Characteristics of foraging sites and protein status in wintering muskoxen: Insights from isotopes of nitrogen. Oikos 2011, 120, 1546–1556. [Google Scholar] [CrossRef]
- Lawler, J.P.; White, R.G. Seasonal changes in metabolic rates in muskoxen following twenty-four hours of starvation. Rangifer 1997, 17, 135–138. [Google Scholar] [CrossRef]
- Munn, A.J.; Barboza, P.S. Could a big gut be too costly for muskoxen (Ovibos moschatus) in their first winter? Zoology 2008, 111, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Adamczewski, J.Z.; Flood, P.F.; Chaplin, R.K.; Schaefer, J.A. Seasonal variation in intake and digestion of a high-roughage diet by muskoxen. Can. J. Anim. Sci. 1994, 74, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Barboza, P.S.; Peltier, T.C.; Forster, R.J. Ruminal fermentation and fill change with season in an arctic grazer: Responses to hyperphagia and hypophagia in muskoxen (Ovibos moschatus). Physiol. Biochem. Zool. 2006, 79, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Peltier, T.C.; Barboza, P.S.; Blake, J.E. Seasonal hyperphagia does not reduce digestive efficiency in an Arctic grazer. Physiol. Biochem. Zool. 2003, 76, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S. Physical constraints on voluntary intake of forages by ruminants. J. Anim. Sci. 1996, 74, 3063–3075. [Google Scholar] [CrossRef] [PubMed]
- Crater, A.R.; Barboza, P.S.; Forster, R.J. Regulation of rumen fermentation during seasonal fluctuations in food intake of muskoxen. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Lechner, I.; Barboza, P.; Collins, W.; Tervoort, T.A.; Südekum, K.-H.; Codron, D.; Hummel, J. The effect of size and density on the mean retention time of particles in the reticulorumen of cattle (Bos primigenius f. taurus), muskoxen (Ovibos moschatus) and moose (Alces alces). Br. J. Nutr. 2011, 105, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Lauper, M.; Lechner, I.; Barboza, P.; Collins, W.; Hummel, J.; Codron, D.; Clauss, M. Rumination of different-sized particles in muskoxen (Ovibos moschatus) and moose (Alces alces) on grass and browse diets, and implications for rumination in different ruminant feeding types. Mamm. Biol.-Z. Säugetierkd. 2013, 78, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Lechner, I.; Barboza, P.; Collins, W.; Fritz, J.; Günther, D.; Hattendorf, B.; Hummel, J.; Südekum, K.-H.; Clauss, M. Differential passage of fluids and different-sized particles in fistulated oxen (Bos primigenius f. taurus), muskoxen (Ovibos moschatus), reindeer (Rangifer tarandus) and moose (Alces alces): Rumen particle size discrimination is independent from contents stratification. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knott, K.K.; Barboza, P.S.; Bowyer, R.T.; Blake, J.E. Nutritional development of feeding strategies in arctic ruminants: Digestive morphometry of reindeer, Rangifer tarandus, and muskoxen, Ovibos moschatus. Zoology 2004, 107, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Prothero, D.R.; Schoch, R.M. Horns, Tusks, and Flippers: The Evolution of Hoofed Mammals; Johns Hopkins University Press: Baltimore, MD, USA, 2002. [Google Scholar]
- Ropiquet, A.; Hassanin, A. Molecular phylogeny of caprines (Bovidae, Antilopinae): The question of their origin and diversification during the Miocene. J. Zool. Syst. Evol. Res. 2005, 43, 49–60. [Google Scholar] [CrossRef]
- Thulin, C.-G.; Englund, L.; Ericsson, G.; Spong, G. The impact of founder events and introductions on genetic variation in the muskox Ovibos moschatus in Sweden. Acta Theriol. 2011, 56, 305–314. [Google Scholar] [CrossRef]
- Holm, L.-E.; Forchhammer, M.C.; Boomsma, J.J. Low genetic variation in muskoxen (Ovibos moschatus) from western Greenland using microsatellites. Mol. Ecol. 1999, 8, 675–679. [Google Scholar] [CrossRef] [PubMed]
- MacPhee, R.D.; Tikhonov, A.N.; Mol, D.; Greenwood, A.D. Late Quaternary loss of genetic diversity in muskox (Ovibos). BMC Evol. Biol. 2005, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Wang, P.; O’Toole, N.; Barboza, P.S.; Ungerfeld, E.; Leigh, M.B.; Selinger, L.B.; Butler, G.; Tsang, A.; McAllister, T.A.; et al. Snapshot of the Eukaryotic Gene Expression in Muskoxen Rumen—A Metatranscriptomic Approach. PLoS ONE 2011, 6, e20521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen-Ranberg, E.U.; Barnes, C.J.; Rasmussen, L.; Salgado-Flores, A.; Grøndahl, C.; Mosbacher, J.B.; Hansen, A.J.; Sundset, M.A.; Schmidt, N.M.; Sonne, C. A Comparative Study on the Faecal Bacterial Community and Potential Zoonotic Bacteria of Muskoxen (Ovibos moschatus) in Northeast Greenland, Northwest Greenland and Norway. Microorganisms 2018, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Flores, A.; Bockwoldt, M.; Hagen, L.H.; Pope, P.B.; Sundset, M.A. First insight into the faecal microbiota of the high Arctic muskoxen (Ovibos moschatus). Microb. Genomics 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Ørskov, E.R.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Kong, Y.; Teather, R.; Forster, R. Composition, spatial distribution, and diversity of the bacterial communities in the rumen of cows fed different forages. FEMS Microbiol. Ecol. 2010, 74, 612–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, S.; Sun, Y.; Wolcott, R.; Domingo, A.; Carroll, J. Bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP) for microbiome studies: Bacterial diversity in the ileum of newly weaned Salmonella-infected pigs. Foodborne Pathog. Dis. 2008, 5, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- SAS. JMP Release 8.0.2. Available online: http://www.jmp.com/support/notes/41/addl/fusion_41004_6_releasenotes8_0_2.pdf (accessed on 1 June 2018).
- Gustine, D.D.; Barboza, P.S.; Adams, L.G.; Griffith, D.B.; Cameron, R.D.; Whitten, K.R. Advancing the match-mismatch framework for large herbivores in the arctic: Evaluating the evidence for a trophic mismatch in caribou. PLoS ONE 2017, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Barboza, P.S.; Van Someren, L.L.; Gustine, D.D.; Bret-Harte, M.S. The nitrogen window for arctic herbivores: Plant phenology and protein gain of migratory caribou (Rangifer tarandus). Ecosphere, 2018, 9. [Google Scholar] [CrossRef]
- Adamczewski, J.; Chaplin, R.; Schaefer, J.; Flood, P.F. Intake, digestibility and passage rate of a supplemented hay diet in captive muskoxen. Rangifer 1993, 13, 57–60. [Google Scholar] [CrossRef]
- Leventini, M.W.; Hunt, C.W.; Roffler, R.E.; Casebolt, D.G. Effect of dietary level of barley-based supplements and ruminal buffer on digestion and growth by beef cattle. J. Anim. Sci. 1990, 68, 4334–4344. [Google Scholar] [CrossRef] [PubMed]
- Messman, M.A.; Weiss, W.P.; Erickson, D.O. Effects of nitrogen fertilization and maturity of bromegrass on in situ ruminal digestion kinetics of fiber. J. Anim. Sci. 1991, 69, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Carey, D.A.; Caton, J.S.; Biondini, M. Influence of energy source on forage intake, digestibility, in situ forage degradation, and ruminal fermentation in beef steers fed medium-quality brome hay. J. Anim. Sci. 1993, 71, 2260–2269. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, P.; Vanhatalo, A. Ruminal and total plant cell-wall digestibility estimated by a combined in situ method utilizing mathematical models. Br. J. Nutr. 1997, 78, 583–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leupp, J.L.; Lardy, G.P.; Karges, K.K.; Gibson, M.L.; Caton, J.S. Effects of increasing levels of corn distillers dried grains with solubles to steers offered moderate-quality forage. J. Anim. Sci. 2009, 87, 4064–4072. [Google Scholar] [CrossRef] [PubMed]
- Adamczewski, J.; Gunn, A.; Laarveld, B.; Flood, P.F. Seasonal changes in weight, condition and nutrition of free-ranging and captive muskox females. Rangifer 1992, 12, 179–183. [Google Scholar] [CrossRef]
- Russell, J.B.; Muck, R.E.; Weimer, P.J. Quantitative analysis of cellulose degradation and growth of cellulolytic bacteria in the rumen. FEMS Microbiol. Ecol. 2009, 67, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. Seoul Korea 2018, 56, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.; Flint, H.; Bryant, M.P. The rumen bacteria. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 10–72. [Google Scholar]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Forster, R.J.; Koike, S.; Armstrong, J.A.; Teather, R.M.; Kobayashi, Y. Development of fluorescent 16S rRNA probes for studies of rumen ecology. Reprod. Nutr. Dev. 2000, 40, 175. [Google Scholar]
- Lau, S.K.P.; McNabb, A.; Woo, G.K.S.; Hoang, L.; Fung, A.M.Y.; Chung, L.M.W.; Woo, P.C.Y.; Yuen, K.-Y. Catabacter hongkongensis gen. nov., sp. nov., isolated from blood cultures of patients from Hong Kong and Canada. J. Clin. Microbiol. 2007, 45, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Benno, Y. Catenibacterium mitsuokai gen. nov., sp. nov., a gram-positive anaerobic bacterium isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 4, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Barcenilla, A.; Stewart, C.S.; Pryde, S.E.; Flint, H.J. Acetate Utilization and Butyryl Coenzyme A (CoA): Acetate-CoA Transferase in Butyrate-Producing Bacteria from the Human Large Intestine. Appl. Environ. Microbiol. 2002, 68, 5186–5190. [Google Scholar] [CrossRef] [PubMed]
- Krumholz, L.R.; Bryant, M.P.; Brulla, W.J.; Vicini, J.L.; Clark, J.H.; Stahl, D.A. Proposal of Quinella ovalis gen. nov., sp. nov., Based on Phylogenetic Analysis. Int. J. Syst. Evol. Microbiol. 1993, 43, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Kopečný, J.; Zorec, M.; Mrázek, J.; Kobayashi, Y.; Marinšek-Logar, R. Butyrivibrio hungatei sp. nov. and Pseudobutyrivibrio xylanivorans sp. nov., butyrate-producing bacteria from the rumen. Int. J. Syst. Evol. Microbiol. 2003, 53, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Aminov, R.I.; Scott, K.P.; Louis, P.; Stanton, T.B.; Flint, H.J. Proposal of Roseburia faecis sp. nov., Roseburia hominis sp. nov. and Roseburia inulinivorans sp. nov., based on isolates from human faeces. Int. J. Syst. Evol. Microbiol. 2006, 56, 2437–2441. [Google Scholar] [CrossRef] [PubMed]
- Mirande, C.; Kadlecikova, E.; Matulova, M.; Capek, P.; Bernalier-Donadille, A.; Forano, E.; Béra-Maillet, C. Dietary fibre degradation and fermentation by two xylanolytic bacteria Bacteroides xylanisolvens XB1AT and Roseburia intestinalis XB6B4 from the human intestine. J. Appl. Microbiol. 2010, 109, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Orpin, C.G.; Joblin, K.N. The rumen anaerobic fungi. In The Rumen Microbial Ecosystem; Springer: Dordrecht, The Netherlands, 1997; pp. 140–195. ISBN 978-94-010-7149-9. [Google Scholar]
- Vaidya, J.D.; van den Bogert, B.; Edwards, J.E.; Boekhorst, J.; van Gastelen, S.; Saccenti, E.; Plugge, C.M.; Smidt, H. The Effect of DNA Extraction Methods on Observed Microbial Communities from Fibrous and Liquid Rumen Fractions of Dairy Cows. Front. Microbiol. 2018, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- De Mulder, T.; Goossens, K.; Peiren, N.; Vandaele, L.; Haegeman, A.; De Tender, C.; Ruttink, T.; de Wiele, T.V.; De Campeneere, S. Exploring the methanogen and bacterial communities of rumen environments: Solid adherent, fluid and epimural. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Mosbacher, J.B.; Michelsen, A.; Stelvig, M.; Hendrichsen, D.K.; Schmidt, N.M. Show Me Your Rump Hair and I Will Tell You What You Ate—The Dietary History of Muskoxen (Ovibos moschatus) Revealed by Sequential Stable Isotope Analysis of Guard Hairs. PLoS ONE 2016, 11, e0152874. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Feed | DM (kg kg−1) | NDF (kg kg DM−1) | ADF (kg kg DM−1) | Cellulose (kg kg DM−1) | Hemicellulose (kg kg DM−1) | Lignin (kg kg DM−1) | N (kg kg DM−1) |
|---|---|---|---|---|---|---|---|
| Hay | 0.875 | 0.700 | 0.379 | 0.318 | 0.322 | 0.061 | 0.0756 |
| Straw | 0.878 | 0.846 | 0.536 | 0.435 | 0.310 | 0.10 | 0.0359 |
| SEM | 1.82 × 10−3 | 0.0090 | 0.00649 | 0.022 | 0.0077 | 0.027 | 4.31 × 10−3 |
| p = | 0.49 | <0.001 | <0.001 | <0.001 | 0.090 | 0.12 | 0.012 |
| Sedge | 0.645–0.717 | 0.284–0.370 | 0.214–0.291 | 0.347–0.395 | 0.0621–0.0797 | 0.0638–0.156 |
| Phylum | Season | Diet | Season p = | Diet p = | Season × Diet p = | SEM | ||
|---|---|---|---|---|---|---|---|---|
| Spring | Fall | Hay | Straw | |||||
| Actinobacteria | 4.98 × 10−3 | 4.42 × 10−3 | 4.94 × 10−3 | 4.46 × 10−3 | 0.40 | 0.47 | 0.25 | 3.3 × 10−4 |
| Bacteroidetes | 0.335 | 0.346 | 0.340 | 0.341 | 0.37 | 0.93 | 0.35 | 5.53 × 10−3 |
| Fibrobacteres | 2.81 × 10−2 | 3.42 × 10−2 | 2.46 × 10−2 | 3.77 × 10−2 | 0.32 | 0.036 | 0.35 | 2.96 × 10−3 |
| TM7 | 5.65 × 10−3 | 3.85 × 10−3 | 5.88 × 10−3 | 3.62 × 10−3 | 0.005 | 0.001 | 0.86 | 3.1 × 10−4 |
| Firmicutes | 0.577 | 0.566 | 0.577 | 0.566 | 0.39 | 0.30 | 0.10 | 5.48 × 10−3 |
| Lentisphaerae | 6.13 × 10−3 | 4.73 × 10−3 | 5.50 × 10−3 | 5.36 × 10−3 | 0.14 | 0.88 | 0.93 | 4.8 × 10−4 |
| Planctomycetes | 1.18 × 10−3 | 3.94 × 10−4 | 4.95 × 10−4 | 1.08 × 10−3 | 0.020 | 0.08 | 0.16 | 1.6 × 10−4 |
| Proteobacteria | 1.44 × 10−2 | 1.63 × 10−2 | 1.63 × 10−2 | 1.44 × 10−2 | 0.33 | 0.29 | 0.91 | 8.8 × 10−4 |
| Spirochaetes | 2.13 × 10−2 | 1.55 × 10−2 | 1.70 × 10−2 | 1.98 × 10−2 | 0.016 | 0.21 | 0.75 | 1.10 × 10−3 |
| Synergistetes | 8.1 × 10−4 | 1.12 × 10−3 | 1.10 × 10−3 | 8.27 × 10−4 | 0.41 | 0.43 | 0.48 | 1.7 × 10−4 |
| Clade | Hay Substrate | Straw Substrate | ||
|---|---|---|---|---|
| Variation in Response Explained (%) | Latent Vectors | Variation in Response Explained (%) | Latent Vectors | |
| Phylum | 14.8 | 2 | 14.9 | 1 |
| Class | 21.4 | 1 | 22.5 | 2 |
| Order | 20.7 | 1 | 25.1 | 2 |
| Family | 34.1 | 1 | 32.2 | 3 |
| Genus | 39.3 | 1 | 36.9 | 2 |
| Species | 35.1 | 1 | 32.7 | 3 |
| NDF Digestion Rate | NDF Digestion at 120 h | Cellulose Digestion Rate | Cellulose Digestion at 120 h | Hemicellulose Digestion Rate | Hemicellulose Digestion at 120 h | |
|---|---|---|---|---|---|---|
| %response explained | 49.8 | 69.5 | 47.7 | 57.9 | 49.8 | 48.4 |
| Number of important genera (variable importance score > 0.8) | 39 | 36 | 42 | 43 | 41 | 37 |
| Positive Associations (≥95% Percentile Variable Importance Score) | ||||||
| Catenibacterium | Catenibacterium | Clostridium | Papillibacter | Catabacter | Clostridium | |
| Catabacter | Clostridium | Catenibacterium | RC9 gut group | Aquiflexum | Barnesiella | |
| Aquiflexum | Papillibacter | Catabacter | Desulfonatrovibrio | Hydrogenoanaerobacterium | Catenibacterium | |
| Clostridium | Catabacter | Ruminobacter | Catenibacterium | Papillibacter | Papillibacter | |
| Negative Associations (≥95% Percentile Variable Importance Score) | ||||||
| Thalassospira | Coprococcus | Prevotella | Thalassospira | Kiloniella | Kiloniella | |
| Prevotella | Ruminococcus | Heliobacillus | Prevotella | Prevotella | Quinella | |
| Roseburia | Pseudobutyrivibrio | Roseburia | Bacteroides | Roseburia | Roseburia | |
| Coprococcus | Roseburia | Thalassospira | Roseburia | Coprococcus | Pseudobutyrivibrio | |
| NDF Digestion Rate | NDF Digestion at 120 h | Cellulose Digestion Rate | Cellulose Digestion at 120 h | Hemicellulose Digestion Rate | Hemicellulose Digestion at 120 h | |
|---|---|---|---|---|---|---|
| %response explained | 59.6 | 72.4 | 49.9 | 56.3 | 56.7 | 56.5 |
| Number of important genera (variable importance > 0.8) | 41 | 37 | 40 | 39 | 42 | 38 |
| Positive Associations (≥95% Percentile Variable Importance Score) | ||||||
| Ruminobacter | Catabacter | Catabacter | Papillibacter | Ruminobacter | Catabacter | |
| Treponema | Catenibacterium | Fibrobacter | RC9 gut group | Clostridium | Barnesiella | |
| Clostridium | Papillibacter | Ethanoligenens | Catenibacterium | Treponema | Clostridium | |
| Catabacter | Clostridium | Anaerofustis | Aquiflexum | Dolosigranulum | Catenibacterium | |
| Negative Associations (≥95% Percentile Variable Importance Score) | ||||||
| Quinella | Quinella | Ruminococcus | Quinella | Geosporobacter | Kiloniella | |
| Pelospora | Pseudobutyrivibrio | Coprococcus | Prevotella | Proxilibacter | Pseudobutyrivibrio | |
| Ruminococcus | Coprococcus | Pelospora | Thalassospira | Desulfonatrovibrio | Coprococcus | |
| Oxobacter | Roseburia | Zhouia | Roseburia | Anaerophaga | Quinella | |
| Reference | Animal | Substrate | ADF (kg kg DM−1) | NDF Digestion Rate (h−1) |
|---|---|---|---|---|
| Leventini et al. [45] | Cattle | Brome/orchard grass hay | 0.39 | 0.029 |
| Messman et al. [46] | Cattle | Brome grass | 0.39 | 0.065 |
| Carey et al. [47] | Cattle | Brome hay | 0.42 | 0.032 |
| Huhtanen & Vanhatalo [48] | Cattle | Dried timothy | 0.37 | 0.035 |
| Leupp et al. [49] | Cattle | Brome hay | 0.38 | 0.037 |
| P. S. Barboza (pers. com.) | Musk oxen | Brome hay (spring) | 0.34 | 0.009 |
| P. S. Barboza (pers. com.) | Musk oxen | Brome hay (fall) | 0.32 | 0.027 |
| P. S. Barboza (pers. com.) | Musk oxen | Brome hay (winter) | 0.35 | 0.016 |
| This study (average spring) | Musk oxen | Brome hay | 0.37 | 0.019 |
| This study (average fall) | Musk oxen | Brome hay | 0.37 | 0.018 |
| Leventini et al. [45] | Cattle | Wheat straw | NR | 0.022 |
| This study (average spring) | Musk oxen | Triticale straw | 0.48 | 0.023 |
| This study (average fall) | Musk oxen | Triticale straw | 0.48 | 0.015 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ungerfeld, E.M.; Leigh, M.B.; Forster, R.J.; Barboza, P.S. Influence of Season and Diet on Fiber Digestion and Bacterial Community Structure in the Rumen of Muskoxen (Ovibos moschatus). Microorganisms 2018, 6, 89. https://doi.org/10.3390/microorganisms6030089
Ungerfeld EM, Leigh MB, Forster RJ, Barboza PS. Influence of Season and Diet on Fiber Digestion and Bacterial Community Structure in the Rumen of Muskoxen (Ovibos moschatus). Microorganisms. 2018; 6(3):89. https://doi.org/10.3390/microorganisms6030089
Chicago/Turabian StyleUngerfeld, Emilio M., Mary Beth Leigh, Robert J. Forster, and Perry S. Barboza. 2018. "Influence of Season and Diet on Fiber Digestion and Bacterial Community Structure in the Rumen of Muskoxen (Ovibos moschatus)" Microorganisms 6, no. 3: 89. https://doi.org/10.3390/microorganisms6030089