Long-Term Biogas Production from Glycolate by Diverse and Highly Dynamic Communities


 ,
,
Abstract
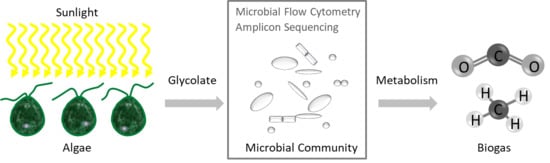
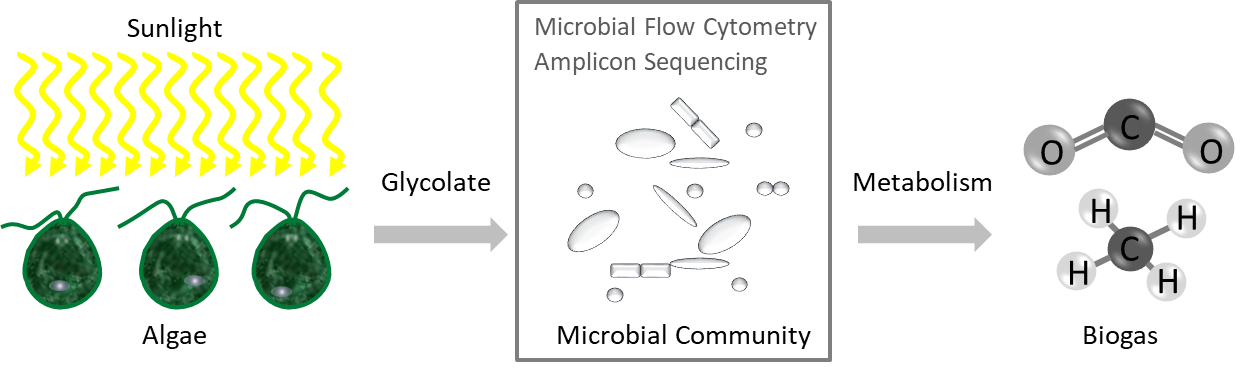
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Digester Configuration and Operational Parameters
2.2. Analytical Methods
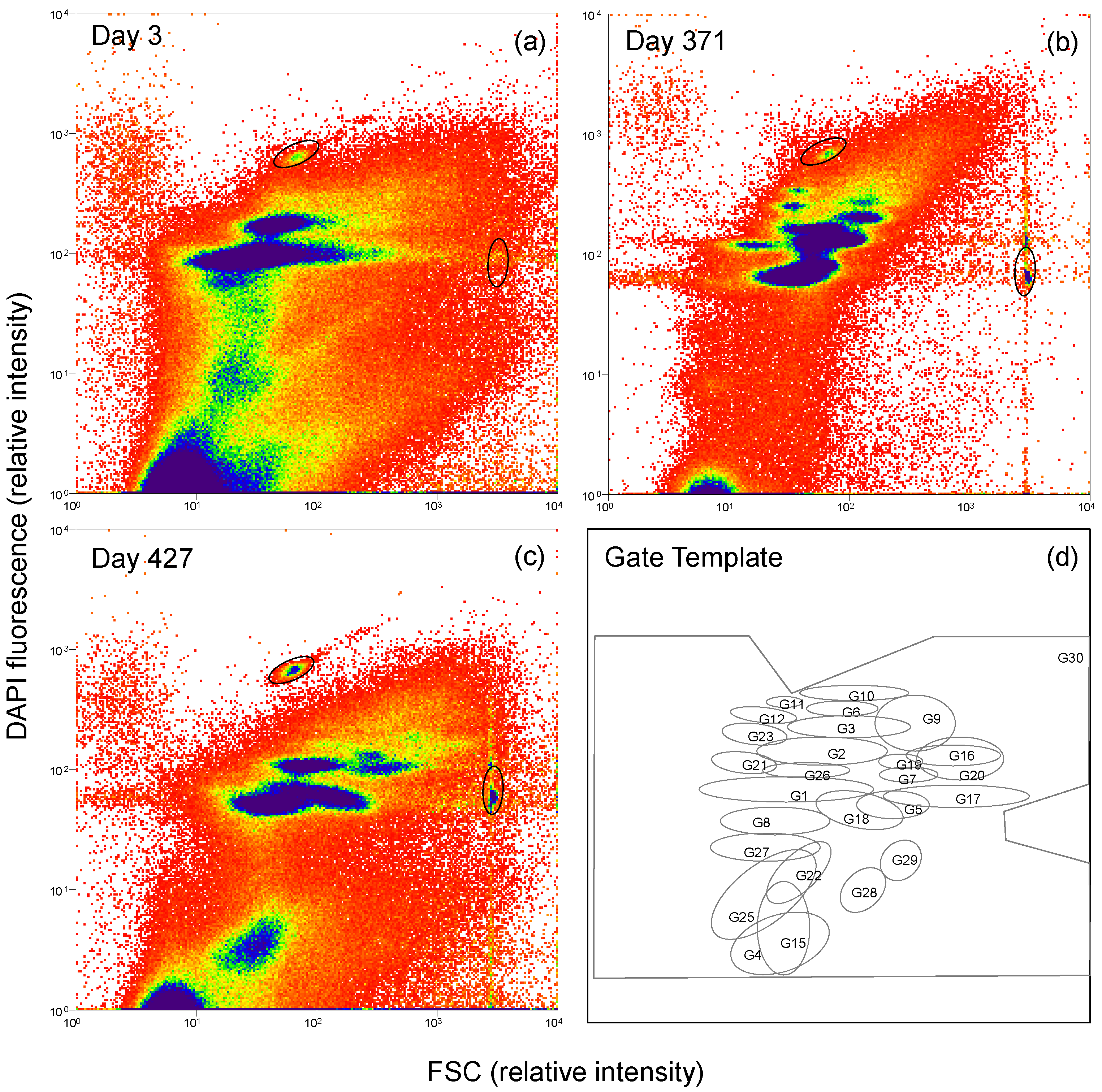
2.3. Flow Cytometry and Cell Sorting
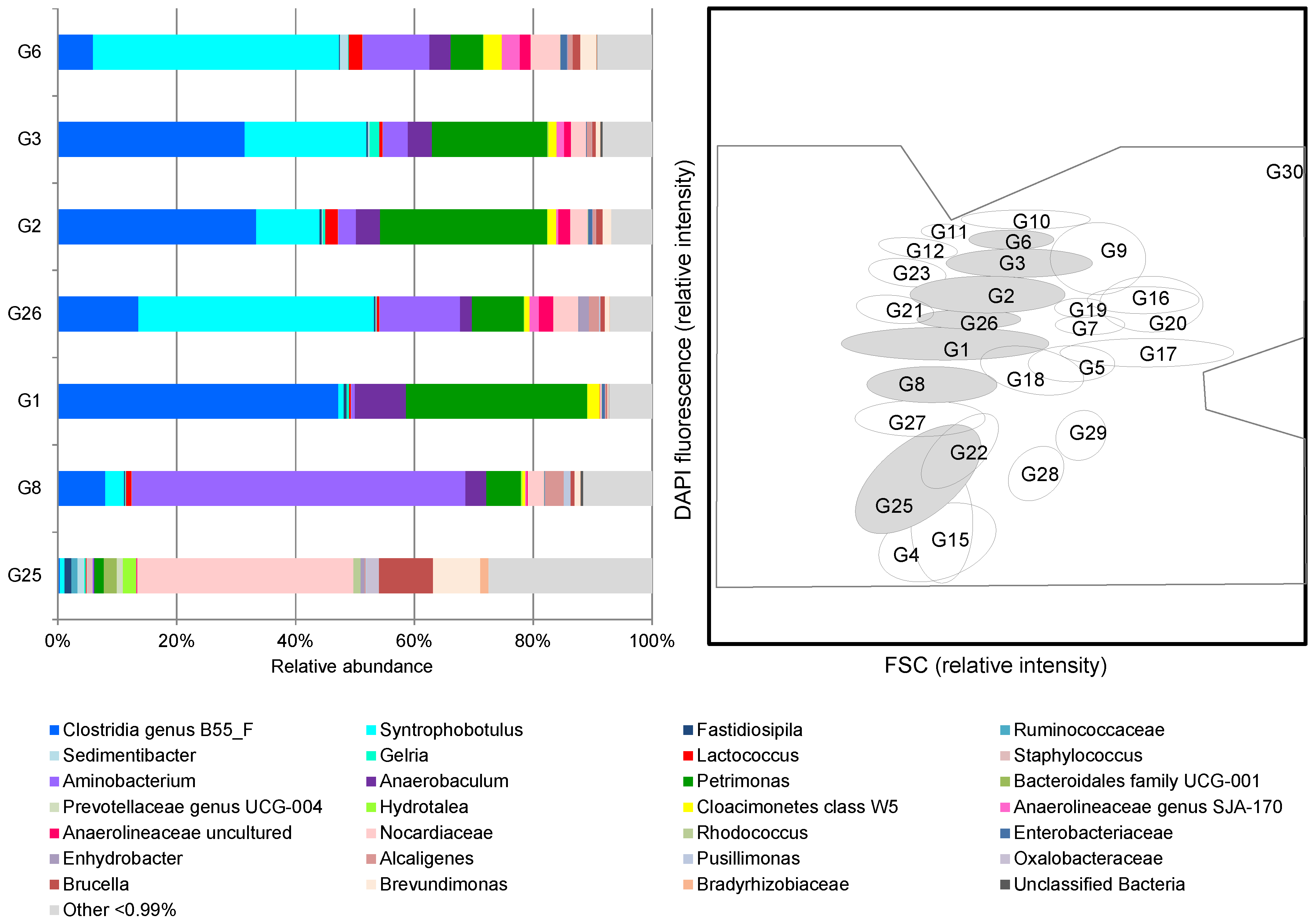
2.4. Microbial Community Analysis by Amplicon Sequencing
2.5. Biostatistics
3. Results
3.1. Glycolate as Sole Carbon Source for Biogas Production
3.2. Microbial Community Dynamics on Glycolate as Mono-Substrate
3.3. Functional Relationships between Biotic and Abiotic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McKendry, P. Energy production from biomass (part 1): Overview of biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef]
- Wilhelm, C.; Weinberg, J.; Kaltschmitt, M. Conversion steps in bioenergy production—Analysis of the energy flow from photon to biofuel. Biofuels 2014, 5, 385–404. [Google Scholar] [CrossRef]
- Gronenberg, L.S.; Marcheschi, R.J.; Liao, J.C. Next generation biofuel engineering in prokaryotes. Curr. Opin. Chem. Biol. 2013, 17, 462–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, E.; Ross, I.L.; King, Z.; Mussgnug, J.H.; Kruse, O.; Posten, C.; Borowitzka, M.A.; Hankamer, B. An economic and technical evaluation of microalgal biofuels. Nat. Biotechnol. 2010, 28, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, J.; Ludley, K.E.; Rowe, R.; Taylor, G.; Howard, D.C. Sources of variability in greenhouse gas and energy balances for biofuel production: A systematic review. GCB Bioenergy 2010, 2, 99–112. [Google Scholar] [CrossRef]
- Wilhelm, C.; Posten, C.; Räbiger, N. Method and Facility for Producing Methane in a Photobioreactor. European Patent EP2614135 A2, 8 September 2011. [Google Scholar]
- Günther, A.; Jakob, T.; Goss, R.; König, S.; Spindler, D.; Räbiger, N.; John, S.; Heithoff, S.; Fresewinkel, M.; Posten, C.; Wilhelm, C. Methane production from glycolate excreting algae as a new concept in the production of biofuels. Bioresour. Technol. 2012, 121, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Laderer, U.; Schink, B. Fermentative degradation of glycolic acid by defined syntrophic cocultures. Arch. Microbiol. 1991, 156, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, M.; Schink, B. Hydrogen formation from glycolate driven by reversed electron transport in membrane vesicles of a syntrophic glycolate-oxidizing bacterium. Eur. J. Biochem. FEBS 1993, 217, 233–240. [Google Scholar] [CrossRef]
- Ornston, L.N.; Ornston, M.K. Regulation of glyoxylate metabolism in Escherichia coli K-12. J. Bacteriol. 1969, 98, 1098–1108. [Google Scholar] [PubMed]
- Kornberg, H.L.; Sadler, J.R. Microbial oxidation of glycollate via a dicarboxylic acid cycle. Nature 1960, 185, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.W.; Hayashi, J.A. Glycolate metabolism in Escherichia coli. J. Bacteriol. 1962, 83, 679–687. [Google Scholar] [PubMed]
- Kornberg, H.L.; Gotto, A.M. The metabolism of C2 compounds in micro-organisms. 6. Synthesis of cell constituents from glycollate by Pseudomonas sp. Biochem. J. 1961, 78, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Kurz, W.G.W.; LaRue, T.A.G. Metabolism of glycolic acid by Azotobacter chroococcum PRL H62. Can. J. Microbiol. 1973, 19, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, H.L.; Morris, J.G. The utilization of glycolate by Micrococcus denitrificans: The β-hydroxyaspartate pathway. Biochem. J. 1965, 95, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Springer, N.; Ludwig, W.; Schink, B. Phylogenetic positions of Desulfofustis glycolicus gen. nov., sp. nov. and Syntrophobotulus glycolicus gen. nov., sp. nov., two new strict anaerobes growing with glycolic acid. Int. J. Syst. Bacteriol. 1996, 46, 1065–1069. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Inokuma, K.; Nakashimada, Y.; Nishio, N. Degradation of glyoxylate and glycolate with ATP synthesis by a thermophilic anaerobic bacterium, Moorella sp. strain HUC22-1. Appl. Environ. Microbiol. 2008, 74, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Seifritz, C.; Fröstl, J.M.; Drake, H.L.; Daniel, S.L. Glycolate as a metabolic substrate for the acetogen Moorella thermoacetica. FEMS Microbiol. Lett. 1999, 170, 399–405. [Google Scholar] [CrossRef]
- Janssen, P.H.; Hugenholtz, P. Fermentation of glycolate by a pure culture of a strictly anaerobic gram-positive bacterium belonging to the family Lachnospiraceae. Arch. Microbiol. 2003, 179, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Schink, B. Electron transport phosphorylation driven by glyoxylate respiration with hydrogen as electron donor in membrane vesicles of a glyoxylate-fermenting bacterium. Arch. Microbiol. 1995, 163, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigematsu, T.; Tang, Y.; Kawaguchi, H.; Ninomiya, K.; Kijima, J.; Kobayashi, T.; Morimura, S.; Kida, K. Effect of dilution rate on structure of a mesophilic acetate-degrading methanogenic community during continuous cultivation. J. Biosci. Bioeng. 2003, 96, 547–558. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Shigematsu, T.; Morimura, S.; Kida, K. Effect of dilution rate on the microbial structure of a mesophilic butyrate-degrading methanogenic community during continuous cultivation. Appl. Microbiol. Biotechnol. 2007, 75, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, T.; Era, S.; Mizuno, Y.; Ninomiya, K.; Kamegawa, Y.; Morimura, S.; Kida, K. Microbial community of a mesophilic propionate-degrading methanogenic consortium in chemostat cultivation analyzed based on 16S rRNA and acetate kinase genes. Appl. Microbiol. Biotechnol. 2006, 72, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Huang, S.; Seston, S.; Xing, J.; Hickey, R.; Criddle, C.; Tiedje, J. How stable is stable? Function versus community composition. Appl. Environ. Microbiol. 1999, 65, 3697–3704. [Google Scholar] [PubMed]
- Koch, C.; Fetzer, I.; Schmidt, T.; Harms, H.; Müller, S. Monitoring functions in managed microbial systems by cytometric bar coding. Environ. Sci. Technol. 2013, 47, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Burr, M.; Camper, A.K. Genotypic microbial community profiling: A critical technical review. Microb. Ecol. 2007, 54, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Shokralla, S.; Spall, J.L.; Gibson, J.F.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol. Ecol. 2012, 21, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Faust, K.; Raes, J. CoNet app: Inference of biological association networks using Cytoscape [version 2; referees: 2 approved]. F1000Research 2016, 5, 1519. [Google Scholar] [CrossRef] [PubMed]
- flowCHIC. Available online: http://www.bioconductor.org/packages/release/bioc/html/flowCHIC.html (accessed on 20 June 2017).
- Koch, C.; Harnisch, F.; Schröder, U.; Müller, S. Cytometric fingerprints: Evaluation of new tools for analyzing microbial community dynamics. Front. Microbiol. 2014, 5, 273. [Google Scholar] [CrossRef] [PubMed]
- Pous, N.; Koch, C.; Vilà-Rovira, A.; Balaguer, M.D.; Colprim, J.; Mühlenberg, J.; Müller, S.; Harnisch, F.; Puig, S. Monitoring and engineering reactor microbiomes of denitrifying bioelectrochemical systems. RSC Adv. 2015, 5, 68326–68333. [Google Scholar] [CrossRef]
- Günther, S.; Faust, K.; Schumann, J.; Harms, H.; Raes, J.; Müller, S. Species-sorting and mass-transfer paradigms control managed natural metacommunities. Environ. Microbiol. 2016, 18, 4862–4877. [Google Scholar] [CrossRef] [PubMed]
- Strach, K. Determination of the VOA value (according to Kapp) and of the VOA/TIC value (in accordance with FAL). In Collection of Methods for Biogas; Liebetrau, J., Pfeiffer, D., Thrän, D., Eds.; German Biomass Research Centre: Leipzig, Germany, 2016; pp. 32–34. [Google Scholar]
- Koch, C.; Günther, S.; Desta, A.F.; Hübschmann, T.; Müller, S. Cytometric fingerprinting for analyzing microbial intracommunity structure variation and identifying subcommunity function. Nat. Protoc. 2013, 8, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Luton, P.E.; Wayne, J.M.; Sharp, R.J.; Riley, P.W. The mcrA gene as an alternative to 16S rRNA in the phylogenetic analysis of methanogen populations in landfill. Microbiology 2002, 148, 3521–3530. [Google Scholar] [CrossRef] [PubMed]
- FASTX-Toolkit. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 29 June 2017).
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- McIlroy, S.J.; Saunders, A.M.; Albertsen, M.; Nierychlo, M.; McIlroy, B.; Hansen, A.A.; Karst, S.M.; Nielsen, J.L.; Nielsen, P.H. MiDAS: The field guide to the microbes of activated sludge. Database 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.A.; Chai, B.; Wang, Q.; Sun, Y.; Brown, C.T.; Tiedje, J.M.; Cole, J.R. FunGene: The functional gene pipeline and repository. Front. Microbiol. 2013, 4, 291. [Google Scholar] [CrossRef] [PubMed]
- Günther, S.; Müller, S. Facilitated gate setting by sequential dot plot scanning. Cytom. Part A 2015, 87, 661–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrell, F.E. Hmisc: Harrell Miscellaneous. Available online: http://cran.R-project.Org/web/packages/Hmisc (accessed on 11 June 2018).
- Schultz, J.A.; Beck, C.; Menz, G.; Neuwirth, B.; Ohlwein, C.; Philipp, A. Sensitivity of proxies on non-linear interactions in the climate system. Sci. Rep. 2015, 21, 18560. [Google Scholar] [CrossRef] [PubMed]
- Cabrieto, J.; Tuerlinckx, F.; Kuppens, P.; Hunyadi, B.; Ceulemans, E. Testing for the presence of correlation changes in a multivariate time series: A permutation based approach. Sci. Rep. 2018, 8, 769. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar]
- Liebetrau, J.; Sträuber, H.; Kretzschmar, J.; Denysenko, V.; Nelles, M. Anaerobic digestion. In Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–19. [Google Scholar] [CrossRef]
- Haller, T.; Buckel, T.; Rétey, J.; Gerlt, J.A. Discovering new enzymes and metabolic pathways: Conversion of succinate to propionate by Escherichia coli. Biochemistry 2000, 39, 4622–4629. [Google Scholar] [CrossRef] [PubMed]
- Stams, A.J.M.; Plugge, C.M. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 2009, 7, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Vanwonterghem, I.; Evan, P.N.; Parks, D.H.; Jensen, P.D.; Woodcroft, P.J.; Hugenholtz, P.; Tyson, G.W. Methylotrophic methanogenesis discovered in the archaeal phylum Verstraetearchaeota. Nat. Microbiol. 2016, 1, 16170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowski, A.; Tindall, B.J.; Bardin, V.; Blanchet, D.; Jeanthon, C. Petrimonas sulfuriphila gen. nov., sp. nov., a mesophilic fermentative bacterium isolated from a biodegraded oil reservoir. Int. J. Syst. Evol. Microbiol. 2005, 55, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Hahnke, S.; Langer, T.; Koeck, D.E.; Klocke, M. Description of Proteiniphilum saccharofermentans sp. nov., Petrimonas mucosa sp. nov. and Fermentimonas caenicola gen. nov., sp. nov., isolated from mesophilic laboratory-scale biogas reactors, and emended description of the genus Proteiniphilum. Int. J. Syst. Evol. Microbiol. 2016, 66, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Baena, S.; Fardeau, M.L.; Labat, M.; Ollivier, B.; Thomas, P.; Garcia, J.L.; Patel, B.K. Aminobacterium colombiense gen. nov. sp. nov., an amino acid-degrading anaerobe isolated from anaerobic sludge. Anaerobe 1998, 4, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Baena, S.; Fardeau, M.L.; Labat, M.; Ollivier, B.; Garcia, J.L.; Patel, B.K. Aminobacterium mobile sp. nov., a new anaerobic amino-acid-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2000, 50, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, O.; Ben Hania, W.; Postec, A.; Bouallagui, H.; Hamdi, M.; Bonin, P.; Ollivier, B.; Fardeau, M.L. Aminobacterium thunnarium sp. nov., a mesophilic, amino acid-degrading bacterium isolated from an anaerobic sludge digester, pertaining to the phylum Synergistetes. Int. J. Syst. Evol. Microbiol. 2015, 65, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Rees, G.N.; Patel, B.K.C.; Grassia, G.S.; Sheehy, A.J. Anaerobaculum thermoterrenum gen. nov., sp. nov., a novel, thermophilic bacterium which ferments citrate. Int. J. Syst. Bacteriol. 1997, 47, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Menes, R.J.; Muxi, L. Anaerobaculum mobile sp. nov., a novel anaerobic, moderately thermophilic, peptide-fermenting bacterium that uses crotonate as an electron acceptor, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2002, 52, 157–164. [Google Scholar] [PubMed]
- Maune, M.W.; Tanner, R.S. Description of Anaerobaculum hydrogeniformans sp. nov., an anaerobe that produces hydrogen from glucose, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2012, 62, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, E.; Kreimeyer, A.; Bocs, S.; Rouy, Z.; Gyapay, G.; Chouari, R.; Rivière, D.; Ganesan, A.; Daegelen, P.; Sghir, A.; et al. “Candidatus Cloacamonas acidaminovorans”: Genome sequence reconstruction provides a first glimpse of a new bacterial division. J. Bacteriol. 2008, 190, 2572–2579. [Google Scholar] [CrossRef] [PubMed]
- Goux, X.; Calusinska, M.; Lemaigre, S.; Marynowska, M.; Klocke, M.; Udelhoven, T.; Benizri, E.; Delfosse, P. Microbial community dynamics in replicate anaerobic digesters exposed sequentially to increasing organic loading rate, acidosis, and process recovery. Biotechnol. Biofuels 2015, 8, 122. [Google Scholar] [CrossRef] [PubMed]
- Nettmann, E.; Bergmann, I.; Mundt, K.; Linke, B.; Klocke, M. Archaea diversity within a commercial biogas plant utilizing herbal biomass determined by 16S rDNA and mcrA analysis. J. Appl. Microbiol. 2008, 105. [Google Scholar] [CrossRef] [PubMed]
- Gunderson, L.H. Ecological resilience—In theory and application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Delvigne, F.; Baert, J.; Sassi, H.; Fickers, P.; Grünberger, A.; Dusny, C. Taking control over microbial populations: Current approaches for exploiting biological noise in bioprocesses. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Günther, S.; Becker, D.; Hübschmann, T.; Reinert, S.; Kleinsteuber, S.; Müller, S.; Wilhelm, C. Long-Term Biogas Production from Glycolate by Diverse and Highly Dynamic Communities. Microorganisms 2018, 6, 103. https://doi.org/10.3390/microorganisms6040103
Günther S, Becker D, Hübschmann T, Reinert S, Kleinsteuber S, Müller S, Wilhelm C. Long-Term Biogas Production from Glycolate by Diverse and Highly Dynamic Communities. Microorganisms. 2018; 6(4):103. https://doi.org/10.3390/microorganisms6040103
Chicago/Turabian StyleGünther, Susanne, Daniela Becker, Thomas Hübschmann, Susann Reinert, Sabine Kleinsteuber, Susann Müller, and Christian Wilhelm. 2018. "Long-Term Biogas Production from Glycolate by Diverse and Highly Dynamic Communities" Microorganisms 6, no. 4: 103. https://doi.org/10.3390/microorganisms6040103