Omics Analysis for Dinoflagellates Biology Research

1
Laboratory of Synthetic Microbiology, School of Chemical Engineering & Technology, Tianjin University, Tianjin 300072, China
2
Frontier Science Center for Synthetic Biology and Key Laboratory of Systems Bioengineering, Ministry of Education of China, Tianjin 300072, China
3
Collaborative Innovation Center of Chemical Science and Engineering, Tianjin 300072, China
4
Center for Biosafety Research and Strategy, Tianjin University, Tianjin 300072, China
*
Authors to whom correspondence should be addressed.
Microorganisms 2019, 7(9), 288; https://doi.org/10.3390/microorganisms7090288
Submission received: 9 July 2019
/
Revised: 20 August 2019
/
Accepted: 21 August 2019
/
Published: 23 August 2019
(This article belongs to the Special Issue Dinoflagellate Biology in the Omics Era)
Abstract
:Dinoflagellates are important primary producers for marine ecosystems and are also responsible for certain essential components in human foods. However, they are also notorious for their ability to form harmful algal blooms, and cause shellfish poisoning. Although much work has been devoted to dinoflagellates in recent decades, our understanding of them at a molecular level is still limited owing to some of their challenging biological properties, such as large genome size, permanently condensed liquid-crystalline chromosomes, and the 10-fold lower ratio of protein to DNA than other eukaryotic species. In recent years, omics technologies, such as genomics, transcriptomics, proteomics, and metabolomics, have been applied to the study of marine dinoflagellates and have uncovered many new physiological and metabolic characteristics of dinoflagellates. In this article, we review recent application of omics technologies in revealing some of the unusual features of dinoflagellate genomes and molecular mechanisms relevant to their biology, including the mechanism of harmful algal bloom formations, toxin biosynthesis, symbiosis, lipid biosynthesis, as well as species identification and evolution. We also discuss the challenges and provide prospective further study directions and applications of dinoflagellates.
1. Introduction
Dinoflagellates are unicellular protists that have two distinctive flagella during their life cycle [1]. The size of dinoflagellates ranges from a few micrometers to two millimeters [2]. The life styles of dinoflagellates include free living, parasitic, or mutualist [2]. So far nearly 4500 species of dinoflagellates have been isolated, and the majority of them (approximate 4000 species) are free living, while only a small percentage of them are found with parasitic or mutualist symbiotic life styles, such as Syndinophyceae [2]. Three major nutrient mechanisms, phototrophy, heterotrophy and mixotrophy, are all found and adopted by various dinoflagellates species, and among them approximately 50% of dinoflagellates are phototrophic [2,3].
Dinoflagellates receive great attention mostly due to their important roles in natural ecosystems, as well as being the producers of key components in human food (e.g., docosahexaenoic acids (DHA) as essential supplements) [4]. First, together with diatoms and coccolithophores, the photosynthetic species of dinoflagellates are among the most prominent primary producers in marine environments and play vital roles in the global carbon cycle [5]. Second, a number of photosynthetic dinoflagellates form a mutualism with other living organisms for maintaining the stability of ecosystems [6]. For example, reef-building corals are maintained based on symbiosis between the coral animal and Symbiodiniaceae, a photosynthetic dinoflagellate. Corals provide shelter and inorganic nutrients for Symbiodiniaceae, and in return, Symbiodiniaceae provides much more than energy metabolites to corals. They translocate antioxidants, signaling compounds, and are vitally important in nutrient cycling [6,7]. Finally, some heterotrophic dinoflagellates are important producers of highly-valued and essential food components. For example, DHA which has various beneficial effects on human health [8,9,10], such as improving cognitive development in infants and inhibiting hypertension, inflammation and certain cancers, cannot be synthesized by human beings, but accumulates in heterotrophic dinoflagellates Crypthecodinium cohnii [4]. The fraction of DHA in C. cohnii can be up to 25% of dry cell weight [11,12]. Therefore, the fermentation of C. cohnii has been considered as a sustainable and alternative way of producing high-quality DHA [4,12,13,14,15]. Meanwhile, dinoflagellates can also result in negative effects to the environment. One notorious phenomenon is “harmful algal blooms (HABs)”, also known as “red tides” [16], which are formed due to a sharp increase in dinoflagellates and bacteria, which can reach up to twenty million cells per liter along the coasts [2]. These blooms are often associated with the production of various secondary metabolites, which can be extremely toxic and can be more potent than some agents used in chemical warfare [17].
Dinoflagellates have additional prominent biological features that are worth of further exploration. For example, some species of dinoflagellates are capable of bioluminescence [18]. The bioluminescence systems consist of luciferase (enzyme), luciferin (substrate), and a protein that binds luciferin [19]. Bioluminescence seems to be separated by discrete particles called ‘‘scintillons’’ within cells [20]. One of the suggested functions is that it reduces the grazing behavior of copepods [21]. Some dinoflagellates have photosensitive organelles called “eyespots”, which consist of lipid droplets wrapped within stacked layers of membranes [22], and allow dinoflagellates to perform phototaxis: i.e., to move relative to the direction and intensity of light. In addition, dinoflagellate plastids originate from either secondary or tertiary endosymbiotic events, making them different from those found in plants or green alga [23]. The most cytologically prominent feature of dinoflagellates is their abnormal genome size and organization [24]. Typical dinoflagellate DNA content is estimated to be 3 to 250 pg·cell−1, equivalent to about 3000–245,000 Mb [25,26], roughly 1- to 80-fold larger than a human haploid genome [27]. Moreover, dinoflagellate nuclear DNA is widely methylated, of which 12–70% of thymine is replaced by 5-hydroxymethyluracil and cytosine is methylated to varying degrees [27,28,29]. In most eukaryotic organisms, these modifications are due to oxidative damage of thymine or 5-methylcytosine, which is quickly repaired by a DNA glycosylase [30]. Furthermore, the chromosomes of most dinoflagellates are permanently condensed without the aid of nucleosomes throughout some or all stages of their life cycle [24,31]. The protein content in the chromatin is usually low, with a ten-times lower protein: DNA ratio than that in other eukaryotes [16]. Dinoflagellates are the only known eukaryotes to apparently lack histone proteins [32]. Dinoflagellates are capable of recruiting other proteins, such as histone-like proteins from bacteria and dinoflagellate/viral nucleoproteins from viruses, as histone substitutes [32,33,34]. Finally, plastid genomes of dinoflagellates appear in the form of plasmid-like minicircles [35], and their mitochondrial genomes contain only three protein-coding genes and lack stop codons [36].
Due to their ecological and economical roles and nutrition contribution, a better understanding of dinoflagellate biology at the molecular level is important. In recent years, omics technologies [37], such as genomics, transcriptomics, proteomics, and metabolomics, have been successfully applied to the systematic analysis of dinoflagellates, in revealing the organization and evolution of dinoflagellates genomes [7,38,39,40,41], and molecular mechanisms related to HAB formation and toxin biosynthesis [42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61], symbiosis [39,40,41,62,63,64,65,66,67,68,69,70,71], and lipid accumulation [11,12,13,15,72,73,74,75,76] (Figure 1). In this review, we summarize recent progress on the omics studies of dinoflagellates biology. We also discuss the challenges and provide prospects for further study.
2. Omics Study on Dinoflagellates
2.1. Genomic Analysis
Genomic analyses help to examine the primary sequence and structural assembly of the complete genome of an organism [77]. Due to the high sequencing cost of traditional methods, referred to as Sanger sequencing, and the large sizes of dinoflagellates genomes, complete sequencing of a large number of nuclear genomes of dinoflagellates has long been out of reach [24]. With the advent and rapid development of next-generation sequencing (NGS), such as 454 sequencing, Solexa technology, the SOLiD platform, the Polonator, and the HeliScope Single Molecule Sequencer technology, the total cost of sequencing is sharply decreased and sequencing throughput is significantly enlarged [78,79,80], which has facilitated rapid, economic sequencing of a number of dinoflagellates species, with improved reliability and accuracy [7,38,39,40,41].
The most frequently used sequencing technologies for dinoflagellates are NGS 454 sequencing and Illumina Hiseq 2000/2500 (Table 1), which use clonal amplification and sequencing by synthesis to allow parallel sequencing [78]. Except for several thousand times higher sequencing throughput and the dramatically increased degree of parallelism, a major advantage of NGS over Sanger sequencing is the cost reduction by over two orders of magnitude [78]. Because the genomes of symbiotic species are among the smallest (1~5 Gbp) compared with other dinoflagellates [81], the first available draft genome of a dinoflagellate is Breviolim minutum (clade B) [38], soon followed by Fugacium kawagutii (clade F) [39], Symbiodinium microadriaticum (clade A) [40], Cladocopium goreaui (clade C) [7], and other Symbiodiniaceae species [41] (Table 1).
Detailed genomic analysis shows that the gene structures of B. minutum and F. kawagutii have several distinctive and divergent characteristics when compared to those of other eukaryotes. For example, analysis of genome sequences of B. minutum [38] showed that: i) the genome contains nearly twice as many gene families as that of its sister group, the apicomplexans; ii) only one-third of putative proteins encoded by the permanently condensed chromosomes of B. minutum have eukaryotic orthologs; iii) the identified genes are enriched in spliceosomal introns that use different recognition nucleotides at 5′ splice site; iv) genes are arranged in one direction throughout the genome, forming a cluster of genes, which was previously known only in trypanosomes, which are evolutionarily distantly related [82]. What’s more, a novel promoter element (motifs TTTT instead of the TATA box used by other eukaryotes) in the F. kawagutii genome, and a microRNA system potentially regulating gene expression in both symbiont and coral were observed [39].
The draft genomes provide an important resource for understanding the characteristics of Symbiodiniaceae biology and genetic differences between Symbiodiniaceae species (Figure 2). Global annotated genome analysis revealed that: i) common metabolic pathways in typical photosynthetic eukaryotes were found in the genomes of F. kawagutii and C. goreaui [7,39]; ii) Symbiodiniaceae-specific gene families, such as genes related to sexual reproduction, cyst formation, and germination and telomere synthesis, were identified in the assembled F. kawagutii and B. minutum genomes [39]; iii) some genes were highly expanded in the Symbiodiniaceae genome, including genes encoding chlorophyll a/b-binding proteins in B. minutum [38] and Symbiodiniaceae cladeA and Symbiodiniaceae cladeC [41], ion channel proteins and DNA repair/recombination proteins in all sequenced Symbiodiniaceae species [7,38,39,40,41,83], calcium channel and calmodulin families in B. minutum [38], heat shock proteins in B. minutum [38] and F. kawagutii [39], antioxidant genes, including the large thioredoxin gene family, superoxide dismutases and ascorbate peroxidases in F. kawagutii [39], as well as genes related to meiosis and response to light stress in S. microadriaticum, C. goreaui, and F. kawagutii [7]; iv) Symbiodiniaceae harbors a wide range of transport proteins [84], which are related to the supply of carbon and nitrogen [40], are responsive to reactive oxygen species and can prevent ultraviolet radiation [83], and all the studied dinoflagellates have more transmembrane transport proteins involved in the exchange of amino acids, lipids, and glycerol than other eukaryotes through the comparative analysis of genomes and transcriptomes [83]. Mycosporine-like amino acids (MAA) as specific ultraviolet radiation blockers protect the photosynthetic machinery of the dinoflagellate [28]. The MAA biosynthetic pathway involves dehydroquinate synthase, O-methyltransferase, an ATP (adenosine triphosphate)-grasp and non-ribosomal peptide synthetase [85]. It was found that the ability to synthesize mycosporine-like amino acids varied significantly among different Symbiodiniaceae clades [7,39,41]. The genome of Symbiodiniaceae (clade A) contains a gene cluster for the biosynthesis of mycosporine-like amino acids, which may be transferred from an endosymbiotic red alga [41]. In the genomes of Symbiodiniaceae (clade C) and F. kawagutii, early research suggested that they have completely lost the gene cluster [39,41]. However, a recent study using known proteins in bacteria, fungi, and cnidarians as queries, allowed the identification of putative genes encoding all five enzymes, including the short-chain dehydrogenase from the S. microadriaticum, C. goreaui, and F. kawagutii genomes [7].
The draft genomes also provide an important resource for understanding the molecular basis of coral-Symbiodiniaceae symbiosis. For example, i) genes related to synthesis and modification of amino acids were discovered in F. kawagutii, C. goreaui, S. microadriaticum, and B. minutum genomes, as many amino acids cannot be synthesized by coral host that must be supplied by Symbiodiniaceae [7,39]; ii) the recognition of Symbiodiniaceae by the hosts was considered to be mediated through the binding of Symbiodiniaceae high-mannose glycans by lectins on the coral cell surface [86], and the genes encoding a glycan biosynthesis pathway have been identified in F. Kawagutii, C. goreaui, S. microadriaticum, and B. minutum genomes [7,39]; iii) the cytosine methyltransferase gene family in C. goreaui, S. microadriaticum, and B. minutum genomes was shown to be expansive [7,39], which may partially explain a high proportion of diverse methylated nucleotides in dinoflagellates species [27]. Therefore, the observed decreases in fluorometrically assayed β-glucoronidase expression over time in Amphidinium and Symbiodiniaceae might suggest transgene silencing by a mechanism involving methylation [87]; iv) the new-found promoter elements and microRNA systems regulating gene expression may be useful as potential regulatory elements for metabolic engineering in dinoflagellates [39].
Besides whole genome sequencing, NGS can also be applied to other aspects of dinoflagellates biology research, such as mapping of structural rearrangements, analysis of DNA methylation, and identification of DNA-protein interactions [78]. Although NGS technologies are powerful, one of their major limitations is the relatively short reads they generated, which may lead to: i) mis-assemblies and gaps, as many repeated sequences that are longer than NGS reads exist in dinoflagellates genomes [88,89]; ii) more challenges to detect and characterize larger structural variations [90]. Another major limitation is that dinoflagellates genomes contain regions of extreme high content of guanine and cytosine, leading to inefficiently amplification by polymerase chain reaction [91]. The last is that a common NGS approach, bisulfite sequencing, can only detect C modification on treatment of DNA by bisulfite, but cannot distinguish 5mC and 5hmC [92]. In contrast, the emerging third-generation sequencing (TGS) technologies (e.g., single-molecule real-time sequencing used by PacBio, nanopore sequencing used by Oxford Nanopore Technologies and synthetic long reads used by Illumina/10X Genomics) can generate very long reads (tens of thousands of bases per nanopore) at the single-molecule level, which allows less sequencing bias and more homogeneous genome coverage [91]. In addition, Oxford Nanopore Technologies can directly detect DNA methylation 5mC and 6mA without amplification by polymerase chain reaction [93,94]. In spite of the major drawback, a high error rate (~15%), TGS technologies have been applied successfully in the analysis of repeated regions and structural variations, haplotype phasing, and transcriptome analysis [91]. In combination of NGS technologies, it is expected that TGS technologies have the potential to solve the sequencing bottleneck of dinoflagellates with large and complex genomes.
2.2. Transcriptomic Analysis of Marine Dinoflagellates
Although the enormous genome size of most dinoflagellates makes it challenging to obtain a whole genome sequence, one promising strategy to understand the function and regulation of both functionally known and unknown genes in an uncharacterized genome can be achieved by examining gene expression by transcriptomics analysis [95,96]. Several common methodologies for transcriptomics analysis used in dinoflagellates include: i) microarrays, a high throughput and relatively inexpensive method, which is based on hybridization but is subject to a number of limitations (e.g., relying upon existing genome sequence, high levels of background noise, and a limited dynamic range of detection of different isoforms and allelic expression) [97,98,99]; ii) expressed sequence tag (EST) sequencing, a method that is based on Sanger sequencing, and is relatively accurate but has many inherent disadvantages, such as low throughput, high cost, and the lack of the ability of quantification [97,100]; iii) massively parallel signature sequencing (MPSS), which is a tag-based high throughput method for sequencing millions of templates cloned on the surface of microbeads, is still subject to the limitations (e.g., high cost, a limited dynamic range of detection of isoforms, and disability of mapping a part of short tags to the reference genome) [97,101]; iv) RNA-Seq (RNA-sequencing), an high throughput approach employing NGS or TGS for direct sequencing of cDNA transcribed from the whole transcriptomes [97]. RNA-Seq shows clear advantages and has good application prospects, compared with other transcriptomic approaches [97]. First, it is not limited to the detection of transcripts that have corresponding genomic sequences, which attracts non-model organisms whose genome sequences have not yet been determined; second, it also has a high resolution to reveal the precise location of transcription boundaries, wide dynamic range to quantify gene expression level, and the ability to distinguish sequence variations, which are especially useful for complex transcriptomes [97]. Finally, RNA-Seq has a relatively low background, high accuracy of quantitative expression level, high technological reproducibility, and a much lower cost [97]. Thus, RNA-Seq technology has been most widely used approach in the transcriptomic analysis of dinoflagellates in recent years. A detailed summary on transcriptomics analysis of dinoflagellates is provided in Table 2.
Through transcriptomics analysis, several important aspects of genome structure and gene expression regulation in dinoflagellates have been discovered: i) transcriptomic analysis had recognized transcripts for all core histones (H2A, H2B, H3, H4) and their variants (H2A.X and H2A.Z) [125,130,131], despite the fact that dinoflagellates had been previously considered to completely lose histones genes [132]; ii) using the EST library, the genomic structures of 47 genes from the dinoflagellate Amphidinium carterae were identified and addressed [133]. The study showed that almost all highly expressed tags exist in large tandem gene arrays with short intergenic spacers, while the second class of genes showed high intron density and significantly lower copy number. A polyadenylation signal was also discovered in genomic copies at the exact polyadenylation site and was conserved between species; iii) the presence of 5′- trans-spliced leader addition in mRNA processing was found in several dinoflagellates using EST data, suggesting that dinoflagellates heavily rely on the posttranscriptional regulation of gene expression [131,133,134,135]; iv) compared to other eukaryotes, a reduced role for the transcriptional regulation of gene expression was discovered. For example, the analysis showed that only about 3% of the genes were significantly changed at transcriptional level in response to a circadian clock [126] and only 4% in response to oxidative stress in Pyrocystis lunula [125].
Transcriptome profiling also allows for the study of the metabolic and physiological response of dinoflagellates responsive to various stresses. As summarized in Table 2, transcriptional changes of dinoflagellates under different growth conditions have been extensively studied. First, for some species of dinoflagellates responsible for forming HABs, studies have been conducted on how environmental conditions conductive to HABs [46,105,109,114,115] affect the expression of toxin related genes and the toxin biosynthesis pathway [47,56,60,108] as well as physiological processes of HABs outbreak [107]. For example, in Alexandrium tamarense, MPSS results showed that transcripts of chlorophyll a/b binding protein, histone family protein, S-adenosylmethionine synthetase, and S-adenosylhomocysteine hydrolase were the highest expressed among all tested conditions that provoke the formation of HABs [46]. Microarray-based comparative transcriptome profiles of dinoflagellate Alexandrium minutum suggested that many genes were more highly expressed in the toxic than in the non-toxic strain, and several genes were even expressed only in the toxic strains [47]. In addition, a variety of metabolic pathways specially related to various N (cyanate, urea, nitrate/nitrite, and ammonium) uptake and assimilation were enriched, which is likely to confer competitive advantages for bloom formation or maintenance in Alexandrium catenella [107]. Using time sequential metatranscriptomic, they analyzed a natural assemblage that evolved from diatom (Skeletonema) dominance to a dinoflagellate (Prorocentrum donghaiense) bloom [136]. The results showed that during the dominant period, a similar series of most active metabolic processes (energy and nutrient acquisition, stress resistance) promoted growth and distinct metabolic pathways were used by diatom and dinoflagellates in their respective dominance, while P. donghaiense possessed more diversified light energy and phosphate acquisition strategy and antimicrobial defense, which might cause them to grow faster than diatoms and form blooms. Second, due to the potential significance in coral bleaching, transcriptional changes of Symbiodiniaceae in response to thermal stress or light have been widely investigated [64,66,67,121,122]. Transcriptomic responses to heat stress in F. kawagutii were determined, and the results showed that 357 genes were differentially expressed under heat stress, and most of them were involved in regulating cell wall modulation and the transport of iron, oxygen, and major nutrients. What’s more, the expression of heat shock proteins was strongly elevated during heat stress, within expectations. The results also showed that the demand for nutrients, iron, and oxygen in F. kawagutii might be higher under heat stress [122]. When grown under autotrophic or mixotrophic conditions, the expression levels of many Symbiodiniaceae SSB01 genes seemed to be significantly affected by light, such as a cryptochrome 2 gene (declined in high light), regulators of Chromatin Condensation (RCC1) (declined in the dark), a light harvesting AcpPC protein (increased in high light autotrophic conditions), and several cell adhesion proteins (rapidly declined when the culture was moved from low light to darkness) [66]. The increased transcript level of AcpPC gene suggests that it is involved in photo-protection and the dissipation of excess absorbed light energy. The decreased cell adhesion protein level is related to the significant change in cell surface morphology, which may reflect the complexity of the extracellular matrix [66]. These results provided valuable insights into the molecular basis of thermal or light resistance of dinoflagellates and coral bleaching. Third, to characterize the mechanisms during DHA production, a transcriptional analysis of the heterotrophic C. cohnii under different cultivation stages and conditions was carried out. By analyzing and comparing the differential gene expression profiles during lipid accumulation and DHA formation stages, core metabolism pathways in C. cohnii were proposed (Figure 3) [72]. In addition, transcripts related to fatty acid biosynthesis, starch and sucrose metabolism, and unsaturated fatty acids biosynthesis were significantly up-regulated during late-stage fermentation [72]. The study also found that some polyketide synthases and fatty acid desaturases may be involved in the biosynthesis of DHA. In addition, some enzymes involved in reducing power metabolism, such as malic enzyme and isocitrate dehydrogenase were up-regulated 1.7- to 2.3-fold during the lipid and DHA accumulation stages respectively, while the transcript of glucose-6-phosphate 1-dehydrogenase was downregulated 0.86- to 0.57-fold, suggesting that C. cohnii might use the malic enzyme and isocitrate dehydrogenase instead of glucose-6-phosphate 1-dehydrogenase to produce NADPH (nicotinamide adenine dinucleotide phosphate) [137]. Furthermore, C. cohnii mutants with high DHA productivity were also examined through transcriptomics analysis [12]. It was found that gene expression levels involved in fatty acid biosynthesis, energy, central carbohydrate, and amino acid metabolism were upregulated in the mutant with high DHA productivity, compared to the wild type. These results provide a basis for understanding for improving lipid accumulation and DHA production by rational engineering of C. cohnii in the future.
Although transcriptomic analysis provides a ‘‘snap shot’’ of gene expression under specific environmental and physiological conditions, it was also found that mRNA expression levels sometimes have a poor correlational relationship with the phenotype in dinoflagellates. For example, oscillating RNAs were not detected over the circadian cycle of the dinoflagellate Lingulodinium polyedrum [138]. They also discovered that the timing of the bioluminescence and photosynthesis rhythms remained unchanged, even when transcription rates had decreased to about 5% of the levels of untreated cells with the addition of inhibitors actinomycin D and cordycepin. Furthermore, there were no detectable changes in gene expression across the two types of Symbiodiniaceae (one was thermotolerant type and the other was more susceptible type) under thermal stress, even as the symbiosis was breaking down [64]. Some of these phenomena could be partially due to many cellular mechanisms acting at a post-transcriptional level. Another important feature is the generally low degree of congruency between mRNA and protein expression. For example, for the 167 proteins in Symbiodiniaceae downregulated at variable temperatures, only two corresponding mRNAs were differentially expressed between different treatments, while for 378 differentially expressed genes, none of their corresponding proteins was differentially expressed [68]. Therefore, it is worth noting the inherent risk of inferring cellular behavior based on transcriptional data alone. More and integrated omics or physiological analysis should be carried out for the full confirmation of metabolic and physiological behaviors of dinoflagellates.
2.3. Proteomic Analysis of Marine Dinoflagellates
As the main component of cellular structure and communication, protein is believed to be a more relevant indicator of an observed cellular phenotype than RNA [139]. Two major strategies for the separation and visualization of proteins are applied in proteomics: i) the two dimensional gel electrophoresis (2-DE) strategy, which separates proteins based on their masses and isoelectric points, followed by mass spectrometric identification; ii) gel-free profiling procedures, which rely on multidimensional separations coupling micro-scale separations with automated tandem mass spectrometry [140,141]. In the past decades, the proteomics technology has been rapidly developed, which make it possible to more directly probe the cellular behaviors of dinoflagellates than before [26,142].
Gel-free proteomics techniques, such as matrix-assisted laser desorption ionization-time-of-flight mass spectrometry (MALDI-TOF-MS) [143] and capillary liquid chromatography followed by tandem mass spectrometry (LC-MS) [144], have been applied to dinoflagellates proteomes. In particular, a non-gel based quantitative proteomic method named iTRAQ (isobaric Tags for Relative and Absolute Quantification), which quantifies proteins based on peptide labeling and allows large-scale identification and accurate quantification of proteins from multiple samples within broad dynamic ranges of protein abundance, have been conducted for A. catenella [58] or Karlodinium veneficum [145]. In the study, a total of 3488 or 4922 proteins were successfully identified from the proteomics of A. catenella or K. veneficum respectively, which are the highest number of proteins identified in dinoflagellates. Comparing the protein profiles of a toxin-producing dinoflagellate A. catenella (ACHK-T) and its non-toxic mutant (ACHK-NT) using a combination of iTRAQ-based proteomic approach and a transcriptomic database, they found different carbon and energy utilization strategies between ACHK-T and ACHK-NT, and discovered seven cyanobacterial toxin-related proteins and sxtG of dinoflagellates, which were identified as candidates involved in toxin biosynthesis but had no obvious difference between the two strains [58]. In K. veneficum, the changes in non-photochemical quenching and molecular mechanism under phosphorus deprivation were studied [145]. Proteomics results based on iTRAQ showed that non-photochemical quenching in K. veneficum increased significantly under phosphorus deprivation. Correspondingly, three light-harvesting complex stress-related proteins and energy production- and conversion-related proteins were up-regulated, while many proteins related to genetic information flow were down-regulated. It is expected that the iTRAQ-based proteomic approach could be utilized more frequently in future work as increasing genomic information on the dinoflagellates appears. However, proteins identified by iTRAQ-based proteomic approach are still far fewer than genes identified by genomics and transcriptomics approaches. One of the important reasons for this is the tedious and immature protein extraction procedures (including cell wall disruption and protein extraction), as most dinoflagellates have a tough and complex cell cortex leading to incomplete cell wall breakage and chemical contamination during extraction, which can interfere protein analysis [142]. Another important reason is protein identification, which is detailed below.
Proteomics has shown its powerful utilization in exploring the physiological and metabolic characteristics of dinoflagellates. Proteomics analysis has been used to identify new dinoflagellates species [48,143,146,147,148,149]. Traditional morphological approaches have been widely applied in dinoflagellate species identification, but these methods exhibit weaknesses in distinguishing closely related species with similar morphologies [26]. Using two-dimensional gel electrophoresis (2-DE) proteomics technology, species-specific protein expression profiles and standards of species identification in ten dinoflagellate species were established [147]. 2-DE has also been used to differentiate morphospecies (toxic and nontoxic) of A. minutum, and differentially expressed proteins between different morphospecies were observed at the proteome level [143]. Proteomics has also been employed to discover proteins of Symbiodiniaceae [150] and to reveal proteins involved in symbiosis and responses to environmental stress [151]. Using proteomics based on LC–MS/MS, 417 protein spots were identified in the endosymbiotic zooxanthellae from Euphyllia glabrescens and three marker proteins (green fluorescent protein R7, Histone H2B, and peridinin chlorophyll-a binding protein) were also found [150]. A total of 8098 MS/MS spectra relevant to peptides from the fraction of endosymbiont were identified, while only 26 peptides showed a significant change when treated with thermal stress or ion limitation. Surprisingly, the expression levels of proteins related to antioxidant or heat stress phenotypes were roughly the same, while proteins involved in protein biosynthesis were highly expressed. Proteomics has also been used to compare the performance of different Symbiodiniaceae species [144,152]. Using LC-MS based proteomics, they compared proteomes of the model sea anemone Exaiptasia pallida colonized by different dinoflagellate symbiont (B. minutum and Durusdinium trenchii) [144]. Results showed that in anemones containing D. trenchii (heterologous symbiont), Niemann-Pick C2 proteins, and glutamine synthetases were lowly expressed, while methionine-synthesizing betaine–homocysteine S-methyltransferases and proteins with predicted oxidative stress response functions were highly expressed when compared with anemones containing B. minutum (homologous symbiont). In another study, the researchers found that high-density symbiotic colonization required avoiding the immune responses of host cells, enhancing ammonium regulation, and inhibiting the phagocytosis of host cells after colonization [152]. iv) Proteins related to toxin biosynthesis were identified in A. catenella through proteomics analysis [153]. The proteomic results showed that the abundance of nine proteins with known functions in paralytic shellfish toxin-producing cyanobacteria was differentially regulated between toxin biosynthesis stages, suggesting that they might be involved in toxin biosynthesis in A. catenella.
In spite of the progress made, proteomics analysis of dinoflagellates still faces some technical challenges, especially for protein identification and validation. Protein identification largely depends on relevant genomic sequences, but so far only six draft whole genome sequences of dinoflagellates are available, and transcriptomes are still insufficient as a substitute for the query due to low coverage. In addition, peptides often contain post-translational modifications that lead to mass change of the fragments, thus hindering identification. Nevertheless, with the rapid development of genomics and transcriptomics in the near future, it is expected that proteomics will become a more powerful tool for studying dinoflagellates biology.
2.4. Metabolomic Analysis of Marine Dinoflagellates
Metabolomics is a global interrogation of cellular components, and has been gradually considered as a crucial supplement to other omics in exploring the mechanisms of dinoflagellates biology [154]. The advantages of metabolomics lie in its ability to detect qualitative and quantitative changes in a large number of metabolites using mass spectrometry and nuclear magnetic resonance spectroscopy [154]. Although there are a number of high-resolution technologies capable of detecting and identifying changes in metabolite profiles, our understanding of how these differently expressed metabolites affect overall biological function is still in infancy [155,156,157]. Currently, gas chromatography time-of-flight mass spectrometry (GC-TOF-MS), high-performance liquid chromatography mass spectrometry (LC-MS), and 13C-labeling based metabolic flux analyses have been mainly used in dinoflagellates metabolomics analysis (Table 3) [69,73,158,159].
Although the application of metabolomics to dinoflagellates is relatively new, it has already made significant contributions to dinoflagellates biology research (Table 3): i) metabolomics studies focusing on lipogenetic mechanisms of C. cohnii have been conducted. For example, metabolomic analysis has been used to identify metabolic modules and hub metabolites related to the positive role of the antioxidant butylated hydroxyanisole on lipogenesis in C. cohnii [13], to determine mechanisms relevant to glucose tolerance through adaptive laboratory evolution [11], the responsive metabolites associated with varying dissolved oxygen levels [74], as well as the metabolic changes among different nitrogen feeding fermentation conditions in C. cohnii [76]; ii) metabolite profiles related to several growth conditions of marine symbiotic dinoflagellates of Symbiodiniaceae were also examined using GC-MS based metabolomic analysis [161], and the results showed that both the production of sterols and sugars and the abundance of hexose and inositol were different between different Symbiodiniaceae species. As acidification markedly inhibited B. minutum growth [164], the responsive metabolite profile of B. minutum to acidification was studied [162]. The results showed that: a) saturated fatty acids and oligosaccharides were accumulated when B. minutum was cultured in the acidification condition, which was considered as an important strategy to adapt to acidification; and b) the inhibition of the growth rate was possibly due to the affection of acidification to the biosynthesis of amino acids and proteins of B. minutum; iii) 13C labelling coupled with GC-MS was used to track carbon changes in metabolic flux dinoflagellates. For example, autotrophic carbon fate was mapped in both Aiptasia and Symbiodiniaceae (a model cnidarian-dinoflagellate symbiosis) exposed to thermal stress [69]. A newly fixed carbon fate in the model cnidarian Exaiptasia pallida was characterized with colonized with either native B. minutum or non-native D. trenchii using 13C-labeling coupled with GC-MS [159]. Different abundance and diversity of metabolites were detected between anemones colonized with different Symbiodiniaceae species and significant alterations to host molecular signaling pathways were also revealed. For heterotrophic C. cohnii, 13C-labeling based metabolic flux analysis has been used to explore the central flux distribution after addition of ethanolamine to stimulate lipid accumulation [73]. It was found that activity of the glycolysis pathway and citrate pyruvate cycle was increased, while that of the pentose phosphate pathway and TCA cycle was attenuated after adding ethanolamine; iv) exometabolomics, which focused on the research of complete small molecules cells secret into their environments [165], was also used for investigating interactions of different dinoflagellates species. For example, when Ostreopsis cf. ovata was co-cultivated with diatom Licmophora paradoxa, the exometabolome of L. paradoxa was found impair the growth and the photochemistry of Ostreopsis cf. ovata in both bioassays and co-cultures, and some biomarkers possibly involved in the inhibition effects were later identified using a metabolomic approach [61].
Although the comprehensiveness and measurement accuracy of metabolomics still needs further improvements [155,156,157], early research has demonstrated that metabolomics is a powerful tool in exploring metabolism and filling the phenotype-genotype gap because it presents a closer picture of cellular activity than other omics methods [157]. Furthermore, metabolomics is considered as a more promising approach for interpreting the basic characteristics of dinoflagellates without sequencing entire genomes.
2.5. Integrated Omics Analysis of Marine Dinoflagellates
In recent years, it has been realized that single “omics” analysis is not sufficient for characterizing the complexity of dinoflagellate biological systems. Therefore, integration of multiple “omics” approach is required to obtain a relatively precise picture of dinoflagellates. Some attempts have been made recently to integrate heterogeneous “omics” datasets in various microbial systems, and the results have demonstrated that the “multi-omics” method is a powerful tool for understanding the functional principles and dynamics of total cellular systems [37,166,167,168,169,170].
The integration of the “multi-omics” approach was also applied to explore complex metabolic networks and global regulatory mechanisms in dinoflagellates [68,71,72,171]. For example, i) integrated transcriptomic and metabolomic analysis was applied to characterize the molecular and physiological processes of DHA synthesis in the fed-batch fermentation of C. cohnii, and differently expressed genes involved in fatty acid and DHA biosynthesis were discovered [72]; ii) integrated transcriptomic and metabolomic analysis was also used to compare the effects on the sea anemone Exaiptasia pallida when colonized by a homologous symbiont B. minutum and a heterologous, opportunistic, and thermally tolerant symbiont D. trenchii [171]. It was shown that the catabolism of fixed carbon storage, metabolic signals, and immune processes were increased in E. pallide accompanied with heterogenous symbiosis D. trenchii, in comparison with homologous B. minutum colonized hosts; iii) integrated transcriptomic and proteomic analysis was used to study the temperature adaptation mechanism of Zooxanthellate cnidarians (the Red Sea (RS), North Carolina (CC7), and Hawaii (H2)) [71]. A common core response to thermal stress (24 h at 32 °C), containing protein folding and oxidative stress pathways, was highlighted through comparisons. The level of antioxidant gene expression was increased in all three anemones, while RS anemones showed the greatest increase. Reactive oxygen species production was different between three strains, which were symbiont-driven, while RS anemones showed significantly lower levels in hospite [68].
When using the multi-omics approach, there still are many tough challenges. For example, low correlation between the transcript and protein levels, which might be due to posttranscriptional regulation of gene expression or a lack of the proper statistical tools for biological interpretation [37]. In addition to the risk of using single omics analysis, these is also a need to develop new bioinformatics tools and improve the availability of public data repositories.
3. Conclusions and Prospective Research
Omics research on dinoflagellates has attracted great interest in the past few decades, as it has substantially contributed to the better understanding of the dinoflagellates at the molecular level. However, there are still challenges in omics analysis, and the risks of only using one kind of omics technology to explore dinoflagellates biology deserve attention.
In future studies, several aspects of omics research on dinoflagellates should be strengthened. First, more attention should be paid to the complete genome sequencing of dinoflagellates. The newly developed DNA sequencing technologies, such as single-molecule DNA sequencing, could compensate for the shortcomings of NGS for this purpose, by acting as long scaffolds to resolve highly repetitive genomic regions [172], and make whole-genome sequencing of dinoflagellates much more feasible. Second, strengthening the description of the proteome/metabolome of a dinoflagellate, and establishing a reference proteome/metabolome of a dinoflagellate. These would will greatly increase the rate of discovery in dinoflagellate biology and reveal biomarkers for certain functional characteristics or responses of dinoflagellates. Third, integrated multi-omics analysis is important for exploring dynamic process spanning multiple cellular components in dinoflagellates [37]. Functional genomics data sets, such as transcriptomics, proteomics, and genome-wide mutant screens, can provide additional layers of gene-specific functional data. This integrated approach can minimize errors during analysis and the result will be more comprehensive and accurate. Fourth, it is urgent that the mechanism of HABs formation and toxin biosynthesis receive intensive study. In recent years, there has been a negative influence of dinoflagellate blooms on the environment and human society, as they are occurring more frequently and becoming larger in scale [173]. Finally, as transformation systems for selective species of dinoflagellates, such as Amphidinium, S. microadriaticum, Symbiodiniaceae, and C. cohnii, have been reported [75,87,174], the results obtained from omics analysis can provide more targets and pathways to accelerate the process of genetic engineering in dinoflagellates. More mutants with superior characteristics, such as C. cohnii, with higher fractions of DHA, or Symbiodiniaceae, with the ability to tolerate higher temperatures will be constructed.
Funding
This research was supported by grants from National Key R&D Program of China (Nos. 2018YFA0903000 and 2018YFA0903600) and the National Natural Science Foundation of China (Nos. 31770100, 91751102, 21621004, 31370115 and 31470217) and Zaoneng Biotechnology Inc.
Acknowledgments
We thank Lei Chen of Tianjin University for the valuable discussion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Matthiessen, J.; Schreck, M. Dinoflagellates. In Encyclopedia of Marine Geosciences; Springer: Dordrecht, The Netherlands, 2015; pp. 1–7. [Google Scholar] [Green Version]
- Costas, E.; Goyanes, V. Architecture and evolution of dinoflagellate chromosomes: An enigmatic origin. Cytogenet. Genome Res. 2005, 109, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Yoo, Y.D.; Kim, J.S.; Seong, K.A.; Kang, N.S.; Kim, T.H. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci. J. 2010, 45, 65–91. [Google Scholar] [CrossRef] [Green Version]
- Mendes, A.; Reis, A.; Vasconcelos, R.; Guerra, P.; Silva, T.L.D. Crypthecodinium cohnii with emphasis on DHA production: A review. J. Appl. Phycol. 2008, 21, 199–214. [Google Scholar] [CrossRef]
- Fukuda, Y.; Suzaki, T. Unusual Features of Dinokaryon, the Enigmatic Nucleus of Dinoflagellates. In Marine Protists; Springer: Tokyo, Japan, 2015; pp. 23–45. [Google Scholar]
- Muscatine, L.; Porter, J.W. Reef corals: Mutualistic Symbioses Adapted to Nutrient-Poor Environments. Bioscience 1977, 1977, 454–460. [Google Scholar] [CrossRef]
- Liu, H.; Stephens, T.G.; Gonzalez-Pech, R.A.; Beltran, V.H.; Lapeyre, B.; Bongaerts, P.; Cooke, I.; Aranda, M.; Bourne, D.G.; Foret, S.; et al. Symbiodinium genomes reveal adaptive evolution of functions related to coral-dinoflagellate symbiosis. Commun. Biol. 2018, 1, 95. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.; Calderon, F.; Wen, Z.; Kim, H.Y. Docosahexaenoic Acid: A positive modulator of Akt signaling in neuronal survival. Proc. Natl. Acad. Sci. USA 2005, 102, 10858–10863. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Lim, G.P.; Yang, F.; Morihara, T.; Teter, B.; Ubeda, O.; Rostaing, P.; Triller, A.; Salem, N., Jr.; Ashe, K.H. Docosahexaenoic acid protects from dendritic pathology in an Alzheimer’s disease mouse model. Neuron 2004, 43, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Gronert, K.; Devchand, P.R.; Moussignac, R.L.; Serhan, C.N. Novel docosatrienes and 17s-resolvins generated from docosahexaenoic acid in murine brain, human blood, and glial cells autacoids in anti-inflammation. J. Biol. Chem. 2003, 278, 14677–14687. [Google Scholar] [CrossRef] [PubMed]
- Li, X.R.; Pei, G.S.; Liu, L.S.; Chen, L.; Zhang, W.W. Metabolomic analysis and lipid accumulation in a glucose tolerant Crypthecodinium cohnii strain obtained by adaptive laboratory evolution. Bioresour. Technol. 2017, 235, 87–95. [Google Scholar] [CrossRef]
- Liu, J.; Pei, G.S.; Diao, J.J.; Chen, Z.X.; Liu, L.S.; Chen, L.; Zhang, W.W. Screening and transcriptomic analysis of Crypthecodinium cohnii mutants with high growth and lipid content using the acetyl-CoA carboxylase inhibitor sethoxydim. Appl. Microbiol. Biotechnol. 2017, 101, 6179–6191. [Google Scholar] [CrossRef]
- Sui, X.; Niu, X.F.; Shi, M.L.; Pei, G.S.; Li, J.H.; Chen, L.; Wang, J.X.; Zhang, W.W. Metabolomic analysis reveals mechanism of antioxidant butylated hydroxyanisole on lipid accumulation in Crypthecodinium cohnii. J. Agric. Food Chem. 2014, 62, 12477–12484. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, J.; Sun, P.P.; Ma, X.N.; Jiang, Y.; Chen, F. Sesamol Enhances Cell Growth and the Biosynthesis and Accumulation of Docosahexaenoic Acid in the Microalga Crypthecodinium cohnii. J. Agric. Food Chem. 2015, 63, 5640–5645. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.J.; Song, X.Y.; Cui, J.Y.; Liu, L.S.; Shi, M.L.; Wang, F.Z.; Zhang, W.W. Rewiring metabolic network by chemical modulator based laboratory evolution doubles lipid production in Crypthecodinium cohnii. Metab. Eng. 2019, 51, 88–98. [Google Scholar] [CrossRef]
- Rizzo, P.J.; Jones, M.; Ray, S.M. Isolation and Properties of Isolated Nuclei from the Florida Red Tide Dinoflagellate Gymnodinium breve (Davis). J. Protozool. 1982, 29, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Fogel, M.; Schmitter, R.E.; Hastings, J.W. On the Physical Identity of Scintillons: Bioluminescent Particles in Gonyaulax Polyedra. J. Cell Sci. Ther. 1972, 11, 305–317. [Google Scholar]
- Desjardins, M.; Morse, D. The polypeptide components of scintillons, the bioluminescence organelles of the dinoflagellate Gonyaulax polyedra. Biochem. Cell Biol. 1993, 71, 3–4. [Google Scholar] [CrossRef]
- Hackett, J.D.; Anderson, D.M.; Erdner, D.L.; Bhattachary, D. Dinoflagellates: A remarkable evolutionary experiment. Am. J. Bot. 2004, 91, 1523–1534. [Google Scholar] [CrossRef]
- Buskey, E.J.; Swift, E. Behavioral responses of the coastal copepod Acartia hudsonica (Pinhey) to simulated dinoflagellate bioluminescence. J. Exp. Mar. Biol. Ecol. 1983, 72, 43–58. [Google Scholar] [CrossRef]
- Gavelis, G.S.; Hayakawa, S.; White, R.A.; Gojobori, T.; Suttle, C.A.; Keeling, P.J.; Leander, B.S. Eye-like ocelloids are built from different endosymbiotically acquired components. Nature 2015, 523, 204–207. [Google Scholar] [CrossRef]
- Keeling, P.J. The endosymbiotic origin, diversification and fate of plastids. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2010, 365, 729–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisecaver, J.H.; Hackett, J.D. Dinoflagellate genome evolution. Annu. Rev. Microbiol. 2011, 65, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Spector, D.L. Dinoflagellate Nuclei. In Dinoflagellates; Academic Press: Cambridge, MA, USA, 1984; pp. 107–147. [Google Scholar]
- Wang, D.Z.; Zhang, H.; Zhang, Y.; Zhang, S.F. Marine dinoflagellate proteomics: Current status and future perspectives. J. Proteomics 2014, 105, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Rae, P. Hydroxymethyluracil in eukaryote DNA: A natural feature of the pyrrophyta (dinoflagellates). Science 1976, 194, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.; Jakobsen, K.S.; Nordby, Ø. Characterization of DNA from the dinoflagellate woloszynskia bostoniensis. J. Prolozool. 1988, 35, 418–422. [Google Scholar] [CrossRef]
- Boorstein, R.J.; Chiu, L.N.; Teebor, G.W. Phylogenetic evidence of a role for 5-hydroxymethyluracil-DNA glycosylase in the maintenance of 5-methylcytosine in DNA. Nucleic Acids Res. 1989, 17, 7653–7661. [Google Scholar] [CrossRef] [Green Version]
- Bouligand, Y.; Norris, V. Chromosome separation and segregation in dinoflagellates and bacteria may depend on liquid crystalline states. Biochimie 2001, 83, 187–192. [Google Scholar] [CrossRef]
- Chan, Y.H.; Wong, J.T. Concentration-dependent organization of DNA by the dinoflagellate histone-like protein HCc3. Nucleic Acids Res. 2007, 35, 2573–2583. [Google Scholar] [CrossRef] [Green Version]
- Janouskovec, J.; Gavelis, G.S.; Burki, F.; Dinh, D.; Bachvaroff, T.R.; Gornik, S.G.; Bright, K.J.; Imanian, B.; Strom, S.L.; Delwiche, C.F.; et al. Major transitions in dinoflagellate evolution unveiled by phylotranscriptomics. Proc. Natl. Acad. Sci. USA 2017, 114, E171–E180. [Google Scholar] [CrossRef]
- Gornik, S.G.; Ford, K.L.; Mulhern, T.D.; Bacic, A.; McFadden, G.I.; Waller, R.F. Loss of nucleosomal DNA condensation coincides with appearance of a novel nuclear protein in dinoflagellates. Curr. Biol. 2012, 22, 2303–2312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Green, B.R.; Cavalier-Smith, T. Single gene circles in dinoflagellate chloroplast genomes. Nature 1999, 400, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.F.; Jackson, C.J. Dinoflagellate mitochondrial genomes: Stretching the rules of molecular biology. Bioessays 2009, 31, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, F.; Nie, L. Integrating multiple ‘omics’ analysis for microbial biology: Application and methodologies. Microbiology 2010, 156, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Shoguchi, E.; Shinzato, C.; Kawashima, T.; Gyoja, F.; Mungpakdee, S.; Koyanagi, R.; Takeuchi, T.; Hisata, K.; Tanaka, M.; Fujiwara, M.; et al. Draft assembly of the Symbiodinium minutum nuclear genome reveals dinoflagellate gene structure. Curr. Biol. 2013, 23, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z.; et al. The Symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Aranda, M.; Li, Y.; Liew, Y.J.; Baumgarten, S.; Simakov, O.; Wilson, M.C.; Piel, J.; Ashoor, H.; Bougouffa, S.; Bajic, V.B.; et al. Genomes of coral dinoflagellate symbionts highlight evolutionary adaptations conducive to a symbiotic lifestyle. Sci. Rep. 2016, 6, 39734. [Google Scholar] [CrossRef] [Green Version]
- Shoguchi, E.; Beedessee, G.; Tada, I.; Hisata, K.; Kawashima, T.; Takeuchi, T.; Arakaki, N.; Fujie, M.; Koyanagi, R.; Roy, M.C.; et al. Two divergent Symbiodinium genomes reveal conservation of a gene cluster for sunscreen biosynthesis and recently lost genes. BMC Genom. 2018, 19, 458. [Google Scholar] [CrossRef]
- Chan, L.L.; Sit, W.H.; Lam, P.K.; Hsieh, D.P.; Hodgkiss, I.J.; Wan, J.M.; Ho, A.Y.; Choi, N.M.; Wang, D.Z.; Dudgeon, D. Identification and characterization of a “biomarker of toxicity” from the proteome of the paralytic shellfish toxin-producing dinoflagellate Alexandrium tamarense (Dinophyceae). Proteomics 2006, 6, 654–666. [Google Scholar] [CrossRef]
- Uribe, P.; Fuentes, D.; Valdes, J.; Shmaryahu, A.; Zuniga, A.; Holmes, D.; Valenzuela, P.D. Preparation and analysis of an expressed sequence tag library from the toxic dinoflagellate Alexandrium catenella. Mar. Biotechnol. 2008, 10, 692–700. [Google Scholar] [CrossRef]
- Erdner, D.L.; Anderson, D.M. Global transcriptional profiling of the toxic dinoflagellate Alexandrium fundyense using Massively Parallel Signature Sequencing. BMC Genom. 2006, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.Y.; Lin, C.H.; Liu, J.K.; Wong, T.Y.; Kuo, J. Analysis of Codon Usage Patterns in Toxic Dinoflagellate Alexandrium tamarense through Expressed Sequence Tag Data. Comp. Funct. Genom. 2010, 2010, 138538. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Evans, A.N.; Kulis, D.M.; Hackett, J.D.; Erdner, D.L.; Anderson, D.M.; Bhattacharya, D. Transcriptome Profiling of a Toxic Dinoflagellate Reveals a Gene-Rich Protist and a Potential Impact on Gene Expression Due to Bacterial Presence. PLoS ONE 2010, 5, e9688. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.; John, U.; Beszteri, S.; Glockner, G.; Krock, B.; Goesmann, A.; Cembella, A.D. Comparative gene expression in toxic versus non-toxic strains of the marine dinoflagellate Alexandrium minutum. BMC Genom. 2010, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Dong, H.P.; Li, C.; Xie, Z.X.; Lin, L.; Hong, H.S. Identification and Characterization of Cell Wall Proteins of a Toxic Dinoflagellate Alexandrium catenella Using 2-D DIGE and MALDI TOF-TOF Mass Spectrometry. Evid.-Based Complement. Altern. 2011, 2011, 984080. [Google Scholar]
- Chan, L.L.; Li, X.; Sit, W.H.; Lam, K.S.; Leung, M.Y. Development of theca specific antisera for the profiling of cell surface proteins in the marine toxic dinoflagellate genus Alexandrium halim. Harmful Algae 2012, 16, 58–62. [Google Scholar] [CrossRef]
- Li, C.; Wang, D.Z.; Dong, H.P.; Xie, Z.X.; Hong, H.S. Proteomics of a toxic dinoflagellate Alexandrium catenella DH01: Detection and identification of cell surface proteins using fluorescent labeling. Chin. Sci. Bull. 2012, 57, 3320–3327. [Google Scholar] [CrossRef]
- Wang, D.Z.; Li, C.; Zhang, Y.; Wang, Y.Y.; He, Z.P.; Lin, L.; Hong, H.S. Quantitative proteomic analysis of differentially expressed proteins in the toxicity-lost mutant of Alexandrium catenella (Dinophyceae) in the exponential phase. J. Proteomics 2012, 75, 5564–5577. [Google Scholar] [CrossRef]
- Wang, D.Z.; Lin, L.; Wang, M.H.; Li, C.; Hong, H.S. Proteomic analysis of a toxic dinoflagellate Alexandrium catenella under different growth phases and conditions. Chin. Sci. Bull. 2012, 57, 3328–3341. [Google Scholar] [CrossRef]
- Gao, D.; Qiu, L.; Hou, Z.; Zhang, Q.; Wu, J.; Gao, Q.; Song, L. Computational Identification of MicroRNAs from the Expressed Sequence Tags of Toxic Dinoflagellate Alexandrium Tamarense. Evolut. Bioinform. 2013, 9, 479–485. [Google Scholar] [CrossRef]
- Pawlowiez, R.; Morey, J.S.; Darius, H.T.; Chinain, M.; Van Dolah, F.M. Transcriptome sequencing reveals single domain Type I-like polyketide synthases in the toxic dinoflagellate Gambierdiscus polynesiensis. Harmful Algae 2014, 36, 29–37. [Google Scholar] [CrossRef]
- Zhang, S.; Sui, Z.; Chang, L.; Kang, K.; Ma, J.; Kong, F.; Zhou, W.; Wang, J.; Guo, L.; Geng, H.; et al. Transcriptome de novo assembly sequencing and analysis of the toxic dinoflagellate Alexandrium catenella using the Illumina platform. Gene 2014, 537, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.F.; Lin, L.; Wang, D.Z. Comparative transcriptome analysis of a toxin-producing dinoflagellate Alexandrium catenella and its non-toxic mutant. Mar. Drugs 2014, 12, 5698–5718. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.W.; Wang, J.; Chan, L.L.; Lam, P.K.; Gu, J.D. Comparison of three protein extraction procedures from toxic and non-toxic dinoflagellates for proteomics analysis. Ecotoxicology 2015, 24, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.F.; Zhang, Y.; Xie, Z.X.; Zhang, H.; Lin, L.; Wang, D.Z. iTRAQ-based quantitative proteomic analysis of a toxigenic dinoflagellate Alexandrium catenella and its non-toxic mutant. Proteomics 2015, 15, 4041–4050. [Google Scholar] [CrossRef] [PubMed]
- Jean, N.; Dumont, E.; Herzi, F.; Balliau, T.; Laabir, M.; Masseret, E.; Mounier, S. Modifications of the soluble proteome of a mediterranean strain of the invasive neurotoxic dinoflagellate Alexandrium catenella under metal stress conditions. Aquat. Toxicol. 2017, 188, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.F.; Lin, L.; Wang, D.Z. Whole Transcriptomic Analysis Provides Insights into Molecular Mechanisms for Toxin Biosynthesis in a Toxic Dinoflagellate Alexandrium catenella (ACHK-T). Toxins 2017, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Ternon, E.; Pavaux, A.S.; Marro, S.; Thomas, O.P.; Lemee, R. Allelopathic interactions between the benthic toxic dinoflagellate Ostreopsis cf. ovata and a co-occurring diatom. Harmful Algae 2018, 75, 35–44. [Google Scholar] [CrossRef]
- Kuo, J.; Chen, M.C.; Lin, C.H.; Fang, L.S. Comparative gene expression in the symbiotic and aposymbiotic Aiptasia pulchella by expressed sequence tag analysis. Biochem. Biophys. Res. Commun. 2004, 318, 176–186. [Google Scholar] [CrossRef]
- Bayer, T.; Aranda, M.; Sunagawa, S.; Yum, L.K.; Desalvo, M.K.; Lindquist, E.; Coffroth, M.A.; Voolstra, C.R.; Medina, M. Symbiodinium transcriptomes: Genome insights into the dinoflagellate symbionts of reef-building corals. PLoS ONE 2012, 7, e35269. [Google Scholar] [CrossRef]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Palumbi, S.R. Lineage-specific transcriptional profiles of Symbiodinium spp. unaltered by heat stress in a coral host. Mol. Biol. Evolut. 2014, 31, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Krueger, T.; Fisher, P.L.; Becker, S.; Pontasch, S.; Dove, S.; Hoegh-Guldberg, O.; Leggat, W.; Davy, S.W. Transcriptomic characterization of the enzymatic antioxidants FeSOD, MnSOD, APX and KatG in the dinoflagellate genus Symbiodinium. BMC Evolut. Biol. 2015, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Nelson, W.; Rodriguez, J.; Tolleter, D.; Grossman, A.R. Symbiodinium transcriptome and global responses of cells to immediate changes in light intensity when grown under autotrophic or mixotrophic conditions. Plant J. 2015, 82, 67–80. [Google Scholar] [CrossRef]
- Levin, R.A.; Beltran, V.H.; Hill, R.; Kjelleberg, S.; McDougald, D.; Steinberg, P.D.; van Oppen, M.J. Sex, Scavengers, and Chaperones: Transcriptome Secrets of Divergent Symbiodinium Thermal Tolerances. Mol. Biol. Evolut. 2016, 33, 2201–2215. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.B.; Wang, Y.B.; Chen, C.S.; Chen, S.H.; Lin, C.Y. Dual-compartmental transcriptomic + proteomic analysis of a marine endosymbiosis exposed to environmental change. Mol. Ecol. 2016, 25, 5944–5958. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, K.E.; Dias, D.A.; Lutz, A.; Roessner, U.; Davy, S.K. Mapping carbon fate during bleaching in a model cnidarian symbiosis: The application of (13) C metabolomics. New Phytol. 2017, 214, 1551–1562. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.A.; Voolstra, C.R.; Weynberg, K.D.; van Oppen, M.J. Evidence for a role of viruses in the thermal sensitivity of coral photosymbionts. ISME J. 2017, 11, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Cziesielski, M.J.; Liew, Y.J.; Cui, G.; Schmidt-Roach, S.; Campana, S.; Marondedze, C.; Aranda, M. Multi-omics analysis of thermal stress response in a zooxanthellate cnidarian reveals the importance of associating with thermotolerant symbionts. Proc. Biol. Sci. 2018, 285, 20172654. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.S.; Li, X.R.; Liu, L.S.; Liu, J.; Wang, F.Z.; Chen, L.; Zhang, W.W. De novo transcriptomic and metabolomic analysis of docosahexaenoic acid (DHA)-producing Crypthecodinium cohnii during fed-batch fermentation. Algal Res. 2017, 26, 380–391. [Google Scholar] [CrossRef]
- Cui, J.Y.; Diao, J.J.; Sun, T.; Shi, M.L.; Liu, L.; Wang, F.Z.; Chen, L.; Zhang, W.W. (13)C Metabolic Flux Analysis of Enhanced Lipid Accumulation Modulated by Ethanolamine in Crypthecodinium cohnii. Front. Microbiol. 2018, 9, 956. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.J.; Li, X.R.; Pei, G.S.; Liu, L.; Chen, L. Comparative metabolomic analysis of Crypthecodinium cohnii in response to different dissolved oxygen levels during docosahexaenoic acid fermentation. Biochem. Biophys. Res. Commun. 2018, 499, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.J.; Song, X.Y.; Zhang, X.Q.; Chen, L.; Zhang, W.W. Genetic Engineering of Crypthecodinium cohnii to Increase Growth and Lipid Accumulation. Front. Microbiol. 2018, 9, 492. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.S.; Wang, F.Z.; Yang, J.; Li, X.R.; Cui, J.Y.; Liu, J.; Shi, M.L.; Wang, K.; Chen, L.; Zhang, W.W. Nitrogen Feeding Strategies and Metabolomic Analysis to Alleviate High-Nitrogen Inhibition on Docosahexaenoic Acid Production in Crypthecodinium cohnii. J. Agric. Food Chem. 2018, 66, 10640–10650. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Marra, M. Applications of next-generation sequencing technologies in functional genomics. Genomics 2008, 92, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shendure, J.; Ji, H. Next-generation DNA sequencing. Nat. Biotechnol. 2008, 26, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Bonetta, L. Whole-genome sequencing breaks the cost barrier. Cell 2010, 141, 917–919. [Google Scholar] [CrossRef] [PubMed]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Lajeunesse, T.C.; Lambert, G.; Andersen, R.A.; Coffroth, M.A.; Galbraith, D.W. Symbiodinium (pyrrhophyta) genome sizes (DNA content) are smallest among dinoflagellates. J. Phycol. 2005, 41, 880–886. [Google Scholar] [CrossRef]
- Sutton, R.E.; Boothroyd, J.C. Evidence for Trans splicing in trypanosomes. Cell 1986, 47, 527–535. [Google Scholar] [CrossRef]
- Gonzalez-Pech, R.A.; Ragan, M.A.; Chan, C.X. Signatures of adaptation and symbiosis in genomes and transcriptomes of Symbiodinium. Sci. Rep. 2017, 7, 15021. [Google Scholar] [CrossRef]
- Sproles, A.E.; Kirk, N.L.; Kitchen, S.A.; Oakley, C.A.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Phylogenetic characterization of transporter proteins in the cnidarian-dinoflagellate symbiosis. Mol. Phylogen. Evolut. 2018, 120, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Qunjie, G.; Ferran, G.P. Microbial ultraviolet sunscreens. Nat. Rev. Microbiol. 2011, 9, 791. [Google Scholar]
- Wood-Charlson, E.M.; Hollingsworth, L.L.; Krupp, D.A.; Weis, V.M. Lectin/glycan interactions play a role in recognition in a coral/dinoflagellate symbiosis. Cell Microbiol. 2006, 8, 1985–1993. [Google Scholar] [CrossRef] [PubMed]
- Ten Lohuis, M.R.; Miller, D.J. Genetic transformation of dinoflagellates (Amphidinium and Symbiodinium): Expression of GUS in microalgae using heterologous promoter constructs. Plant J. 1998, 13, 427–435. [Google Scholar]
- Goodwin, S.; Mcpherson, J.D.; Mccombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Salzberg, S.L.; Yorke, J.A. Beware of mis-assembled genomes. Bioinformatics 2005, 21, 4320–4321. [Google Scholar] [CrossRef] [PubMed]
- Joachim, W.; Orsolya, S.; FranOis, S.; Korbel, J.O. Phenotypic impact of genomic structural variation: Insights from and for human disease. Nat. Rev. Genet. 2013, 14, 125–138. [Google Scholar]
- Van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Flusberg, B.A.; Webster, D.R.; Lee, J.H.; Travers, K.J.; Olivares, E.C.; Clark, T.A.; Korlach, J.; Turner, S.W. Direct detection of DNA methylation during single-molecule, real-time sequencing. Nat. Methods 2010, 7, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Rand, A.C.; Jain, M.; Eizenga, J.M.; Musselman-Brown, A.; Olsen, H.E.; Akeson, M.; Paten, B. Mapping DNA methylation with high-throughput nanopore sequencing. Nat. Methods 2017, 14, 411. [Google Scholar] [CrossRef]
- Simpson, J.T.; Workman, R.E.; Zuzarte, P.C.; David, M.; Dursi, L.J.; Timp, W. Detecting DNA cytosine methylation using nanopore sequencing. Nat. Methods 2017, 14, 407–410. [Google Scholar] [CrossRef]
- Horak, C.E.; Snyder, M. Global analysis of gene expression in yeast. Funct. Integr. Genom. 2002, 2, 171–180. [Google Scholar] [CrossRef]
- Ye, R.W.; Wang, T.; Bedzyk, L.; Croker, K.M. Applications of DNA microarrays in microbial systems. J. Microbiol. Methods 2001, 47, 257–272. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Royce, T.E.; Rozowsky, J.S.; Gerstein, M.B. Toward a universal microarray: Prediction of gene expression through nearest-neighbor probe sequence identification. Nucleic Acids Res. 2007, 35, e99. [Google Scholar] [CrossRef]
- Okoniewski, M.J.; Miller, C.J. Hybridization interactions between probesets in short oligo microarrays lead to spurious correlations. BMC Bioinform. 2006, 7, 276. [Google Scholar] [CrossRef]
- Boguski, M.S.; Tolstoshev, C.M.; Bassett, D.E. Gene discovery in dbEST. Science 1994, 265, 1993. [Google Scholar] [CrossRef]
- Brenner, S.; Johnson, M.; Bridgham, J.; Golda, G.; Lloyd, D.H.; Johnson, D.E.A. Gene expression analysis by massively parallel signature sequencing (MPSS) on microbead arrays. Nat. Biotechnol. 2000, 18, 630–634. [Google Scholar] [CrossRef]
- Lidie, K.B.; Ryan, J.C.; Barbier, M.; Dolah, F.M.V. Gene Expression in Florida Red Tide Dinoflagellate Karenia brevis: Analysis of an Expressed Sequence Tag Library and Development of DNA Microarray. Mar. Biotechnol. 2005, 7, 481–493. [Google Scholar] [CrossRef]
- Van Dolah, F.M.; Lidie, K.B.; Morey, J.S.; Brunelle, S.A.; Ryan, J.C.; Monroe, E.A.; Haynes, B.L. Microarray Analysis of Diurnal- and Circadian-Regulated Genes in the Florida Red-Tide Dinoflagellate karenia Brevis (Dinophyceae). J. Phycol. 2007, 43, 741–752. [Google Scholar] [CrossRef]
- Monroe, E.A.; Johnson, J.G.; Wang, Z.H.; Pierce, R.K.; Van Dolah, F.M. Characterization and Expression of Nuclear-Encoded Polyketide Synthases in the Brevetoxin-Producing Dinoflagellate Karenia Brevis. J. Phycol. 2010, 46, 541–552. [Google Scholar] [CrossRef]
- Morey, J.S.; Monroe, E.A.; Kinney, A.L.; Beal, M.; Johnson, J.G.; Hitchcock, G.L.; Van Dolah, F.M. Transcriptomic response of the red tide dinoflagellate, Karenia brevis, to nitrogen and phosphorus depletion and addition. BMC Genom. 2011, 12, 346. [Google Scholar] [CrossRef]
- Johnson, J.G.; Morey, J.S.; Neely, M.G.; Ryan, J.C.; Van Dolah, F.M. Transcriptome remodeling associated with chronological aging in the dinoflagellate, Karenia brevis. Mar. Genom. 2012, 5, 15–25. [Google Scholar] [CrossRef]
- Zhuang, Y.Y.; Zhang, H.; Hannick, L.; Lin, S.J. Metatranscriptome profiling reveals versatile N-nutrient utilization, CO2 limitation, oxidative stress, and active toxin production in an Alexandrium fundyense bloom. Harmful Algae 2015, 42, 60–70. [Google Scholar] [CrossRef]
- Yang, I.; Selander, E.; Pavia, H.; John, U. Grazer-induced toxin formation in dinoflagellates: A transcriptomic model study. Eur. J. Phycol. 2011, 46, 66–73. [Google Scholar] [CrossRef]
- Harke, M.J.; Juhl, A.R.; Haley, S.T.; Alexander, H.; Dyhrman, S.T. Conserved Transcriptional Responses to Nutrient Stress in Bloom-Forming Algae. Front. Microbiol. 2017, 8, 1279. [Google Scholar] [CrossRef]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glockner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative genomic and transcriptomic characterization of the toxigenic marine dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef]
- Lauritano, C.; De Luca, D.; Ferrarini, A.; Avanzato, C.; Minio, A.; Esposito, F.; Ianora, A. De novo transcriptome of the cosmopolitan dinoflagellate Amphidinium carterae to identify enzymes with biotechnological potential. Sci. Rep. 2017, 7, 11701. [Google Scholar] [CrossRef]
- Meyer, J.M.; Rodelsperger, C.; Eichholz, K.; Tillmann, U.; Cembella, A.; McGaughran, A.; John, U. Transcriptomic characterisation and genomic glimps into the toxigenic dinoflagellate Azadinium spinosum, with emphasis on polykeitde synthase genes. BMC Genom. 2015, 16, 27. [Google Scholar] [CrossRef]
- Wang, H.; Tomasch, J.; Michael, V.; Bhuju, S.; Jarek, M.; Petersen, J.; Wagner-Dobler, I. Identification of Genetic Modules Mediating the Jekyll and Hyde Interaction of Dinoroseobacter shibae with the Dinoflagellate Prorocentrum minimum. Front. Microbiol. 2015, 6, 1262. [Google Scholar] [CrossRef]
- Wang, H.; Guo, R.; Ki, J.S. 6.0K microarray reveals differential transcriptomic responses in the dinoflagellate Prorocentrum minimum exposed to polychlorinated biphenyl (PCB). Chemosphere 2018, 195, 398–409. [Google Scholar] [CrossRef]
- Guo, R.; Wang, H.; Suh, Y.S.; Ki, J.S. Transcriptomic profiles reveal the genome-wide responses of the harmful dinoflagellate Cochlodinium polykrikoides when exposed to the algicide copper sulfate. BMC Genom. 2016, 17, 29. [Google Scholar] [CrossRef]
- Luo, H.; Lin, X.; Li, L.; Lin, L.; Zhang, C.; Lin, S. Transcriptomic and physiological analyses of the dinoflagellate Karenia mikimotoi reveal non-alkaline phosphatase-based molecular machinery of ATP utilisation. Environ. Microbiol. 2017, 19, 4506–4518. [Google Scholar] [CrossRef]
- Cooper, J.T.; Sinclair, G.A.; Wawrik, B. Transcriptome Analysis of Scrippsiella trochoidea CCMP 3099 Reveals Physiological Changes Related to Nitrate Depletion. Front. Microbiol. 2016, 7, 639. [Google Scholar] [CrossRef]
- Price, D.C.; Farinholt, N.; Gates, C.; Shumaker, A.; Wagner, N.E.; Bienfang, P.; Bhattacharya, D. Analysis of Gambierdiscus transcriptome data supports ancient origins of mixotrophic pathways in dinoflagellates. Environ. Microbiol. 2016, 18, 4501–4510. [Google Scholar] [CrossRef]
- Hou, D.Y.; Mao, X.T.; Gu, S.; Li, H.Y.; Liu, J.S.; Yang, W.D. Systems-level analysis of metabolic mechanism following nitrogen limitation in benthic dinoflagellate Prorocentrum lima. Algal Res. 2018, 33, 389–398. [Google Scholar] [CrossRef]
- Baumgarten, S.; Bayer, T.; Aranda, M.; Liew, Y.J.; Voolstra, C.R. Integrating microRNA and mRNA expression profiling in Symbiodinium microadriaticum, a dinoflagellate symbiont of reef-building corals. BMC Genom. 2013, 14, 704. [Google Scholar] [CrossRef]
- Gierz, S.L.; Foret, S.; Leggat, W. Transcriptomic Analysis of Thermally Stressed Symbiodinium Reveals Differential Expression of Stress and Metabolism Genes. Front. Plant Sci. 2017, 8, 271. [Google Scholar] [CrossRef]
- Lin, S.J.; Yu, L.Y.; Zhang, H. Transcriptomic Responses to Thermal Stress and Varied Phosphorus Conditions in Fugacium kawagutii. Microorganisms 2019, 7, 96. [Google Scholar] [CrossRef]
- Jang, S.H.; Jeong, H.J.; Chon, J.K.; Lee, S.Y. De novo assembly and characterization of the transcriptome of the newly described dinoflagellate Ansanella granifera: Spotlight on flagellum-associated genes. Mar. Genom. 2017, 33, 47–55. [Google Scholar] [CrossRef]
- Ludewig-Klingner, A.K.; Michael, V.; Jarek, M.; Brinkmann, H.; Petersen, J. Distribution and Evolution of Peroxisomes in Alveolates (Apicomplexa, Dinoflagellates, Ciliates). Genome Biol. Evolut. 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Hastings, J.W. Genome-wide analysis of redox-regulated genes in a dinoflagellate. Gene 2003, 321, 73–81. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Hastings, J.W. Novel dinoflagellate clock-related genes identified through microarray analysis. J. Phycol. 2003, 39, 519–526. [Google Scholar] [CrossRef]
- Roy, S.; Letourneau, L.; Morse, D. Cold-induced cysts of the photosynthetic dinoflagellate Lingulodinium polyedrum have an arrested circadian bioluminescence rhythm and lower levels of protein phosphorylation. Plant Physiol. 2014, 164, 966–977. [Google Scholar] [CrossRef]
- Lowe, C.D.; Mello, L.V.; Samatar, N.; Martin, L.E.; Watts, P.C. The transcriptome of the novel dinoflagellate Oxyrrhis marina (Alveolata Dinophyceae) response to salinity examined by 454 sequencing. BMC Genom. 2011, 12, 519. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Hackett, J.D. Transcriptome analysis reveals nuclear-encoded proteins for the maintenance of temporary plastids in the dinoflagellate Dinophysis acuminata. BMC Genom. 2010, 11, 366. [Google Scholar] [CrossRef]
- Hackett, J.D.; Scheetz, T.E.; Yoon, H.S.; Soares, M.B.; Bonaldo, M.F.; Casavant, T.L.; Bhattacharya, D. Insights into a dinoflagellate genome through expressed sequence tag analysis. BMC Genom. 2005, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.J.; Zhang, H.; Zhuang, Y.Y.; Tran, B.; Gill, J. Spliced leader-based metatranscriptomic analyses lead to recognition of hidden genomic features in dinoflagellates. Proc. Natl. Acad. Sci. USA 2010, 107, 20033–20038. [Google Scholar] [CrossRef]
- Bhaud, Y.; Géraud, M.L.; Ausseil, J.; Soyer-Gobillard, M.O.; Moreau, H. Cyclic expression of a nuclear protein in a dinoflagellate. J. Eukaryot. Microbiol. 1999, 46, 259–267. [Google Scholar] [CrossRef]
- Bachvaroff, T.R.; Place, A.R. From stop to start: Tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidinium carterae. PLoS ONE 2008, 3, e2929. [Google Scholar] [CrossRef]
- Lidie, K.B.; van Dolah, F.M. Spliced leader RNA-mediated trans-splicing in a dinoflagellate, Karenia brevis. J. Eukaryot. Microbiol. 2007, 54, 427–435. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, Y.; Miranda, L.; Campbell, D.A.; Sturm, N.R.; Gaasterland, T.; Lin, S. Spliced leader RNA trans-splicing in dinoflagellates. Proc. Natl. Acad. Sci. USA 2007, 104, 4618–4623. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lin, X.; Shi, X.; Lin, L.; Luo, H.; Li, L.; Lin, S. Metatranscriptomic Signatures Associated with Phytoplankton Regime Shift From Diatom Dominance to a Dinoflagellate Bloom. Front. Microbiol. 2019, 10, 590. [Google Scholar] [CrossRef]
- Wase, N.; Black, P.N.; Stanley, B.A.; Dirusso, C.C. Integrated quantitative analysis of nitrogen stress response in Chlamydomonas reinhardtii using metabolite and protein profiling. J. Proteome Res. 2014, 13, 1373. [Google Scholar] [CrossRef]
- Roy, S.; Beauchemin, M.; Dagenais-Bellefeuille, S.; Letourneau, L.; Cappadocia, M.; Morse, D. The Lingulodinium circadian system lacks rhythmic changes in transcript abundance. BMC Biol. 2014, 12, 107. [Google Scholar] [CrossRef]
- Graham, R.; Graham, C.; McMullan, G. Microbial proteomics: A mass spectrometry primer for biologists. Microb. Cell Fact. 2007, 6, 26. [Google Scholar] [CrossRef]
- Geert, B.; Evy, V.; Arnold, D.l.; Liliane, S. Gel-based versus gel-free proteomics: A review. Comb. Chem. High Throughput Screen. 2005, 8, 669–677. [Google Scholar]
- Nie, L.; Wu, G.; Zhang, W. Statistical application and challenges in global gel-free proteomic analysis by mass spectrometry. Crit. Rev. Biotechnol. 2008, 28, 297. [Google Scholar] [CrossRef]
- Morse, D.; Tse, S.P.K.; Lo, S.C.L. Exploring dinoflagellate biology with high-throughput proteomics. Harmful Algae 2018, 75, 16–26. [Google Scholar] [CrossRef]
- Chan, L.L.; Hodgkiss, I.J.; Lam, K.S.; Wan, M.F.; Chou, H.N.; Lum, H.K. Use of two-dimensional gel electrophoresis to differentiate morphospecies of Alexandrium minutum, a paralytic shellfish poisoning toxin-producing dinoflagellate of harmful algal blooms. Proteomics 2005, 5, 1580–1593. [Google Scholar] [CrossRef]
- Sproles, A.E.; Oakley, C.A.; Matthews, J.L.; Peng, L.; Owen, J.G.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Proteomics quantifies protein expression changes in a model cnidarian colonised by a thermally tolerant but suboptimal symbiont. ISME J. 2019. [Google Scholar] [CrossRef]
- Cui, Y.D.; Zhang, H.; Lin, S.J. Enhancement of Non-photochemical Quenching as an Adaptive Strategy under Phosphorus Deprivation in the Dinoflagellate Karlodinium veneficum. Front. Microbiol. 2017, 8, 404. [Google Scholar] [CrossRef]
- Lee, W.F.; Ho, K.C.; Lo, C.L. Rapid identification of dinoflagellates using protein profiling with matrix-assisted laser desorption/ionization mass spectrometry. Harmful Algae 2008, 7, 551–559. [Google Scholar] [CrossRef]
- Chan, L.L.; Hodgkiss, I.J.; Lu, S.; Lo, S.C.L. Use of two-dimensional gel electrophoresis proteome reference maps of dinoflagellates for species recognition of causative agents of harmful algal blooms. Proteomics 2010, 4, 180–192. [Google Scholar] [CrossRef]
- Minge, M.A.; Shalchian-Tabrizi, K.; TRresen, O.K.; Takishita, K.; Probert, I.; Inagaki, Y.E.A. A phylogenetic mosaic plastid proteome and unusual plastid-targeting signals in the green-colored dinoflagellate Lepidodinium chlorophorum. BMC Evolut. Biol. 2010, 10, 191. [Google Scholar] [CrossRef]
- Lee, W.F.; Ho, K.C.; Mak, Y.L.; Lo, C.L. Authentication of the proteins expression profiles (PEPs) identification methodology in a bloom of Karenia digitata, the most damaging harmful algal bloom causative agent in the history of Hong Kong. Harmful Algae 2011, 12, 1–10. [Google Scholar] [CrossRef]
- Peng, S.E.; Luo, Y.J.; Huang, H.J.; Lee, I.T.; Hou, L.S.; Chen, W.N.U.; Fang, L.S.; Chen, C.S. Isolation of tissue layers in hermatypic corals by N -acetylcysteine: Morphological and proteomic examinations. Coral Reefs 2008, 27, 133–142. [Google Scholar] [CrossRef]
- Weston, A.J.; Dunlap, W.C.; Shick, J.M.; Klueter, A.; Iglic, K.; Vukelic, A.; Starcevic, A.; Ward, M.; Wells, M.L.; Trick, C.G.; et al. A profile of an endosymbiont-enriched fraction of the coral Stylophora pistillata reveals proteins relevant to microbial-host interactions. Mol. Cell. Proteomics 2012, 11, M111-015487. [Google Scholar] [CrossRef]
- Medrano, E.; Merselis, D.G.; Bellantuono, A.J.; Rodriguez-Lanetty, M. Proteomic Basis of Symbiosis: A Heterologous Partner Fails to Duplicate Homologous Colonization in a Novel Cnidarian—Symbiodiniaceae Mutualism. Front. Microbiol. 2019, 10, 1153. [Google Scholar] [CrossRef]
- Wang, D.Z.; Gao, Y.; Lin, L.; Hong, H.S. Comparative proteomic analysis reveals proteins putatively involved in toxin biosynthesis in the marine dinoflagellate Alexandrium catenella. Mar. Drugs 2013, 11, 213–232. [Google Scholar] [CrossRef]
- Weckwerth, W. Metabolomics in systems biology. Annu. Rev. Plant. Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp. Funct. Genom. 2001, 2, 155–168. [Google Scholar] [CrossRef]
- Kell, D.B. Metabolomics and systems biology: Making sense of the soup. Curr. Opin. Microbiol. 2004, 7, 296–307. [Google Scholar] [CrossRef]
- Cascante, M.; Marin, S. Metabolomics and fluxomics approaches. Essays Biochem. 2008, 45, 67–81. [Google Scholar] [CrossRef]
- Hillyer, K.E.; Dias, D.; Lutz, A.; Roessner, U.; Davy, S.K. (13)C metabolomics reveals widespread change in carbon fate during coral bleaching. Metabol. Off. J. Metabol. Soc. 2017, 14, 12. [Google Scholar]
- Matthews, J.L.; Oakley, C.A.; Lutz, A.; Hllyer, K.E.; Roessner, U.E.A. Partner switching and metabolic flux in a model cnidarian–dinoflagellate symbiosis. Proc. R. Soc. B Biol. Sci. 2018, 285, 20182336. [Google Scholar] [CrossRef]
- Weng, L.C.; Pasaribu, B.; Lin, I.P.; Tsai, C.H.; Chen, C.S.; Jiang, P.L. Nitrogen deprivation induces lipid droplet accumulation and alters fatty acid metabolism in symbiotic dinoflagellates isolated from Aiptasia pulchella. Sci. Rep. 2014, 4, 5777. [Google Scholar] [CrossRef]
- Klueter, A.; Crandall, J.B.; Archer, F.I.; Teece, M.A.; Coffroth, M.A. Taxonomic and environmental variation of metabolite profiles in marine dinoflagellates of the genus symbiodinium. Metabolites 2015, 5, 74–99. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y. Metabolite profiling of Breviolum minutum in response to acidification. Aquat. Toxicol. 2019, 213, 105215. [Google Scholar] [CrossRef]
- Sun, M.; Yang, Z.; Wawrik, B. Metabolomic Fingerprints of Individual Algal Cells Using the Single-Probe Mass Spectrometry Technique. Front. Plant Sci. 2018, 9, 571. [Google Scholar] [CrossRef]
- Ventura, P.; Jarrold, M.D.; Merle, P.L.; Barnay-Verdier, S.; Zamoum, T.; Rodolfo-Metalpa, R.; Calosi, P.; Furla, P. Resilience to ocean acidification: Decreased carbonic anhydrase activity in sea anemones under high pCO2 conditions. Mar. Ecol. Prog. Ser. 2016, 559, 257. [Google Scholar] [CrossRef]
- Silva, L.P.; Northen, T.R. Exometabolomics and MSI: Deconstructing how cells interact to transform their small molecule environment. Curr. Opin. Biotechnol. 2015, 34, 209–216. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef]
- Hegde, P.S.; White, I.R.; Debouck, C. Interplay of transcriptomics and proteomics. Curr. Opin. Biotechnol. 2003, 14, 647–651. [Google Scholar] [CrossRef]
- Mootha, V.K.; Jakob, B.; Olsen, J.V.; Majbrit, H.; Wisniewski, J.R.; Erich, S.; Bolouri, M.S.; Ray, H.N.; Smita, S.; Michael, K. Integrated analysis of protein composition, tissue diversity, and gene regulation in mouse mitochondria. Cell 2003, 115, 629–640. [Google Scholar] [CrossRef]
- Mootha, V.K.; Pierre, L.; Kathleen, M.; Jakob, B.; Michael, R.; Majbrit, H.; Terrye, D.; Amelie, V.; Robert, S.; Fenghao, X. Identification of a gene causing human cytochrome c oxidase deficiency by integrative genomics. Proc. Natl. Acad. Sci. USA 2003, 100, 605–610. [Google Scholar] [CrossRef]
- Orly, A.; Golub, G.H. Integrative analysis of genome-scale data by using pseudoinverse projection predicts novel correlation between DNA replication and RNA transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 16577–16582. [Google Scholar] [Green Version]
- Matthews, J.L.; Crowder, C.M.; Oakley, C.A.; Lutz, A.; Roessner, U.; Meyer, E.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Optimal nutrient exchange and immune responses operate in partner specificity in the cnidarian-dinoflagellate symbiosis. Proc. Natl. Acad. Sci. USA 2017, 114, 13194–13199. [Google Scholar] [CrossRef] [Green Version]
- Hayden, C.; Erika. Genome sequencing: The third generation. Nature 2009, 457, 768–769. [Google Scholar]
- Gong, W.; Browne, J.; Hall, N.; Schruth, D.; Paerl, H.; Marchetti, A. Molecular insights into a dinoflagellate bloom. ISME J. 2017, 11, 439–452. [Google Scholar] [CrossRef]
- Matamoros, O.; Fernando, M. Transient transformation of cultured photosynthetic dinoflagellates (Symbiodinium spp.) with plant-targeted vectors. Cienc. Mar. 2015, 41, 21–32. [Google Scholar] [CrossRef]
Figure 1.
Application of omics technologies for dinoflagellate biology research.

Figure 2.
Major metabolic pathways in F. kawagutii. Complete pathways for the mitochondrial tricarboxylic acid cycle (TCAcycle), fatty acid oxidation and the urea cycle, the chloroplastic Calvin cycle, dicarboxylic acid cycle (C4 cycle), crassulacean acid metabolism (CAM), fatty acid biosynthesis, peroxisomal fatty acid oxidation, glyoxylate cycle, and photorespiration are found in the F. kawagutii genome. FAS, fatty acid synthase; GOT1, aspartate aminotransferase; ppc, phosphoenolpyruvate carboxylase; MDH2, malate dehydrogenase; maeB, malic enzyme; ALT, alanine transaminase; ppdK, pyruvate, phosphate dikinase; rpe, L-ribulose phosphate epimerase; tktA, transketolase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PGK, phosphoglycerate kinase; rbcL, ribulose-bisphosphate carboxylase large chain; PRK, phosphoribulokinase; rpiA, ribose 5-phosphate isomerase A; glyK, glycerate kinase; ACSL, long-chain acyl-CoA synthetase; ACOX1, acyl-CoA oxidase; echA, enoyl-CoA hydratase; HADH, 3-hydroxyacyl-CoA dehydrogenase; ACAA1, acetyl-CoA acyltransferase; acnB, aconitate hydratase; aceA, isocitrate lyase; AceB, malate synthase; HAO, (S)-2-hydroxy-acid oxidase; katG, catalase; GGAT, glutamate--glyoxylate aminotransferase; GCSH, glycinSe cleavage system H protein; GyaR, glyoxylate reductase; AGXT, alanine-glyoxylate transaminase / serine-glyoxylate transaminase / serine-pyruvate transaminase; CS, citrate synthase; IDH, isocitrate dehydrogenase; OGDH, α-ketoglutarate dehydrogenase; LSC, Succinyl-CoA synthesase; SDH, succinate dehydrogetase; fumA, fumarase; CPS1, carbamoyl phosphate synthetase I; argF, ornithine carbamoyltransferase; argG, argininosuccinate synthase; argH, argininosuccinate lyase; NOS, NO synthetase; FA, fatty acid; OAA, Oxaloacetic acid; α-KG, α-ketoglutarate. (Schemed based on the study [38]).
Figure 2.
Major metabolic pathways in F. kawagutii. Complete pathways for the mitochondrial tricarboxylic acid cycle (TCAcycle), fatty acid oxidation and the urea cycle, the chloroplastic Calvin cycle, dicarboxylic acid cycle (C4 cycle), crassulacean acid metabolism (CAM), fatty acid biosynthesis, peroxisomal fatty acid oxidation, glyoxylate cycle, and photorespiration are found in the F. kawagutii genome. FAS, fatty acid synthase; GOT1, aspartate aminotransferase; ppc, phosphoenolpyruvate carboxylase; MDH2, malate dehydrogenase; maeB, malic enzyme; ALT, alanine transaminase; ppdK, pyruvate, phosphate dikinase; rpe, L-ribulose phosphate epimerase; tktA, transketolase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PGK, phosphoglycerate kinase; rbcL, ribulose-bisphosphate carboxylase large chain; PRK, phosphoribulokinase; rpiA, ribose 5-phosphate isomerase A; glyK, glycerate kinase; ACSL, long-chain acyl-CoA synthetase; ACOX1, acyl-CoA oxidase; echA, enoyl-CoA hydratase; HADH, 3-hydroxyacyl-CoA dehydrogenase; ACAA1, acetyl-CoA acyltransferase; acnB, aconitate hydratase; aceA, isocitrate lyase; AceB, malate synthase; HAO, (S)-2-hydroxy-acid oxidase; katG, catalase; GGAT, glutamate--glyoxylate aminotransferase; GCSH, glycinSe cleavage system H protein; GyaR, glyoxylate reductase; AGXT, alanine-glyoxylate transaminase / serine-glyoxylate transaminase / serine-pyruvate transaminase; CS, citrate synthase; IDH, isocitrate dehydrogenase; OGDH, α-ketoglutarate dehydrogenase; LSC, Succinyl-CoA synthesase; SDH, succinate dehydrogetase; fumA, fumarase; CPS1, carbamoyl phosphate synthetase I; argF, ornithine carbamoyltransferase; argG, argininosuccinate synthase; argH, argininosuccinate lyase; NOS, NO synthetase; FA, fatty acid; OAA, Oxaloacetic acid; α-KG, α-ketoglutarate. (Schemed based on the study [38]).

Figure 3.
Metabolic pathways in C. cohnii. Pathways associated with central carbohydrate, fatty acid and TAG biosynthesis, fatty acid oxidation, glyoxylate cycle, and nitrogen metabolic pathways. GK, glucokinase; G6PD, glucose-6-phosphate 1-dehydrogenase; G6PD2, 6-phosphogluconate dehydrogenase; PFK, 6-phosphofructokinase; FBA, fructosebisphosphate aldolase; GPD1, glycerol-3-phosphate dehydrogenase; GUT2, glycerol-kinase; PCK, phosphoenolpyruvate carboxykinase; PK, pyruvate kinase; ME, malic enzyme; MDE, malate dehydrogenase; ACL, ATP citrate lyase; ACC, acetyl-CoA carboxylase; PUFAS, polyunsaturated fatty acid synthase; FAS, fatty acid synthase; MCAT, malonyl-CoA: ACP transacylase; ACSL, long-chain acyl-CoA synthetase; DHA, docosahexaenoic acid; PUFA, polyunsaturated fatty acid; TAG, triacylglycerol. (Schemed based on the study [72]).
Figure 3.
Metabolic pathways in C. cohnii. Pathways associated with central carbohydrate, fatty acid and TAG biosynthesis, fatty acid oxidation, glyoxylate cycle, and nitrogen metabolic pathways. GK, glucokinase; G6PD, glucose-6-phosphate 1-dehydrogenase; G6PD2, 6-phosphogluconate dehydrogenase; PFK, 6-phosphofructokinase; FBA, fructosebisphosphate aldolase; GPD1, glycerol-3-phosphate dehydrogenase; GUT2, glycerol-kinase; PCK, phosphoenolpyruvate carboxykinase; PK, pyruvate kinase; ME, malic enzyme; MDE, malate dehydrogenase; ACL, ATP citrate lyase; ACC, acetyl-CoA carboxylase; PUFAS, polyunsaturated fatty acid synthase; FAS, fatty acid synthase; MCAT, malonyl-CoA: ACP transacylase; ACSL, long-chain acyl-CoA synthetase; DHA, docosahexaenoic acid; PUFA, polyunsaturated fatty acid; TAG, triacylglycerol. (Schemed based on the study [72]).


{kind=link}
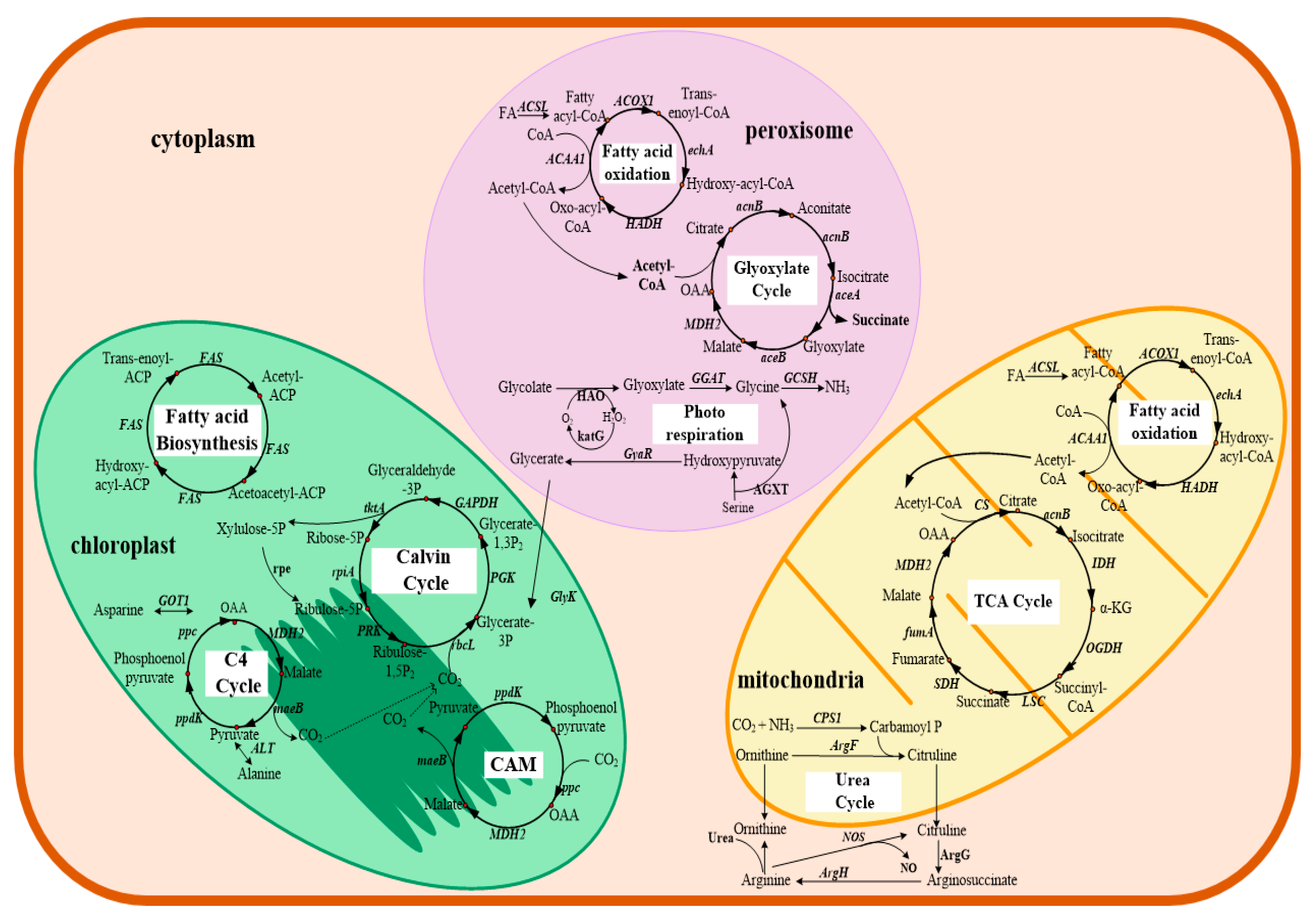{kind=link}
{kind=link}
Table 1.
Genomic summary of six dinoflagellates draft genomes.
| Breviolum minutum [38] | Fugacium kawagutii CCMP2468 [39] | Symbiodinium microadriaticum CCMP2467 [40] | Cladocopium goreaui SCF055-01 [7] | Fugacium kawagutii CCMP2468 [7] | Symbiodiniaceae [41] | Symbiodiniaceae [41] | ||
|---|---|---|---|---|---|---|---|---|
| Clade | B | F | A | C | F | A | C | |
| Total assembled length (bp) | 615,520,517 | 935,067,369 | 808,242,489 | 1,027,792,016 | 1,048,482,934 | 766,659,703 | 704,779,698 | |
| G+C content (%) | 43.6 | 43.97 | 50.51 | 44.83 | 45.72 | 49.9 | 43.0 | |
| Genes | Number of genes | 41,925 | 36,850 | 49,109 | 35,913 | 26,609 | 69,018 | 65,832 |
| Mean length of genes (bp) | 11,959 | 3788 | 12,898 | 6967 | 6507 | 8834 | 8192 | |
| Mean length of transcripts (bp) | 2067 | 1041 | 2377 | 1766 | 1736 | 1423 | 1479 | |
| Exons | No. of exons per gene | 19.6 | 4.1 | 21.8 | 10 | 8.7 | 13.38 | 11.27 |
| Mean length (bp) | 99.8 | 256 | 109.5 | 175.9 | 199.5 | 105 | 130 | |
| Total length (Mb) | 82.1 | 38.4 | 117.3 | 63.4 | 46.2 | 98.2 | 97.3 | |
| Introns | No. of genes with introns (%) | 95.3 | 64.1 | 98.2 | 92.9 | 94 | 83.4 | 80.3 |
| Mean length (bp) | 499 | 893 | 504.7 | 575.1 | 619.4 | 561 | 622 | |
| Total length (Mb) | 331.5 | 101.2 | 516.1 | 186.8 | 126.9 | 481.8 | 421.2 | |
| Intergenic regions | Mean length (bp) | 2064 | 17,888 | 3633 | 10,627 | 23,042 | 2008 | 2202 |
| platform | Roche 454 GS-FLX and Illumina (GAIIx) | Illumina HiSeq 2000 | Illumina HiSeq | Illumina HiSeq 2500 | Illumina HiSeq 2500 | Illumina (GAIIx) and Hiseq | Illumina (GAIIx) and Hiseq | |
| Bioproject ID | PRJDB732 | SRA148697 | PRJNA292355 | PRJEB20399 | PRJEB20399 | PRJDB3242 | PRJDB3243 |
GAIIx: Genome Analyzer IIx.
Table 2.
Transcriptomics studies of marine dinoflagellates.
| Species | Propose | Platform | Ref./year |
|---|---|---|---|
| Dinoflagellates responsible for HABs and toxic | |||
| Karenia brevis | To establish a database of K. brevis ESTs and identify conserved eukaryotic genes | microarray | [102] 2005 |
| Karenia brevis | To compare gene expression in response to light and dark | microarray | [103] 2007 |
| Karenia brevis | To investigate characterization and expression of nuclear-encoded polyketide synthases (PKSs) | microarray | [104] 2010 |
| Karenia brevis | To address transcriptional responses to nitrogen and phosphorus depletion and addition | microarray | [105] 2011 |
| Karenia brevis | To compare global transcriptome changes that accompany the entry and maintenance of stationary phase up to the onset of cell death. | microarray | [106] 2012 |
| Alexandrium catenella | To study the content and regulation of the dinoflagellate genome | MPSS | [44] 2006 |
| Alexandrium catenella CCMP1719 | To generate time-serial ESTs throughout a diel cycle during bloom | Roche 454 GS FLX | [107] 2015 |
| Alexandrium minutum | To compare gene expression in toxic versus non-toxic strains | Microarray | [47] 2010 |
| Alexandrium minutum | To determine transcriptional changes during the copepod-provoked induction of higher toxicity in A. minutum. | microarray | [108] 2011 |
| Alexandrium tamarense CCMP1598 | To investigate global transcriptional regulation under four different conditions, with xenic, nitrogen-limited, phosphorus- limited, and nutrient-replete | MPSS | [46] 2010 |
| Alexandrium monilatum CCMP3105 | To study transcriptional responses to limiting N and P conditions | Illumina HiSeq 2000 | [109] 2017 |
| Alexandrium ostenfeldii | To analyze gene composition, and structure and peculiarities of gene regulation | [110] 2011 | |
| Gambierdiscus polynesiensis | To reveal the mechanisms of CTX biosynthesis using transcriptomics | Roche 454 GS FLX | [54] 2014 |
| Alexandrium catenella | To construct an expressed sequence tag (EST) library from Alexandrium catenella | * | [43] 2008 |
| Alexandrium catenella | To determine the gene repertoire based on (NGS) technologies | Illumina Genome Analyzer. | [55] 2014 |
| Alexandrium catenella | To study the mechanism of PSTs synthesis using transcriptome profiles of a toxin-producing and its non-toxic mutant form | Illumina Hiseq 2000 | [56] 2014 |
| Alexandrium catenella (ACHK-T) | To study molecular mechanisms for PST biosynthesis using the transcriptome profiles of a toxin-producing dinoflagellates at different toxin biosynthesis stages within the cell cycle | Illumina Hiseq 2000 | [60] 2017 |
| Amphidinium carterae | To study de novo transcriptome for the identification of enzymes with biotechnological potential | Illumina HiSeq.1000 | [111] 2017 |
| Azadinium spinosum | Transcriptomic and genomic characterization of the toxigenic dinoflagellate with emphasis on polyketide synthase genes | Roche 454 GS FLX | [112] 2015 |
| Prorocentrum minimum CCMP 1329 | To identify genetic modules mediating the Jekyll and Hyde Interaction | Illumina HiSeq 2500 | [113] 2015 |
| Prorocentrum minimum D-127 | To evaluate genome-scale responses when exposed to polychlorinated biphenyl | microarray | [114] 2018 |
| Cochlodinium polykrikoides | To compare transcriptional responses to the algicide copper sulfate | Illumina HiSeq 2500 | [115] 2016 |
| Karenia mikimotoi C32-HK | To reveal non-alkaline phosphatase-based molecular machinery of ATP utilization | Roche 454 GS FLX | [116] 2017 |
| Dinoflagellates responsible for HABs and nontoxic | |||
| Scrippsiella trochoidea CCMP 3099 | To study the biochemical and physiological adaptations related to nutrient depletion | Illumina HiSeq 2000 | [117] 2016 |
| Dinoflagellate producing ciguatoxin | |||
| Gambierdiscus caribaeus | To trace the evolutionary history of C and N pathways in this phylum using transcriptome data | Illumina MiSeq | [118] 2016 |
| Dinoflagellates responsible for diarrheic shellfish poisoning | |||
| Prorocentrum lima CCMP 2579 | To compare the molecular and cellular responses to N limitation | Illumina HiSeq 2500 | [119] 2018 |
| Symbiotic dinoflagellates | |||
| Symbiodiniaceae (clade A) Symbiodiniaceae (clade B) | To construct an EST dataset for the genetic study of Symbiodiniaceae | Roche 454 GS FLX | [63] 2012 |
| Symbiodiniaceae Type D2 Symbiodiniaceae Type C3K | To compare transcriptional responses to thermal stress and the differences among physiologically susceptible and tolerant types | Illumina HiSeq | [64] 2014 |
| Symbiodinium microadriaticum clade A1, CCMP 2467 | To study the repertoire of endogenous smRNAs and to identify potential gene targets in dinoflagellates | Illumina HiSeq 2000 | [120] 2013 |
| Symbiodiniaceae SSB01 | To study transcriptional responses to immediate changes in light intensity when grown under autotrophic or mixotrophic conditions | Illumina HiSeq 2000 | [66] 2015 |
| Symbiodiniaceae type C1 | To study physiological and transcriptional responses to heat stress and to identify the gene related thermal response | Illumina HiSeq 2500 | [67] 2016 |
| Symbiodiniaceae type C1 | To study effects of viral infections to Symbiodiniaceae when heat-stressed | Illumina HiSeq 2500 | [70] 2017 |
| Symbiodiniaceae (clade F) | To study the transcriptional response of cellular mechanisms under future temperature conditions | Illumina HiSeq 2000 | [121] 2017 |
| Fugacium kawagutii CCMP2468 | To study transcriptional responses to thermal stress and varied phosphorus conditions | * | [122] 2019 |
| The fastest swimming dinoflagellates | |||
| Ansanella granifera | To study the structural and functional genes of dinoflagellate flagelle | Illumina HiSeq 2500 | [123] 2017 |
| high DHA yields dinoflagellate | |||
| Crypthecodinium cohnii ATCC 30556 | To compare transcriptional differences on a high lipid producing mutant with the wide-type strain | Illumina HiSeq 4000 | [12] 2017 |
| Crypthecodinium cohnii ATCC 30556 | To compare transcriptional difference following the growth course during fed-batch fermentation | Illumina HiSeq 2500 | [72] 2017 |
| Basal dinoflagellates | |||
| Perkinsus olseni | To study distribution and evolution of peroxisomes in the super ensemble Alveolata | MiSeq and HiSeq 2000 | [124] 2017 |
| Pyrocystis lunula | To study gene expression in Pyrocystis lunula in responsive to the addition of sodium nitrite and paraquat | microarray | [125] 2003 |
| Pyrocystis lunula | To analyze circadian regulation at transcriptional levels in Pyrocystis lunula | microarray | [126] 2003 |
| Lingulodinium polyedrum | To study molecular underpinnings of cold-induced cyst formation Sin the dinoflagellate L. polyedrum | Illumina HiSeq | [127] 2014 |
| Oxyrrhis marina | To study transcriptional responses to salinity | [128] 2011 | |
| Dinophysis acuminata | To determine whether the dinoflagellates contain nuclear-encoded genes for plastid function | Roche 454 GS FLX | [129] 2010 |
*, not mentioned; HABs: harmful algal blooms; ESTs: expressed sequence tags; MPSS: Massively Parallel Signature Sequencing; CTX: Ciguatoxins; NGS: next-generation sequencing; EDCs: Endocrine disrupting chemicals; PST: paralytic shellfish toxins.
Table 3.
Metabolomics studies of marine dinoflagellates.
| Species | Propose | Methods | Ref./year |
|---|---|---|---|
| Symbiodiniaceae (clade B) | To analysis fatty acid composition in cellular TAGs | GC-MS | [160] 2014 |
| Symbiodinium microadriaticum, Breviolum minutum, Breviolum psygmophilum, Durusdinium trenchii, | To study the metabolite profile of marine symbiotic dinoflagellates of Symbiodiniaceae | GC-MS | [161] 2015 |
| Symbiodiniaceae | To compare widespread change in carbon fate during coral bleaching | 13C labelling coupled to GC-MS | [158] 2017 |
| Symbiodiniaceae | To map carbon fate during bleaching | 13C labelling coupled to GC-MS | [69] 2017 |
| Breviolum minutum | to determine metabolomic changes of Breviolum minutum in acidification condition, and explore the possible mechanisms | LC-MS/MS | [162] 2019 |
| Crypthecodinium cohnii ATCC 30772 | To study mechanism of antioxidant butylated hydroxyanisole on lipid accumulation | LC-MS and GC-MS | [13] 2014 |
| Crypthecodinium cohnii ATCC 30556 | To reveal mechanisms related to glucose tolerance of C. cohnii through adaptive laboratory evolution | GC-MS | [11] 2017 |
| Crypthecodinium cohnii ATCC 30556 | To study physiological metabolism of C. cohnii for increased DHA production | GC-MS | [72] 2017 |
| Crypthecodinium cohnii ATCC 30556 | To gain understanding of the lipid metabolism and mechanism for the positive effects of modulator ethanolamine | 13C labelling coupled to GC-MS | [73] 2018 |
| Crypthecodinium cohnii ATCC 30556 | To study metabolic responses to different dissolved oxygen levels during DHA fermentation | GC-MS | [74] 2018 |
| Crypthecodinium cohnii ATCC 30556 | To compare molecular mechanisms of lipid accumulation in different strains | LC-MS and GC-MS | [75] 2018 |
| Crypthecodinium cohnii M-1-2 | To determine the metabolic changes under different nitrogen feeding conditions | GC-MS | [76] 2018 |
| Crypthecodinium cohnii ATCC 30556 | To comparative analyze C. cohnii mutants obtained from laboratory evolution | LC-MS and GC-MS | [15] 2018 |
| Scrippsiella trochoidea CCMP 3099 | To development single-cell metabolomics methodologies for small protists such as marine dinoflagellates | ‘Single-probe’ MS | [163] 2018 |
| Ostreopsis cf. ovata | To determine allelopathic interactions between the benthic toxic dinoflagellate Ostreopsis cf. ovata and a co-occurring diatom | LC-MS | [61] 2018 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bi, Y.; Wang, F.; Zhang, W. Omics Analysis for Dinoflagellates Biology Research. Microorganisms 2019, 7, 288. https://doi.org/10.3390/microorganisms7090288
AMA Style
Bi Y, Wang F, Zhang W. Omics Analysis for Dinoflagellates Biology Research. Microorganisms. 2019; 7(9):288. https://doi.org/10.3390/microorganisms7090288
Chicago/Turabian StyleBi, Yali, Fangzhong Wang, and Weiwen Zhang. 2019. "Omics Analysis for Dinoflagellates Biology Research" Microorganisms 7, no. 9: 288. https://doi.org/10.3390/microorganisms7090288
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.