Autophagic Proteome in Two Saccharomyces cerevisiae Strains during Second Fermentation for Sparkling Wine Elaboration


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Second Fermentation Conditions
2.2. Protein Extraction, Identification, and Quantification
2.3. Confidence Criteria and Statistical Analysis
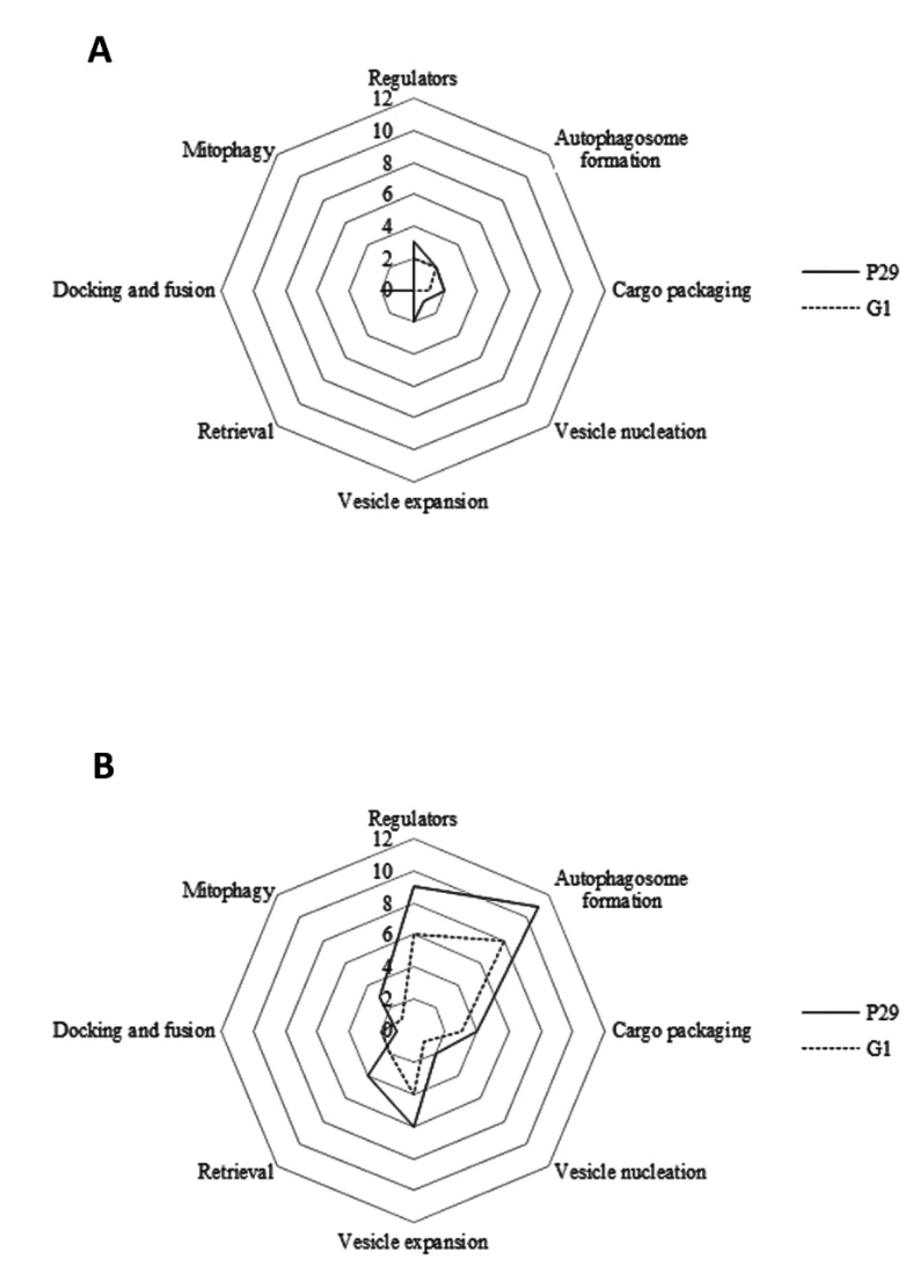
3. Results and Discussion
3.1. Regulation of Induction
3.2. Autophagosome-Generating Machinery
3.3. Cargo Packaging
3.4. Vesicle Nucleation
3.5. Vesicle Expansion and Completion
3.6. Retrieval
3.7. Docking and Fusion
3.8. Vesicle Breakdown and Permease Efflux
3.9. Mitophagy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torresi, S.; Frangipane, M.T.; Anelli, G. Biotechnologies in sparkling wine production. Interesting approaches for quality improvement: A review. Food Chem. 2011, 129, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, R.; García-Martínez, T.; Puig-Pujol, A.; Mauricio, J.C.; Moreno, J. Changes in sparkling wine aroma during the second fermentation under CO2 pressure in sealed bottle. Food Chem. 2017, 237, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine–a review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Kemp, B.; Alexandre, H.; Robillard, B.; Marchal, R. Effect of production phase on bottle-fermented sparkling wine quality. J. Agr. Food Chem. 2015, 63, 19–38. [Google Scholar] [CrossRef]
- Cebollero, E.; Carrascosa, A.V.; Gonzalez, R. Evidence for yeast autophagy during simulation of sparkling wine aging: A reappraisal of the mechanism of yeast autolysis in wine. Biotechnol. Prog. 2005, 21, 614–616. [Google Scholar] [CrossRef]
- Cebollero, E.; Gonzalez, R. Induction of autophagy by second-fermentation yeasts during elaboration of sparkling wines. Appl. Environ. Microbiol. 2006, 72, 4121–4127. [Google Scholar] [CrossRef] [Green Version]
- Nakatogawa, H.; Suzuki, K.; Kamada, Y.; Ohsumi, Y. Dynamics and diversity in autophagy mechanisms: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2009, 10, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Torggler, R.; Papinski, D.; Kraft, C. Assays to monitor autophagy in Saccharomyces cerevisiae. Cells 2017, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Reggiori, F.; Klionsky, D.J. Autophagic processes in yeast: Mechanism, machinery and regulation. Genetics 2013, 194, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Lynch-Day, M.A.; Klionsky, D.J. The Cvt pathway as a model for selective autophagy. FEBS Lett. 2010, 584, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Moreau, K.; Luo, S.; Rubinsztein, D.C. Cytoprotective roles for autophagy. Curr. Opin. Cell Biol. 2010, 22, 206–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero, E.; Tronchoni, J.; Morales, P.; Gonzalez, R. Autophagy is required for sulfur dioxide tolerance in Saccharomyces cerevisiae. Microb. Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Martinez-Rodriguez, A.J.; Carrascosa, A.V. Yeast autolytic mutants potentially useful for sparkling wine production. Int. J. Food Microbiol. 2003, 84, 21–26. [Google Scholar] [CrossRef]
- Cebollero, E.; Martinez-Rodriguez, A.; Carrascosa, A.V.; Gonzalez, R. Overexpression of csc1-1. A plausible strategy to obtain wine yeast strains undergoing accelerated autolysis. FEMS Microbiol. Lett. 2005, 246, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tabera, L.; Muñoz, R.; Gonzalez, R. Deletion of BCY1 from the Saccharomyces cerevisiae genome is semidominant and induces autolytic phenotypes suitable for improvement of sparkling wines. Appl. Environ. Microbiol. 2006, 72, 2351–2358. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, Y.P.; Carrascosa, A.V.; González, R.; Polo, M.C.; Martínez-Rodríguez, A.J. Effect of accelerated autolysis of yeast on the composition and foaming properties of sparkling wines elaborated by a champenoise method. J. Agr. Food Chem. 2005, 53, 7232–7237. [Google Scholar] [CrossRef]
- Legras, J.L.; Moreno-García, J.; Zara, S.; Zara, G.; García-Martínez, T.; Mauricio, J.C.; Mannazzu, I.; Coi, A.L.; Zeidan, M.B.; Dequin, S.; et al. Flor yeast: New perspectives beyond wine aging. Front. Microbiol. 2016, 7, 503. [Google Scholar] [CrossRef] [Green Version]
- Porras-Agüera, J.A.; Moreno-García, J.; Mauricio, J.C.; Moreno, J.; García-Martínez, T. First Proteomic Approach to Identify Cell Death Biomarkers in Wine Yeasts during Sparkling Wine Production. Microorganisms 2019, 7, 542. [Google Scholar] [CrossRef] [Green Version]
- Moreno-García, J.; García-Martínez, T.; Moreno, J.; Mauricio, J.C. Proteins involved in flor yeast carbon metabolism under biofilm formation conditions. Food Microbiol. 2015, 46, 25–33. [Google Scholar] [CrossRef]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- Dasari, S.; Chambers, M.C.; Slebos, R.J.; Zimmerman, L.J.; Ham, A.J.L.; Tabb, D.L. TagRecon: High-throughput mutation identification through sequence tagging. J. Proteome Res. 2010, 9, 1716–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turetschek, R.; Lyon, D.; Desalegn, G.; Kaul, H.P.; Wienkoop, S. A proteomic workflow using high-throughput de novo sequencing towards complementation of genome information for improved comparative crop science. In Proteomics in Systems Biology; Humana Press: New York, NY, USA, 2016; pp. 233–243. [Google Scholar]
- Hughes, H.; Stephens, D.J. Assembly, organization, and function of the COPII coat. Histochem. Cell Biol. 2008, 129, 129–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchaud, N.; Péli-Gulli, M.P. SEACing the GAP that nEGOCiates TORC1 activation: Evolutionary conservation of Rag GTPase regulation. Cell Cycle 2013, 12, 2948–2952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorimitsu, T.; Zaman, S.; Broach, J.R.; Klionsky, D.J. Protein kinase A and Sch9 cooperatively regulate induction of autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 4180–4189. [Google Scholar] [CrossRef] [Green Version]
- Yeasmin, A.M.; Waliullah, T.M.; Kondo, A.; Kaneko, A.; Koike, N.; Ushimaru, T. Orchestrated Action of PP2A Antagonizes Atg13 Phosphorylation and Promotes Autophagy after the Inactivation of TORC1. PLoS ONE 2016, 11, e0166636. [Google Scholar] [CrossRef] [Green Version]
- Farhan, H.; Kundu, M.; Ferro-Novick, S. The link between autophagy and secretion: A story of multitasking proteins. Mol. Biol. Cell 2017, 28, 1161–1164. [Google Scholar] [CrossRef]
- Krick, R.; Bremer, S.; Welter, E.; Schlotterhose, P.; Muehe, Y.; Eskelinen, E.; Thumm, M. Cdc48/p97 and Shp1/p47 regulate autophagosome biogenesis in concert with ubiquitin-like Atg8. J. Cell Biol. 2010, 190, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Nair, U.; Klionsky, D.J. Atg8 controls phagophore expansion during autophagosome formation. Mol. Biol. Cell 2008, 19, 3290–3298. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.P.; Scott, S.V.; Kim, J.; Klionsky, D.J. The itinerary of a vesicle component, Aut7p/Cvt5p, terminates in the yeast vacuole via the autophagy/Cvt pathways. J. Biol. Chem. 2000, 275, 5845–5851. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, N.; Hamasaki, M.; Yokota, S.; Suzuki, K.; Kamada, Y.; Kihara, A.; Yoshimori, T.; Noda, T.; Ohsumi, Y. Autophagosome requires specific early Sec proteins for its formation and NSF/SNARE for vacuolar fusion. Mol. Biol. Cell 2001, 12, 3690–3702. [Google Scholar] [CrossRef]
- Penacho, V.; Valero, E.; Gonzalez, R. Transcription profiling of sparkling wine second fermentation. Int. J. Food Microbiol. 2012, 153, 176–182. [Google Scholar] [CrossRef]
- Suzuki, K.; Kubota, Y.; Sekito, T.; Ohsumi, Y. Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells 2007, 12, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Akioka, M.; Kondo-Kakuta, C.; Yamamoto, H.; Ohsumi, Y. Fine mapping of autophagy-related proteins during autophagosome formation in Saccharomyces cerevisiae. J. Cell Sci. 2013, 126, 2534–2544. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Kamada, Y.; Ohsumi, Y. Studies of cargo delivery to the vacuole mediated by autophagosomes in Saccharomyces cerevisiae. Dev. Cell 2002, 3, 815–824. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, M.U.; Klionsky, D.J. Vacuolar Localization of Oligomeric α-Mannosidase Requires the Cytoplasm to Vacuole Targeting and Autophagy Pathway Components in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 20491–20498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onodera, J.; Ohsumi, Y. Ald6p is a preferred target for autophagy in yeast, Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 16071–16076. [Google Scholar] [CrossRef] [Green Version]
- Aranda, A.; del Olmo, M. Response to acetaldehyde stress in the yeast Saccharomyces cerevisiae involves a strain-dependent regulation of several ALD genes and is mediated by the general stress response pathway. Yeast 2003, 20, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Khalfan, W.A.; Klionsky, D.J. Molecular machinery required for autophagy and the cytoplasm to vacuole targeting (Cvt) pathway in S. cerevisiae. Curr. Opin. Cell Biol. 2002, 14, 468–475. [Google Scholar] [CrossRef]
- Kihara, A.; Noda, T.; Ishihara, N.; Ohsumi, Y. Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae. J. Cell Biol. 2001, 152, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Geng, J.; Nair, U.; Yasumura-Yorimitsu, K.; Klionsky, D.J. Post-Golgi Sec proteins are required for autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2010, 21, 2257–2269. [Google Scholar] [CrossRef] [Green Version]
- Lipatova, Z.; Belogortseva, N.; Zhang, X.Q.; Kim, J.; Taussig, D.; Segev, N. Regulation of selective autophagy onset by a Ypt/Rab GTPase module. Proc. Natl. Acad. Sci. USA 2012, 109, 6981–6986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, W.L.; Legakis, J.E.; Nair, U.; Klionsky, D.J. Atg27 is required for autophagy-dependent cycling of Atg9. Mol. Biol. Cell 2007, 18, 581–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, W.L.; Klionsky, D.J. Atg27 is a second transmembrane cycling protein. Autophagy 2007, 3, 254–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, H.; Meiling-Wesse, K.; Epple, U.D.; Thumm, M. Autophagy and the cytoplasm to vacuole targeting pathway both require Aut10p. FEBS Lett. 2001, 508, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Levine, B.; Klionsky, D.J. Development by self-digestion: Molecular mechanisms and biological functions of autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- McNew, J.A.; Sogaard, M.; Lampen, N.M.; Machida, S.; Ye, R.R.; Lacomis, L.; Tempst, P.; Rothman, J.E.; Söllner, T.H. Ykt6p, a prenylated SNARE essential for endoplasmic reticulum-Golgi transport. J. Biol. Chem. 1997, 272, 17776–17783. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Gomez, T.S.; Sackey, B.K.; Billadeau, D.D.; Burd, C.G. Rab GTPase regulation of retromer-mediated cargo export during endosome maturation. Mol. Biol. Cell 2012, 23, 2505–2515. [Google Scholar] [CrossRef]
- Seals, D.F.; Eitzen, G.; Margolis, N.; Wickner, W.T.; Price, A. A Ypt/Rab effector complex containing the Sec1 homolog Vps33p is required for homotypic vacuole fusion. Proc. Natl. Acad. Sci. USA 2000, 97, 9402–9407. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Kanki, T. Mechanisms and physiological roles of mitophagy in yeast. Mol. Cells 2018, 41, 35. [Google Scholar] [CrossRef]
- Kanki, T.; Klionsky, D.J. Mitophagy in yeast occurs through a selective mechanism. J. Biol. Chem. 2008, 283, 32386–32393. [Google Scholar] [CrossRef] [Green Version]
- Kanki, T.; Wang, K.; Cao, Y.; Baba, M.; Klionsky, D.J. Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 2009, 17, 98–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemasters, J.J. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res. 2005, 8, 3–5. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Protein Frequency in Saccharomyces cerevisiae | PC T1 | NPCT1 | PCT2 | NPCT2 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Protein Frequency | Protein Frequency | Protein Frequency | Protein Frequency | ||||||
| P29 | G1 | P29 | G1 | P29 | G1 | P29 | G1 | ||
| Total proteins | 94 out of 6721, 1.4% | 11 out of 594, 1.85% | 7 out of 568, 1.23% | 29 out of 1517, 1.91% | 19 out of 1000, 1.90% | 4 out of 419, 1.33% | − | 4 out of 392, 1.86% | 2 out of 218, 0.91% |
| Regulation of induction | 26 out of 6721, 0.39% | 3 out of 594, 0.51% | 2 out of 568, 0.35% | 9 out of 1517, 0.59% | 6 out of 1000, 0.60% | 1 out of 419, 0.24% | - | - | - |
| Autophagosome-generating machinery | 24 out of 6721, 0.36% | 2 out of 594, 0.34% | 2 out of 568, 0.35% | 10 out of 1517, 0.66% | 8 out of 1000, 0.80% | 1 out of 419, 0.24% | - | 2 out of 392, 0.51% | - |
| Cargo packaging | 8 out of 6721, 0.12% | 1 out of 594, 0.17% | 1 out of 568, 0.18% | 4 out of 1517, 0.26% | 3 out of 1000, 0.30% | 2 out of 419, 0.48% | − | 2 out of 392, 0.51% | 2 out of 218, 0.92% |
| Vesicle nucleation | 5 out of 6721, 0.07% | 1 out of 594, 0.17% | - | 2 out of 1517, 0.13% | 1 out of 1000, 0.10% | - | - | - | - |
| Vesicle expansion | 26 out of 6721, 0.39% | 2 out of 594, 0.34% | 1 out of 568, 0.18% | 6 out of 1517, 0.40% | 4 out of 1000, 0.40% | - | - | - | - |
| Retrieval | 7 out of 6721, 0.10% | - | - | 4 out of 1517, 0.26% | 2 out of 1000, 0.20% | - | - | - | - |
| Docking and fusion | 14 out of 6721, 0.21% | 2 out of 594, 0.34% | 1 out of 568, 0.18% | 1 out of 1517, 0.07% | 1 out of 1000, 0.10% | - | - | - | - |
| Vesicle breakdown | 1 out of 6721, 0.01% | - | - | - | - | - | - | - | - |
| Permease efflux | 1 out of 6721, 0.01% | - | - | - | - | - | - | - | - |
| Mitophagy | 10 out of 6721, 0.15% | − | − | 3 out of 1517, 0.20% | 1 out of 1000, 0.10% | − | − | 1 out of 392, 0.26% | − |
| Yeast Strains | S. cerevisiae P29 | S. cerevisiae G1 | ||
|---|---|---|---|---|
| Sampling times | T1 | T2 | T1 | T2 |
| Regulators/inductors | - | Bcy1p (Specific, 0.04) | Sec13p (3.21) | - |
| Autophagosome-generating machinery | - | Shp1p (Specific, 0.05) | Sec18p (3.40), Shp1p (3.08) | - |
| Cargo packaging | - | Ams1p (Specific, 0.02) | Ape1p (2.90) | - |
| Vesicle nucleation | Vps15p (2.27) | - | - | - |
| Vescicle expansion | Sec2p (3.43) | - | - | - |
| Retrieval | - | - | - | - |
| Docking and fusion | Ykt6p (Specific, 0.10), Ypt7p (Specific, 0.11) | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porras-Agüera, J.A.; Moreno-García, J.; González-Jiménez, M.d.C.; Mauricio, J.C.; Moreno, J.; García-Martínez, T. Autophagic Proteome in Two Saccharomyces cerevisiae Strains during Second Fermentation for Sparkling Wine Elaboration. Microorganisms 2020, 8, 523. https://doi.org/10.3390/microorganisms8040523
Porras-Agüera JA, Moreno-García J, González-Jiménez MdC, Mauricio JC, Moreno J, García-Martínez T. Autophagic Proteome in Two Saccharomyces cerevisiae Strains during Second Fermentation for Sparkling Wine Elaboration. Microorganisms. 2020; 8(4):523. https://doi.org/10.3390/microorganisms8040523
Chicago/Turabian StylePorras-Agüera, Juan Antonio, Jaime Moreno-García, María del Carmen González-Jiménez, Juan Carlos Mauricio, Juan Moreno, and Teresa García-Martínez. 2020. "Autophagic Proteome in Two Saccharomyces cerevisiae Strains during Second Fermentation for Sparkling Wine Elaboration" Microorganisms 8, no. 4: 523. https://doi.org/10.3390/microorganisms8040523