Human Cytomegalovirus Infection Suppresses CD34+ Progenitor Cell Engraftment in Humanized Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. CD34+ HPC Isolation
2.2. CD34+ HPC Colony Formation Assay
2.3. Mice
2.4. Generation of HuBLT Mice
2.5. Analysis of Human Cell Engraftment
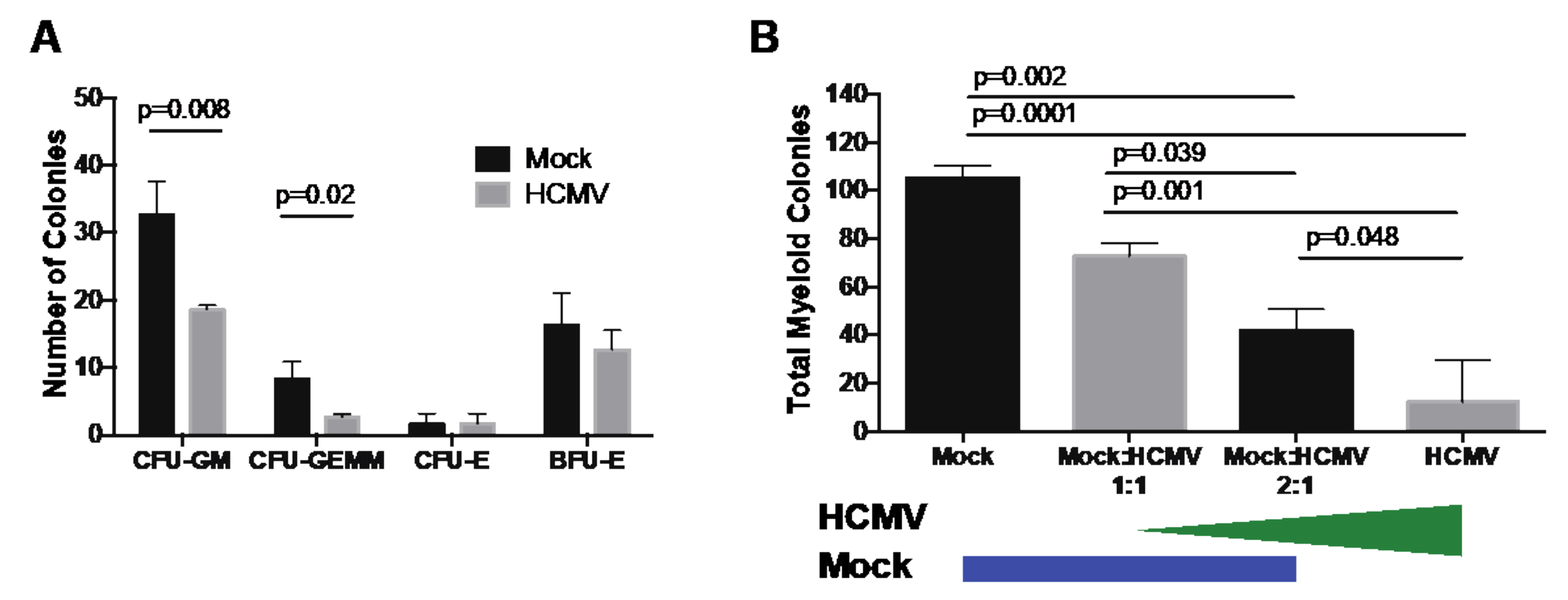
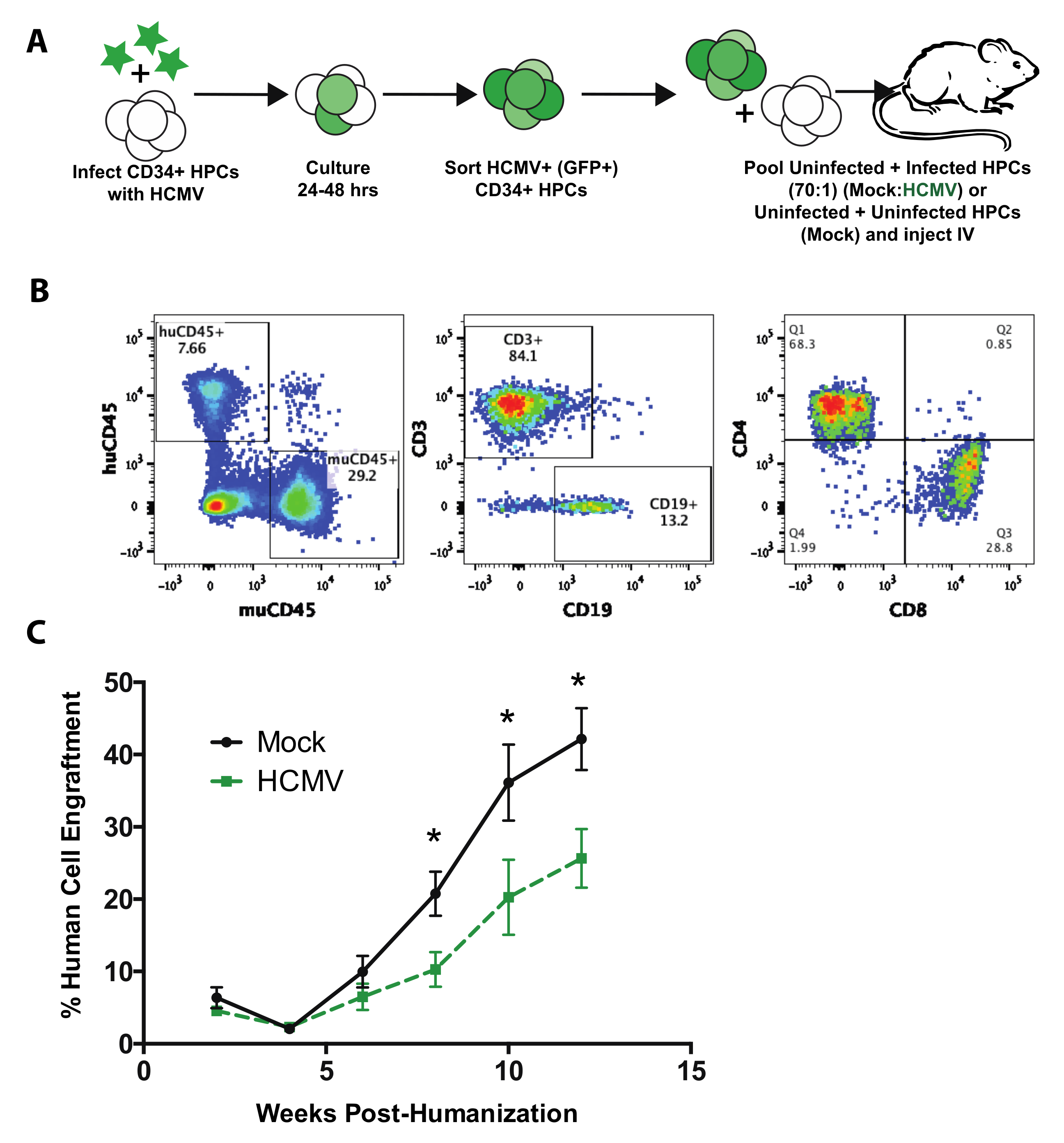
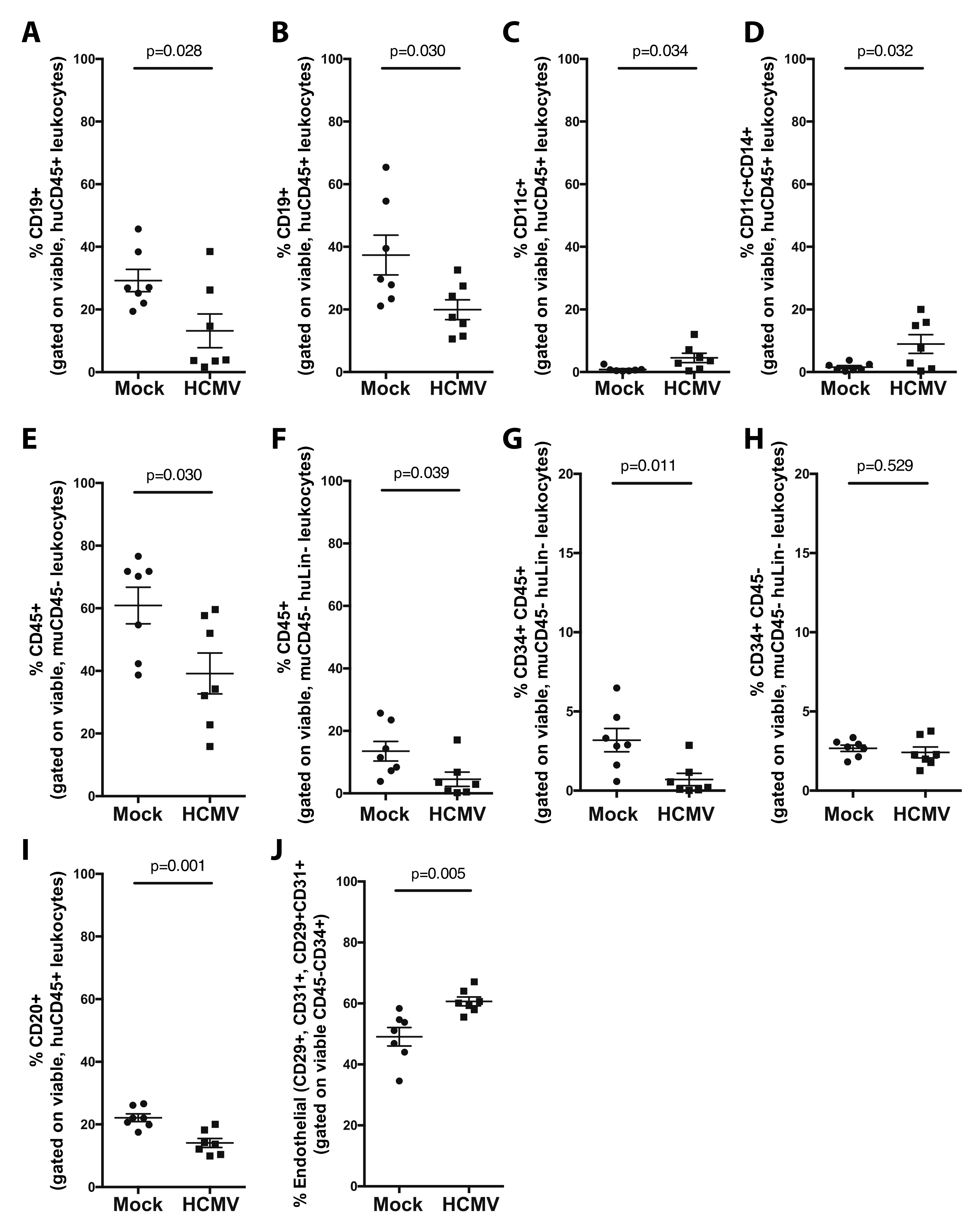
3. Results and Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Boeckh, M.; Geballe, A.P. Cytomegalovirus: pathogen, paradigm, and puzzle. J. Clin. Investig. 2011, 121, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Rakusan, T.A.; Juneja, H.S.; Fleischmann, W.R. Inhibition of hemopoietic colony formation by human cytomegalovirus in vitro. J. Infect. Dis. 1989, 159, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Maciejewski, J.P.; Bruening, E.E.; Donahue, R.E.; Mocarski, E.S.; Young, N.S.; St Jeor, S.C. Infection of hematopoietic progenitor cells by human cytomegalovirus. Blood 1992, 80, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Goodrum, F.; Jordan, C.T.; Terhune, S.S.; High, K.; Shenk, T. Differential outcomes of human cytomegalovirus infection in primitive hematopoietic cell subpopulations. Blood 2004, 104, 687–695. [Google Scholar] [CrossRef]
- Sing, G.K.; Ruscetti, F.W. Preferential suppression of myelopoiesis in normal human bone marrow cells after in vitro challenge with human cytomegalovirus. Blood 1990, 75, 1965–1973. [Google Scholar] [CrossRef]
- Hancock, M.H.; Crawford, L.B.; Pham, A.H.; Mitchell, J.; Struthers, H.M.; Yurochko, A.D.; Caposio, P.; Nelson, J.A. Human Cytomegalovirus miRNAs Regulate TGF-beta to Mediate Myelosuppression while Maintaining Viral Latency in CD34(+) Hematopoietic Progenitor Cells. Cell Host Microbe 2020, 27, 104–114. [Google Scholar] [CrossRef]
- Smith, M.S.; Goldman, D.C.; Bailey, A.S.; Pfaffle, D.L.; Kreklywich, C.N.; Spencer, D.B.; Othieno, F.A.; Streblow, D.N.; Garcia, J.V.; Fleming, W.H.; et al. Granulocyte-colony stimulating factor reactivates human cytomegalovirus in a latently infected humanized mouse model. Cell Host Microbe 2010, 8, 284–291. [Google Scholar] [CrossRef]
- Anderson, D.; DeFor, T.; Burns, L.; McGlave, P.; Miller, J.; Wagner, J.; Weisdorf, D. A comparison of related donor peripheral blood and bone marrow transplants: Importance of late-onset chronic graft-versus-host disease and infections. Biol. Blood Marrow Transplant. 2003, 9, 52–59. [Google Scholar] [CrossRef]
- Blaise, D.; Kuentz, M.; Fortanier, C.; Bourhis, J.H.; Milpied, N.; Sutton, L.; Jouet, J.-P.; Attal, M.; Bordigoni, P.; Cahn, J.-Y.; et al. Randomized Trial of Bone Marrow Versus Lenograstim-Primed Blood Cell Allogeneic Transplantation in Patients With Early-Stage Leukemia: A Report From the Société Française de Greffe de Moelle. J. Clin. Oncol. 2000, 18, 537. [Google Scholar] [CrossRef]
- Champlin, R.E.; Schmitz, N.; Horowitz, M.M.; Chapuis, B.; Chopra, R.; Cornelissen, J.J.; Gale, R.P.; Goldman, J.M.; Loberiza, F.R.J.; Hertenstein, B.; et al. Blood stem cells compared with bone marrow as a source of hematopoietic cells for allogeneic transplantation. IBMTR Histocompatibility and Stem Cell Sources Working Committee and the European Group for Blood and Marrow Transplantation (EBMT). Blood 2000, 95, 3702–3709. [Google Scholar]
- Goodrum, F. Human Cytomegalovirus Latency: Approaching the Gordian Knot. Annu. Rev. Virol. 2016, 3, 333–357. [Google Scholar] [CrossRef] [PubMed]
- Crawford, L.B.; Tempel, R.; Streblow, D.N.; Kreklywich, C.; Smith, P.; Picker, L.J.; Nelson, J.A.; Caposio, P. Human Cytomegalovirus Induces Cellular and Humoral Virus-specific Immune Responses in Humanized BLT Mice. Sci. Rep. 2017, 7, 937. [Google Scholar] [CrossRef] [PubMed]
- Covassin, L.; Jangalwe, S.; Jouvet, N.; Laning, J.; Burzenski, L.; Shultz, L.D.; Brehm, M.A. Human immune system development and survival of non-obese diabetic (NOD)-scid IL2rgamma(null) (NSG) mice engrafted with human thymus and autologous haematopoietic stem cells. Clin. Exp. Immunol. 2013, 174, 372–388. [Google Scholar] [CrossRef] [PubMed]
- Wahl, A.; De, C.; Abad Fernandez, M.; Lenarcic, E.M.; Xu, Y.; Cockrell, A.S.; Cleary, R.A.; Johnson, C.E.; Schramm, N.J.; Rank, L.M.; et al. Precision mouse models with expanded tropism for human pathogens. Nature Biotechnol. 2019. [Google Scholar] [CrossRef]
- Page, K.M.; Zhang, L.; Mendizabal, A.; Wease, S.; Carter, S.; Gentry, T.; Balber, A.E.; Kurtzberg, J. Total Colony-Forming Units Are a Strong, Independent Predictor of Neutrophil and Platelet Engraftment after Unrelated Umbilical Cord Blood Transplantation: A Single-Center Analysis of 435 Cord Blood Transplants. Biol. Blood Marrow Transplant. 2011, 17, 1362–1374. [Google Scholar] [CrossRef]
- Prasad, V.K.; Mendizabal, A.; Parikh, S.H.; Szabolcs, P.; Driscoll, T.A.; Page, K.; Lakshminarayanan, S.; Allison, J.; Wood, S.; Semmel, D.; et al. Unrelated donor umbilical cord blood transplantation for inherited metabolic disorders in 159 pediatric patients from a single center: influence of cellular composition of the graft on transplantation outcomes. Blood 2008, 112, 2979–2989. [Google Scholar] [CrossRef]
- Slobedman, B.; Mocarski, E.S. Quantitative analysis of latent human cytomegalovirus. J. Virol. 1999, 73, 4806–4812. [Google Scholar] [CrossRef]
- Sindre, H.; Tjoonnfjord, G.E.; Rollag, H.; Ranneberg-Nilsen, T.; Veiby, O.P.; Beck, S.; Degre, M.; Hestdal, K. Human cytomegalovirus suppression of and latency in early hematopoietic progenitor cells. Blood 1996, 88, 4526–4533. [Google Scholar] [CrossRef]
- Goodrum, F.D.; Jordan, C.T.; High, K.; Shenk, T. Human cytomegalovirus gene expression during infection of primary hematopoietic progenitor cells: a model for latency. Proc. Natl. Acad. Sci. USA 2002, 99, 16255–16260. [Google Scholar] [CrossRef]
- Mendelson, M.; Monard, S.; Sissons, P.; Sinclair, J. Detection of endogenous human cytomegalovirus in CD34+ bone marrow progenitors. J. Gen. Virol. 1996, 77, 3099–3102. [Google Scholar] [CrossRef]
- Von Laer, D.; Meyer-Koenig, U.; Serr, A.; Finke, J.; Kanz, L.; Fauser, A.A.; Neumann-Haefelin, D.; Brugger, W.; Hufert, F.T. Detection of cytomegalovirus DNA in CD34+ cells from blood and bone marrow. Blood 1995, 86, 4086–4090. [Google Scholar] [CrossRef] [PubMed]
- Verdonck, L.; de Gast, G.; van Heugten, H.; Nieuwenhuis, H.; Dekker, A. Cytomegalovirus infection causes delayed platelet recovery after bone marrow transplantation. Blood 1991, 78, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Torok-Storb, B.; Simmons, P.; Khaira, D.; Stachel, D.; Myerson, D. Cytomegalovirus and marrow function. Ann. Hematol. 1992, 64, A128–A131. [Google Scholar] [CrossRef] [PubMed]
- Florescu, D.F.; Kalil, A.C.; Qiu, F.; Schmidt, C.M.; Sandkovsky, U. What Is the Impact of Hypogammaglobulinemia on the Rate of Infections and Survival in Solid Organ Transplantation? A Meta-Analysis. Am. J. Transplant. 2013, 13, 2601–2610. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, M.A.; Nelson, J.A. Human cytomegalovirus tropism for endothelial cells: not all endothelial cells are created equal. J. Virol. 2007, 81, 2095–2101. [Google Scholar] [CrossRef]
- Crawford, L.B.; Kim, J.H.; Collins-McMillen, D.; Lee, B.J.; Landais, I.; Held, C.; Nelson, J.A.; Yurochko, A.D.; Caposio, P. Human Cytomegalovirus Encodes a Novel FLT3 Receptor Ligand Necessary for Hematopoietic Cell Differentiation and Viral Reactivation. MBio 2018. [Google Scholar] [CrossRef]
- Crawford, L.B.; Caposio, P.; Kreklywich, C.; Pham, A.H.; Hancock, M.H.; Jones, T.A.; Smith, P.P.; Yurochko, A.D.; Nelson, J.A.; Streblow, D.N. Human Cytomegalovirus US28 Ligand Binding Activity Is Required for Latency in CD34(+) Hematopoietic Progenitor Cells and Humanized NSG Mice. MBio 2019, 10, e01889-19. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crawford, L.B.; Tempel, R.; Streblow, D.N.; Yurochko, A.D.; Goodrum, F.D.; Nelson, J.A.; Caposio, P. Human Cytomegalovirus Infection Suppresses CD34+ Progenitor Cell Engraftment in Humanized Mice. Microorganisms 2020, 8, 525. https://doi.org/10.3390/microorganisms8040525
Crawford LB, Tempel R, Streblow DN, Yurochko AD, Goodrum FD, Nelson JA, Caposio P. Human Cytomegalovirus Infection Suppresses CD34+ Progenitor Cell Engraftment in Humanized Mice. Microorganisms. 2020; 8(4):525. https://doi.org/10.3390/microorganisms8040525
Chicago/Turabian StyleCrawford, Lindsey B., Rebecca Tempel, Daniel N. Streblow, Andrew D. Yurochko, Felicia D. Goodrum, Jay A. Nelson, and Patrizia Caposio. 2020. "Human Cytomegalovirus Infection Suppresses CD34+ Progenitor Cell Engraftment in Humanized Mice" Microorganisms 8, no. 4: 525. https://doi.org/10.3390/microorganisms8040525
APA StyleCrawford, L. B., Tempel, R., Streblow, D. N., Yurochko, A. D., Goodrum, F. D., Nelson, J. A., & Caposio, P. (2020). Human Cytomegalovirus Infection Suppresses CD34+ Progenitor Cell Engraftment in Humanized Mice. Microorganisms, 8(4), 525. https://doi.org/10.3390/microorganisms8040525