Effects of In Ovo Supplementation with Nanonutrition (L-Arginine Conjugated with Ag NPs) on Muscle Growth, Immune Response and Heat Shock Proteins at Different Chicken Embryonic Development Stages

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Chemicals
2.3. Synthesis of Ag NPs Using BOL Extracts
2.4. Synthesis of Ag NPs’ Composites Using Polyvinylpyrrolidone (PVP)
2.5. Characterization of Ag NPs
2.6. Experimental Design and Incubation
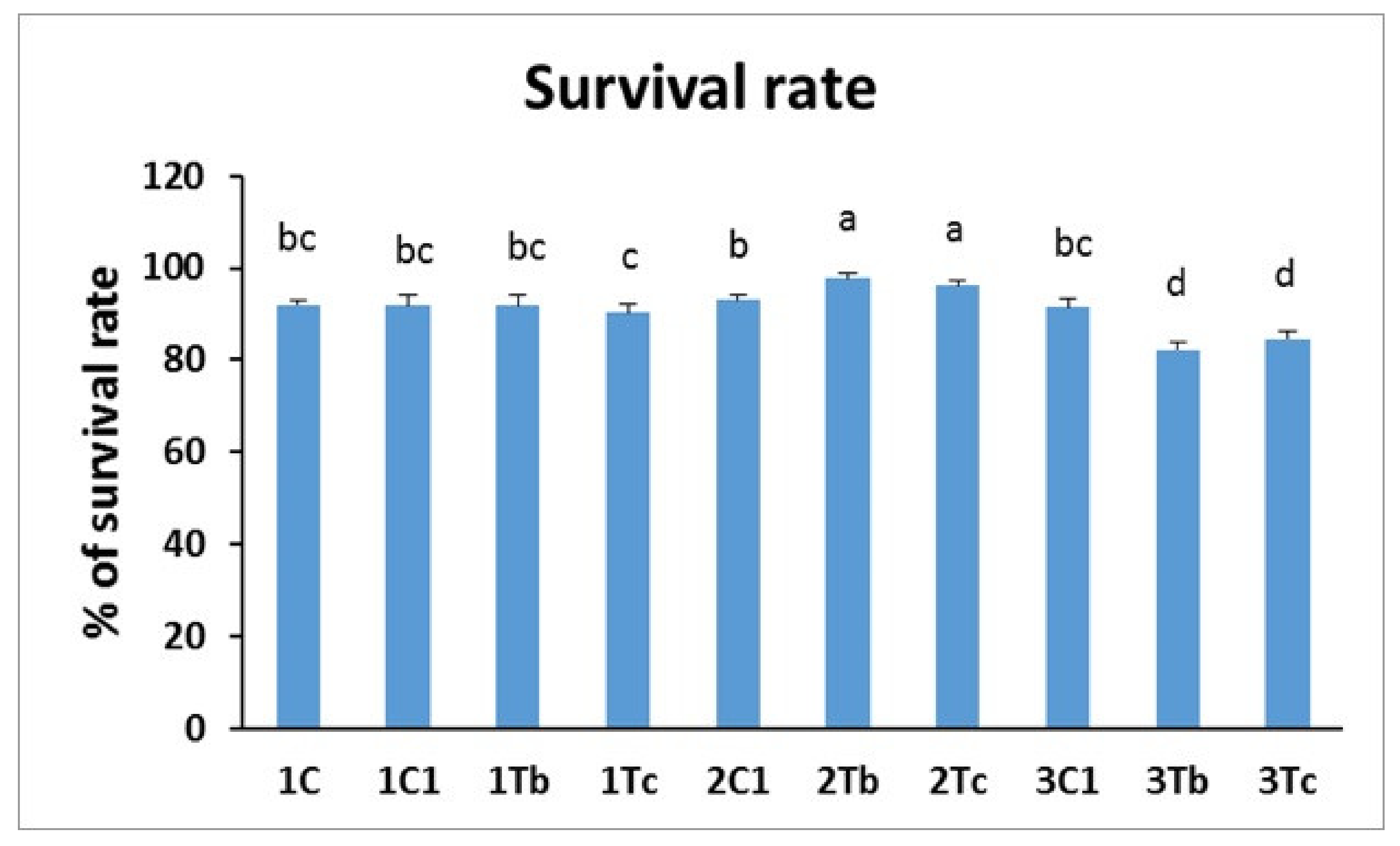
2.7. Survival Rate Measurement
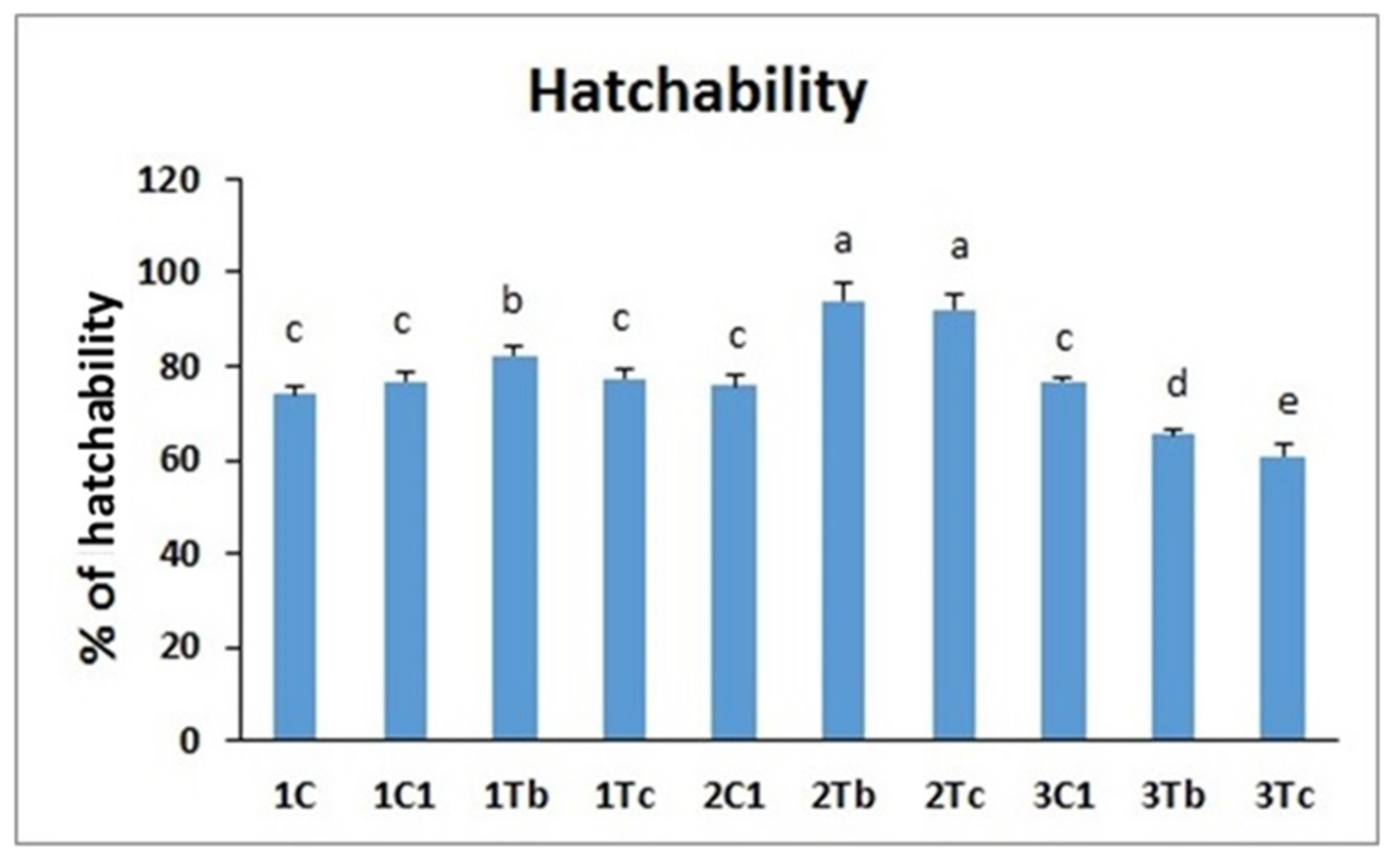
2.8. Hatching Rate and Body Weight Measurements
2.9. Biochemical Indices
2.10. Measurement of IgM Concentration in Serum
2.11. Analysis of Heat-Shock Proteins (HSPs) and Muscle Related Markers by Western Blot
3. Statistical Analysis
4. Results and Discussion
4.1. Characteristics of Ag NPs
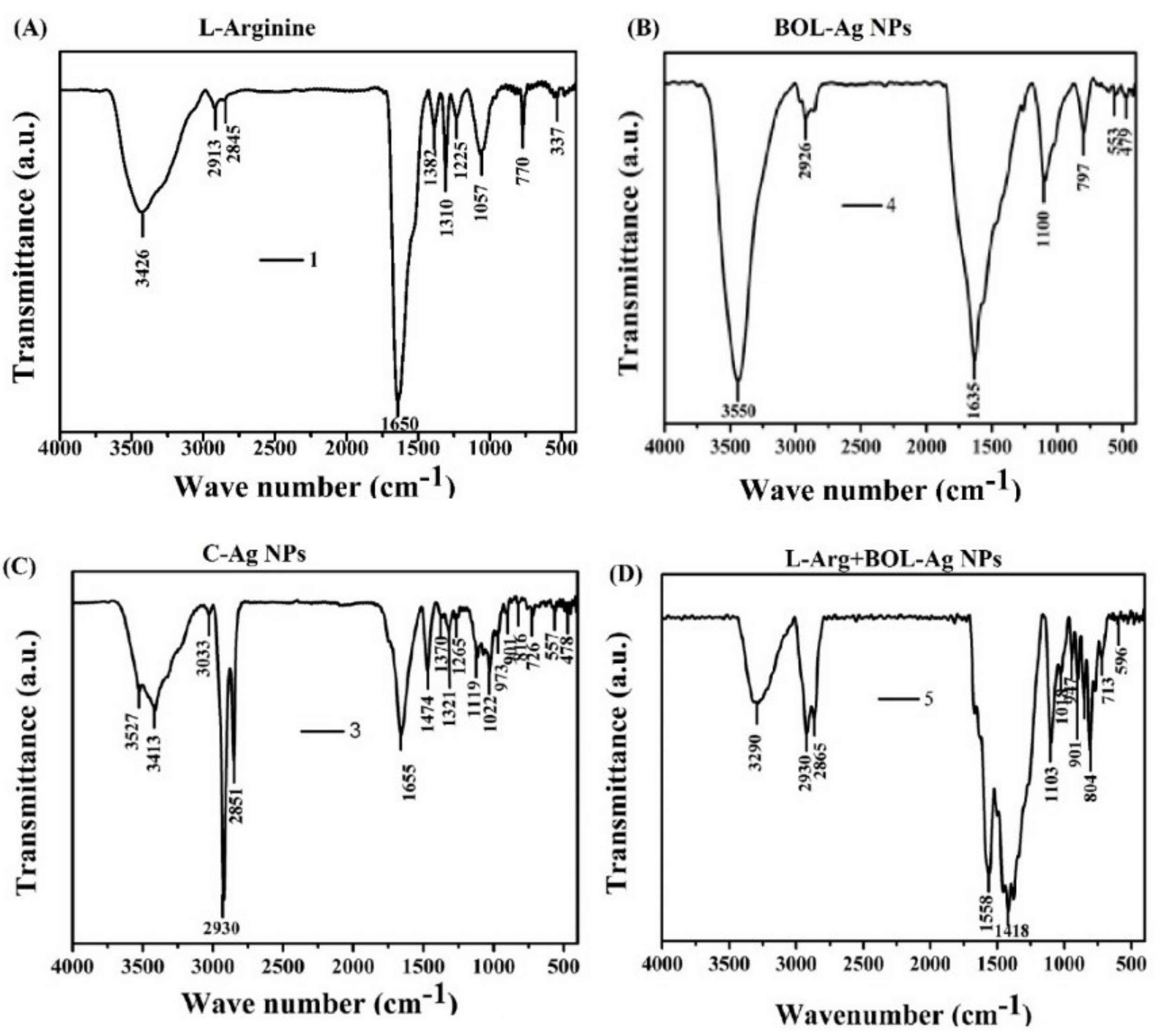
4.2. FT-IR
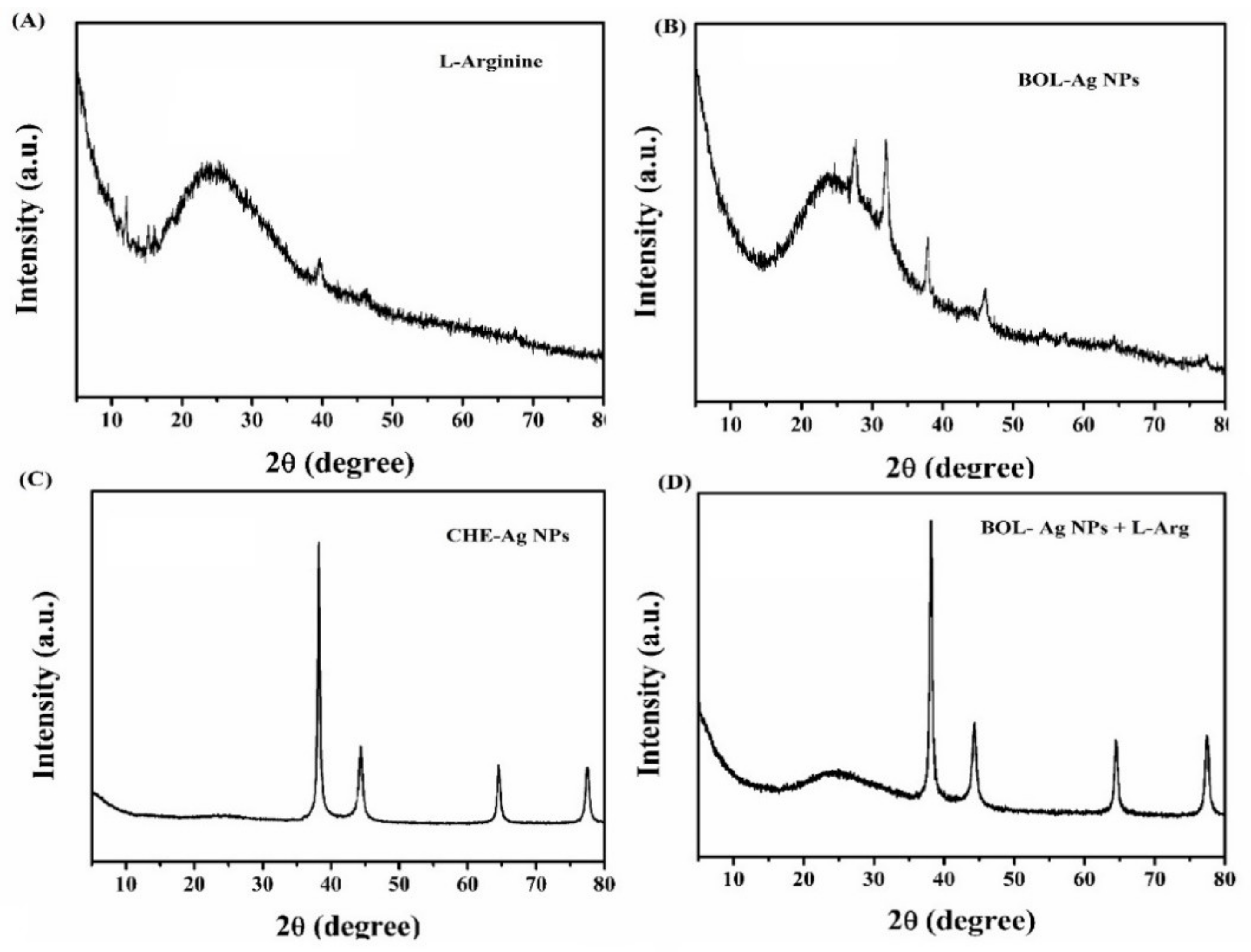
4.3. XRD
4.4. In Ovo Study
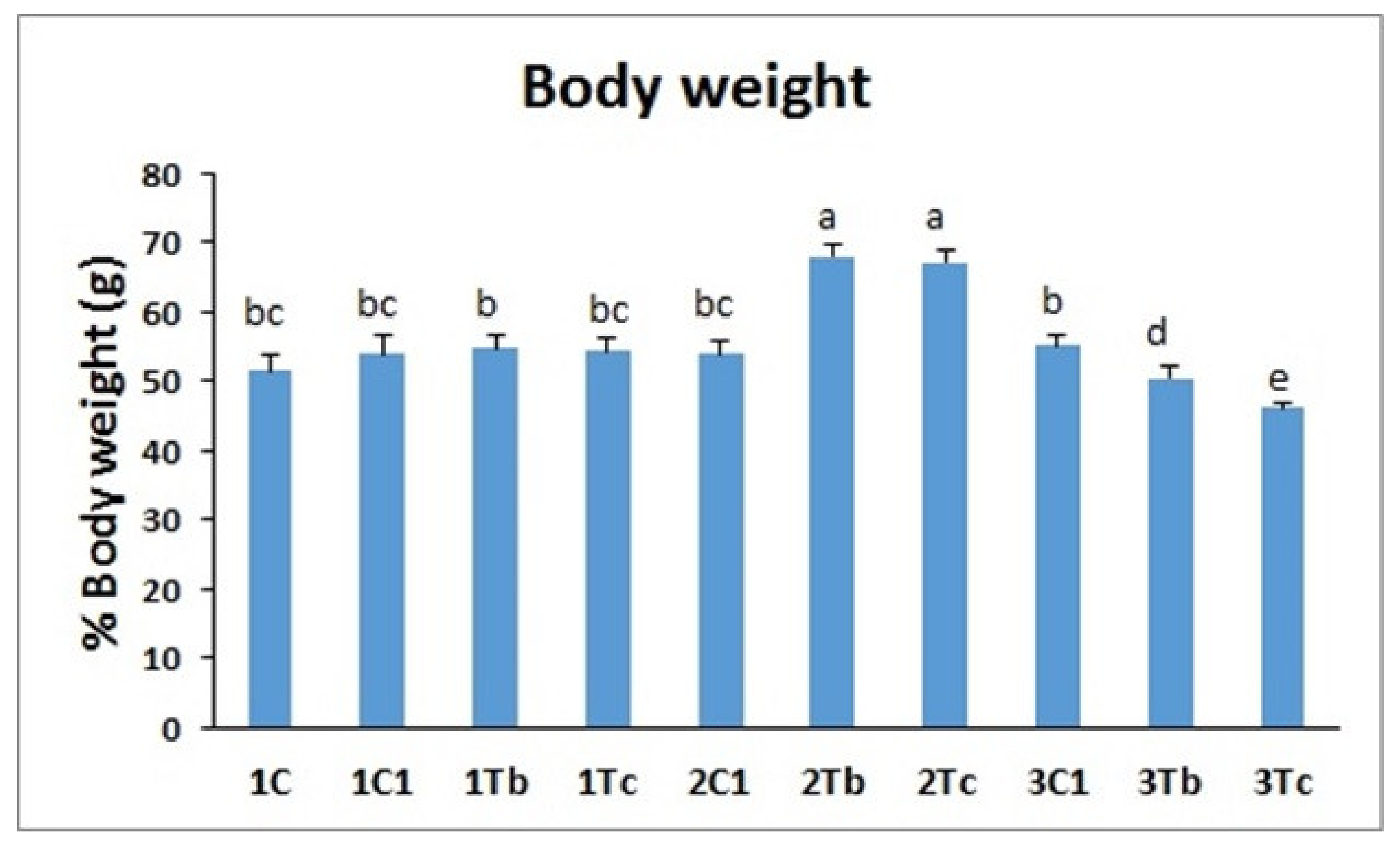
4.5. Body Weight
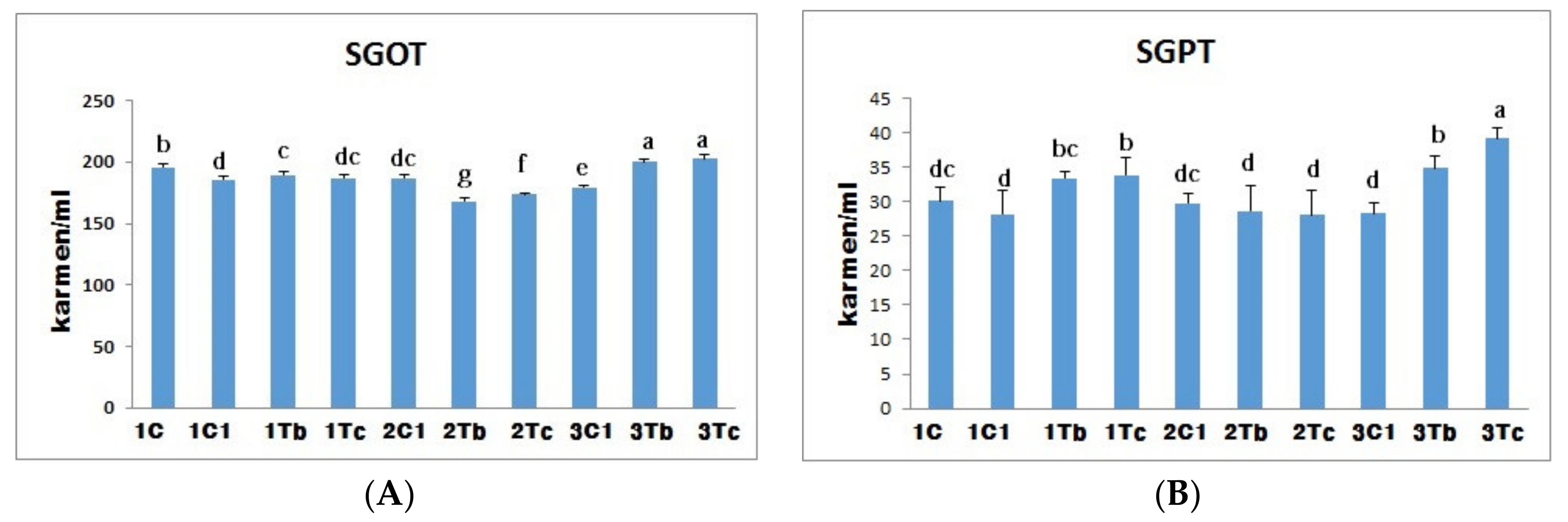
4.6. Biochemical Indices (SGOT and SGPT)
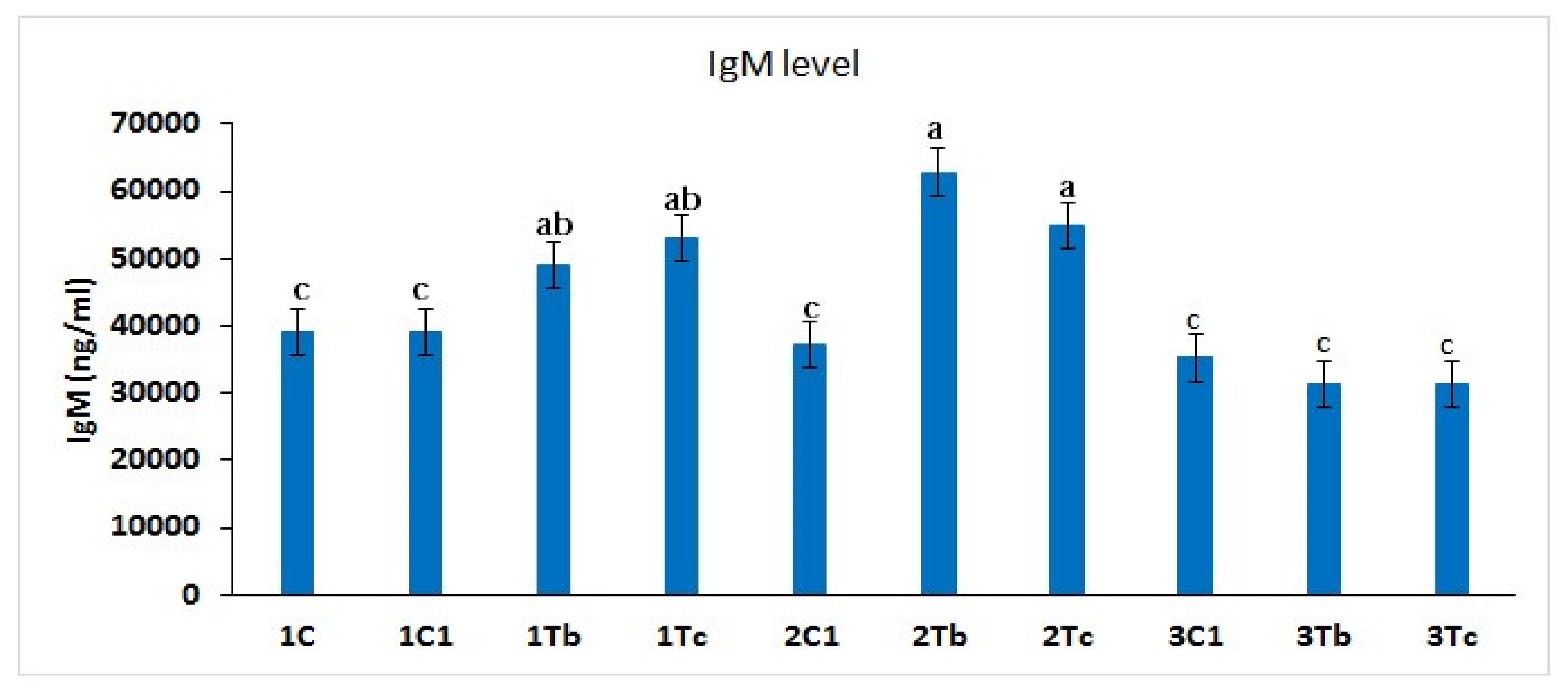
4.7. Measurement of IgM Concentration in Serum
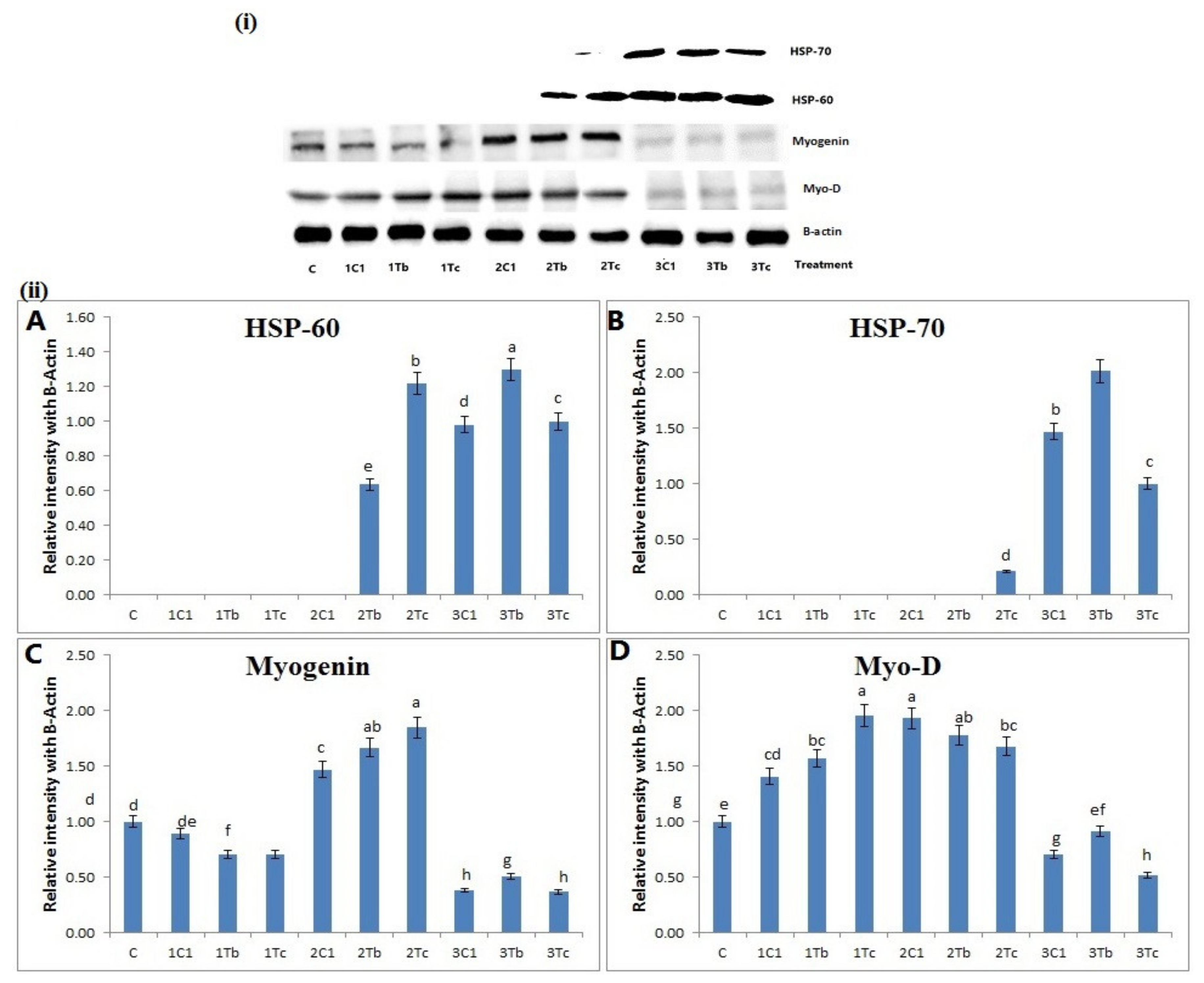
4.8. Protein Analysis by Western Blot
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beck, I.; Hotowy, A.; Sawosz, E.; Grodzik, M.; Wierzbicki, M.; Kutwin, M.; Jaworski, S.; Chwalibog, A. Effect of silver nanoparticles and hydroxyproline, administered in ovo, on the development of blood vessels and cartilage collagen structure in chicken embryos. Arch. Anim. Nutr. 2015, 69, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Lalhriatpuii, M. Progress and Prospect of Essential Mineral Nanoparticles in Poultry Nutrition and Feeding—A Review. Biol. Trace Elem. Res. 2019, 1–21. [Google Scholar] [CrossRef]
- Grodzik, M.; Sawosz, F.; Sawosz, E.; Hotowy, A.; Wierzbicki, M.; Kutwin, M.; Jaworski, S.; Chwalibog, A. Nano-nutrition of chicken embryos. The effect of in ovo administration of diamond nanoparticles and L-glutamine on molecular responses in chicken embryo pectoral muscles. Int. J. Mol. 2013, 14, 23033–23044. [Google Scholar] [CrossRef]
- Ohta, Y.; Kidd, M.T.; Ishibashi, T. Embryo growth and amino acid concentration profiles of broiler breeder eggs, embryos, and chicks after in ovo administration of amino acids. Poult. Sci. 2001, 80, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Bhanja, S.K.; Mandal, A.B. Effect of in ovo injection of critical amino acids on pre-and post-hatch growth, immunocompetence and development of digestive organs in broiler chickens. Asian Austral. J. Anim. 2005, 18, 524–531. [Google Scholar] [CrossRef]
- Murakami, A.E.; Fernandes, J.I.; Hernandes, L.; Santos, T.C. Effects of starter diet supplementation with arginine on broiler production performance and on small intestine morphometry. Pesqui. Vet. Brasil. 2012, 32, 259–266. [Google Scholar] [CrossRef]
- Azhar, M.; Rahardja, D.P.; Pakiding, W. Embryo Development and Post-Hatch Performances of Kampung Chicken by in Ovo Feeding of L-Arginine. Media Peternak. 2016, 39, 168–172. [Google Scholar] [CrossRef]
- Subramaniyan, S.A.; Kang, D.R.; Park, J.R.; Siddiqui, S.H.; Ravichandiran, P.; Yoo, D.J.; Na, C.S.; Shim, K.S. Effect of In Ovo Injection of L-Arginine in Different Chicken Embryonic Development Stages on Post-Hatchability, Immune Response, and Myo-D and Myogenin Proteins. Animals 2019, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Subramaniyan, S.A.; Belal, S.A.; Choe, H.S.; Shim, K.S. A Comparative Study of Biologically and Inorganic Fabricated Synthesized AgNPs’ Supplementation with Respect to Heat-Shock Proteins, Survival, and Hatching Rates of Chicken Embryos: An In Ovo Study. J. Clust. Sci. 2018, 29, 129–139. [Google Scholar] [CrossRef]
- Sawosz, F.; Pineda, L.; Hotowy, A.; Hyttel, P.; Sawosz, E.; Szmidt, M.; Niemiec, T.; Chwalibog, A. Nano-nutrition of chicken embryos. The effect of silver nanoparticles and glutamine on molecular responses, and the morphology of pectoral muscle. Baltic J. Comp. Clin. Syst. Biol. 2012, 2, 29–45. [Google Scholar]
- Bhanja, S.; Hotowy, A.; Mehra, M.; Sawosz, E.; Pineda, L.; Vadalasetty, K.; Kurantowicz, N.; Chwalibog, A. In ovo administration of silver nanoparticles and/or amino acids influence metabolism and immune gene expression in chicken embryos. Int. J. Mol. 2015, 16, 9484–9503. [Google Scholar] [CrossRef] [PubMed]
- Velleman, S.G. Muscle development in the embryo and hatchling. Poult. Sci. 2007, 86, 1050–1054. [Google Scholar] [CrossRef]
- Yu, L.L.; Gao, T.; Zhao, M.M.; Lv, P.A.; Zhang, L.; Li, J.L.; Jiang, Y.; Gao, F.; Zhou, G.H. In ovo feeding of L-arginine alters energy metabolism in post-hatch broilers. Poult. Sci. 2018, 97, 140–148. [Google Scholar] [CrossRef]
- Ramachandran, R.; Krishnaraj, C.; Sivakumar, A.S.; Prasannakumar, P.; Kumar, V.A.; Shim, K.S.; Song, C.G.; Yun, S.I. Anticancer activity of biologically synthesized silver and gold nanoparticles on mouse myoblast cancer cells and their toxicity against embryonic zebrafish. Mater. Sci. Eng. C 2017, 73, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Shah, S.; Devi, S. Microwave-assisted synthesis of silver nanoparticles using ethanol as a reducing agent. Mater. Chem. Phys. 2009, 114, 530–532. [Google Scholar] [CrossRef]
- Keralapurath, M.M.; Corzo, A.; Pulikanti, R.; Zhai, W.; Peebles, E.D. Effects of in ovo injection of L-carnitine on hatchability and subsequent broiler performance and slaughter yield. Poult. Sci. 2010, 89, 1497–1501. [Google Scholar] [CrossRef]
- Bhanja, S.K.; Mandal, A.B.; Majumdar, S.; Mehra, M.; Goel, A. Effect of in ovo injection of vitamins on the chick weight and post-hatch growth performance in broiler chickens. Indian J. Poult. Sci. 2012, 47, 306–310. [Google Scholar]
- Ibrahim, N.; Wakwak, M.; Khalifa, H.; Dep, B.A. Effect of in ovo injection of some nutrients and vitamins upon improving hatchability and hatching performances of ostrich embroyos. Egypt. Poult. Sci. 2012, 32, 981–994. [Google Scholar]
- Pineda, L.; Chwalibog, A.; Sawosz, E.; Lauridsen, C.; Engberg, R.; Elnif, J.; Hotowy, A.; Sawosz, F.; Gao, Y.; Ali, A.; et al. Effect of silver nanoparticles on growth performance, metabolism and microbial profile of broiler chickens. Arch. Anim. Nutr. 2012, 66, 416–429. [Google Scholar] [CrossRef]
- Vilchez, C.; Touchburn, S.P.; Chavez, E.R.; Chan, C.W. Effect of feeding palmitic, oleic, and linoleic acids to Japanese quail hens (Coturnix coturnix japonica). 2. Maternal diets and stage of incubation on the lipid metabolism of quail embryos. Poult. Sci. 1992, 71, 1032–1042. [Google Scholar] [CrossRef]
- Nouboukpo, K.E.; Tona, K.; Kamers, B.; Everaert, N.; Willemsen, H.; Onagbesan, O.M.; Gbeassor, M.; Decuypere, E. Effects of in ovo administration of L-carnitine on hatching events and juvenile performance of layer-type chick. Int. J. Poult. Sci. 2010, 9, 980–983. [Google Scholar] [CrossRef]
- Al-Daraji, H.J.; Tahir, A.O. Effect of L-carnitine on fertility, hatchability and sex hormones in duck breeder. Res. Opin. Anim. Vet. Sci. 2014, 4, 608–613. [Google Scholar]
- Hotowy, A.; Sawosz, E.; Pineda, L.; Sawosz, F.; Grodzik, M.; Chwalibog, A. Silver nanoparticles administered to chicken affect VEGFA and FGF2 gene expression in breast muscle and heart. Nanoscale Res. Lett. 2012, 7, 418. [Google Scholar] [CrossRef] [PubMed]
- Debbage, P.; Thurner, G.C. Nanomedicine faces barriers. Pharmaceuticals 2010, 3, 3371–3416. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, X.; Wei, Y.; Liu, W.; Li, S.; Yu, G.; Fu, X.; Cao, T.; Deng, X. Cytotoxicity of silver nanoparticles in human embryonic stem cell-derived fibroblasts and an L-929 cell line. J. Nanomater. 2012, 2012, 160145. [Google Scholar] [CrossRef]
- Daniel, S.K.; Tharmaraj, V.; Sironmani, T.A.; Pitchumani, K. Toxicity and immunological activity of silver nanoparticles. Appl. Clay Sci. 2010, 48, 547–551. [Google Scholar] [CrossRef]
- Sawosz, E.; Grodzik, M.; Zielińska, M.; Niemiec, T.; Olszańska, B.; Chwalibog, A. Nanoparticles of silver do not affect growth, development and DNA oxidative damage in chicken embryos. Arch. Geflugelkd. 2009, 73, 208–213. [Google Scholar]
- Cela, P.; Vesela, B.; Matalova, E.; Vecera, Z.; Buchtova, M. Embryonic toxicity of nanoparticles. Cells Tissues Organs 2014, 199, 1–23. [Google Scholar] [CrossRef]
- Wong, K.K.; Liu, X. Silver nanoparticles—The real “silver bullet” in clinical medicine. MedChemComm 2010, 1, 125–131. [Google Scholar] [CrossRef]
- Rabie, M.H.; Ismail, F.S.A.; Ahmed, A.A.S. Effect of in ovo injection of L-carnitine at different incubational ages on egg hatchability in broiler breeders and post-hatch performance. Asian J. Anim. Vet. Adv. 2015, 10, 875–884. [Google Scholar] [CrossRef]
- Ahmadi, F.; Branch, S. Impact of different levels of silver nanoparticles (Ag-NPs) on performance, oxidative enzymes and blood parameters in broiler chicks. Pak. Vet. J. 2012, 32, 325–328. [Google Scholar]
- Bhanja, S.K.; Sudhagar, M.; Goel, A.; Pandey, N.; Mehra, M.; Agarwal, S.K.; Mandal, A. Differential expression of growth and immunity related genes influenced by in ovo supplementation of amino acids in broiler chickens. Czech. J. Anim. Sci. 2014, 59, 399–408. [Google Scholar] [CrossRef]
- Vinoth, A.; Thirunalasundari, T.; Shanmugam, M.; Uthrakumar, A.; Suji, S.; Rajkumar, U. Evaluation of DNA methylation and mRNA expression of heat shock proteins in thermal manipulated chicken. Cell Stress Chaperones 2018, 23, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Hasheimi, S.R.; Zulkifli, I.; Somchit, M.N.; Zunita, Z.; Loh, T.C.; Soleimani, A.F.; Tang, S.C. Dietary supplementation of Zingiber officinale and Zingiber zerumbet to heat-stressed broiler chickens and its effect on heat shock protein 70 expression, blood parameters and body temperature. J. Anim. Physiol. Anim. Nutr. 2013, 97, 632–638. [Google Scholar] [CrossRef]
- Al-Rasheed, N.M.; Fadda, L.; Mohamed, A.M.; Attia, H.A.; Al-Rasheed, N.M. Regulating effect of carnosine and/or l-arginine on the expression of inflammatory molecules induced nephropathy in the hypoxic rat model. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef]
- Murray, J.; Auwerx, J.; Huss, J.M. Impaired myogenesis in estrogen-related receptor (ERRγ)-deficient skeletal myocytes due to oxidative stress. FASEB J. 2013, 27, 135–150. [Google Scholar] [CrossRef]
- Wust, R.C.; Degens, H. Factors contributing to muscle wasting and dysfunction in COPD patients. Int. J. Chron. Obstruct. Pulmon. Dis. 2007, 2, 289–300. [Google Scholar]
- Mok, G.F.; Mohammed, R.H.; Sweetman, D. Expression of myogenic regulatory factors in chicken embryos during somite and limb development. J. Anat. 2015, 227, 352–360. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dosage | No. of Replication | Total No. of Eggs |
|---|---|---|---|
| 1C | Control | 4 | 80 |
| 1C1 (8th day) | PBS/100 µL/egg | 4 | 80 |
| 1Tb (8th day) | 100 µg (L-Arg) + 1000 µg (BOL-Ag NPs)/100 µL/egg | 4 | 80 |
| 1Tc (8th day) | 100 µg (L-Arg) + 100 µg (C-Ag NPs)/100 µL/egg | 4 | 80 |
| 2C1 (14th day) | PBS/100 µL/egg | 4 | 80 |
| 2Tb (14th day) | 100 µg (L-Arg) + 1000 µg (BOL-Ag NPs)/100 µL/egg | 4 | 80 |
| 2Tc (14th day) | 100 µg (L-Arg) + 100 µg (C-Ag NPs)/100 µL/egg | 4 | 80 |
| 3C1 (18th day) | PBS/100 µL/egg | 4 | 80 |
| 3Tb (18th day) | 100 µg (L-Arg) + 1000 µg (BOL-Ag NPs)/100 µL/egg | 4 | 80 |
| 3Tc (18th day) | 100 µg (L-Arg) + 100 µg (C-Ag NPs)/100 µL/egg | 4 | 80 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramaniyan, S.A.; Kang, D.; Siddiqui, S.H.; Park, J.; Tian, W.; Park, B.; Shim, K. Effects of In Ovo Supplementation with Nanonutrition (L-Arginine Conjugated with Ag NPs) on Muscle Growth, Immune Response and Heat Shock Proteins at Different Chicken Embryonic Development Stages. Animals 2020, 10, 564. https://doi.org/10.3390/ani10040564
Subramaniyan SA, Kang D, Siddiqui SH, Park J, Tian W, Park B, Shim K. Effects of In Ovo Supplementation with Nanonutrition (L-Arginine Conjugated with Ag NPs) on Muscle Growth, Immune Response and Heat Shock Proteins at Different Chicken Embryonic Development Stages. Animals. 2020; 10(4):564. https://doi.org/10.3390/ani10040564
Chicago/Turabian StyleSubramaniyan, Sivakumar Allur, Darae Kang, Sharif Hasan Siddiqui, Jinryong Park, Weishun Tian, Byungyong Park, and Kwanseob Shim. 2020. "Effects of In Ovo Supplementation with Nanonutrition (L-Arginine Conjugated with Ag NPs) on Muscle Growth, Immune Response and Heat Shock Proteins at Different Chicken Embryonic Development Stages" Animals 10, no. 4: 564. https://doi.org/10.3390/ani10040564
APA StyleSubramaniyan, S. A., Kang, D., Siddiqui, S. H., Park, J., Tian, W., Park, B., & Shim, K. (2020). Effects of In Ovo Supplementation with Nanonutrition (L-Arginine Conjugated with Ag NPs) on Muscle Growth, Immune Response and Heat Shock Proteins at Different Chicken Embryonic Development Stages. Animals, 10(4), 564. https://doi.org/10.3390/ani10040564