
Tumor Immune Microenvironment and Its Clinicopathological and Prognostic Associations in Canine Splenic Hemangiosarcoma
 , ,
, ,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Selection
- histological diagnosis of HSA;
- in case of poorly differentiated tumors, a diagnosis of endothelial origin had to be confirmed by immunolabeling with vWF (FVIII) and/or CD31;
- available neoplastic tissue with an area >0.5 cm2.
2.2. Immunohistochemistry
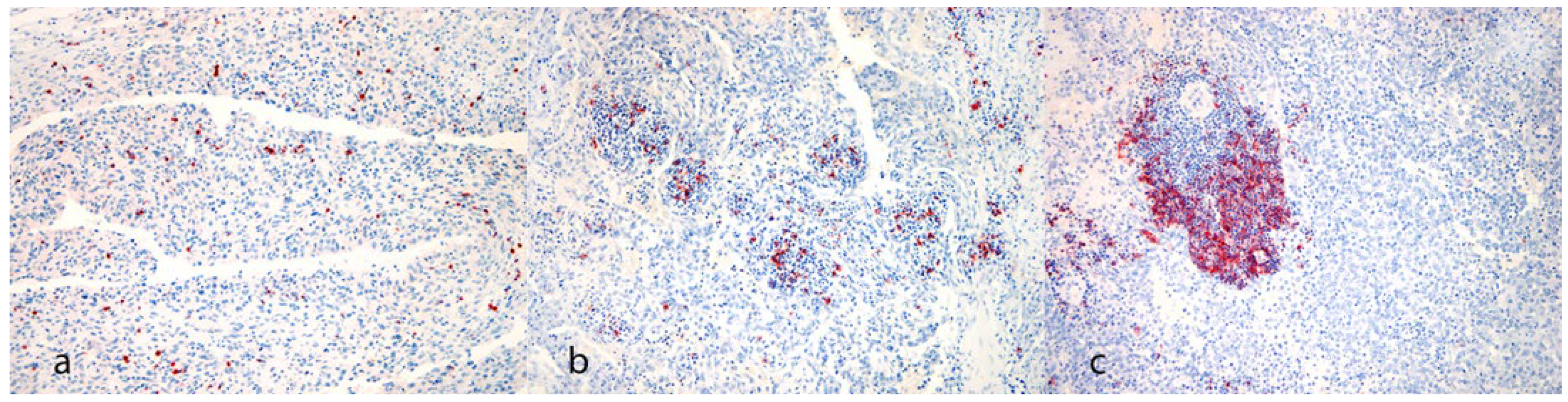
- Iba-1, CTLA-4, FoxP3, CD3, CD20: “Distribution” (Model of distribution): 0 = absent, 1 = focal, 2 = multifocal, 3 = diffuse.
- CD3, CD20: “Groups” (Arrangement of positive cells): 0 = single cells, 1 = small aggregates, 2 = massive aggregates (≥30 positive cells). If more than one type of arrangement of inflammatory cells was present in the same case, the final score was assigned to the more commonly represented type.
- Iba-1, CTLA-4, FoxP3, CD3, CD20: “Quantity”: average number of positive cells per unit area (0.237 mm2). If there were massive lymphocyte aggregations, the wording “>100” was used.
2.3. Statistical Analysis
3. Results
3.1. Patients Characteristics and Immunohistochemical Findings
3.2. Prognostic Significance of Immunohistochemical TIME-Related Markers in Canine Splenic Hemangiosarcoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.H.; Graef, A.J.; Dickerson, E.B.; Modiano, J.F. Pathobiology of hemangiosarcoma in dogs: Research advances and future perspectives. Vet. Sci. 2015, 2, 388–405. [Google Scholar] [CrossRef]
- Lamerato-Kozicki, A.R.; Helm, K.M.; Jubala, C.M.; Cutter, G.C.; Modiano, J.F. Canine hemangiosarcoma originates from hematopoietic precursors with potential for endothelial differentiation. Exp. Hematol. 2006, 34, 870–878. [Google Scholar] [CrossRef]
- Pimentel, P.A.B.; Giuliano, A.; Bęczkowski, P.M.; Horta, R.D.S. Molecular Profile of Canine Hemangiosarcoma and Potential Novel Therapeutic Targets. Vet. Sci. 2023, 10, 387. [Google Scholar] [CrossRef]
- Withrow, S.J.; Vail, D.M.; Liptak, J.M. Withrow and MacEwen’s Small Animal Clinical Oncology, 6th ed.; Elsevier Health Sciences: St. Louis, MS, USA, 2013; ISBN 0854046356. [Google Scholar]
- Eberle, N.; von Babo, V.; Nolte, I.; Baumgärtner, W.; Betz, D. Splenic masses in dogs. Part 1: Epidemiologic, clinical characteristics as well as histopathologic diagnosis in 249 cases (2000–2011). Tierarztl Prax Ausg K Kleintiere Heimtiere 2012, 40, 250–260. [Google Scholar]
- Borgatti, A.; Winter, A.L.; Stuebner, K.; Scott, R.; Ober, C.P.; Anderson, K.L.; Feeney, D.A.; Vallera, D.A.; Koopmeiners, J.S.; Modiano, J.F.; et al. Evaluation of 18-F-fluoro-2-deoxyglucose (FDG) positron emission tomography/computed tomography (PET/CT) as a staging and monitoring tool for dogs with stage-2 splenic hemangiosarcoma—A pilot study. PLoS ONE 2017, 12, e0172651. [Google Scholar] [CrossRef]
- Kim, J.H.; Megquier, K.; Thomas, R.; Sarver, A.L.; Song, J.M.; Kim, Y.T.; Cheng, N.; Schulte, A.J.; Linden, M.A.; Murugan, P.; et al. Genomically Complex Human Angiosarcoma and Canine Hemangiosarcoma Establish Convergent Angiogenic Transcriptional Programs Driven by Novel Gene Fusions. Mol. Cancer Res. 2021, 19, 847–861. [Google Scholar] [CrossRef]
- Megquier, K.; Turner-Maier, J.; Swofford, R.; Kim, J.H.; Sarver, A.L.; Wang, C.; Sakthikumar, S.; Johnson, J.; Koltookian, M.; Lewellen, M.; et al. Comparative Genomics Reveals Shared Mutational Landscape in Canine Hemangiosarcoma and Human Angiosarcoma. Mol. Cancer Res. 2019, 17, 2410–2421. [Google Scholar] [CrossRef]
- Rodrigues, L.; Watson, J.; Feng, Y.; Lewis, B.; Harvey, G.; Post, G.; Megquier, K.; White, M.E.; Lambert, L.; Miller, A.; et al. Shared hotspot mutations in oncogenes position dogs as an unparalleled comparative model for precision therapeutics. Sci. Rep. 2023, 13, 10935. [Google Scholar] [CrossRef]
- Wong, S.; Ehrhart, E.J.; Stewart, S.; Zismann, V.; Cawley, J.; Halperin, R.; Briones, N.; Richter, K.; Sivaprakasam, K.; Perdigones, N.; et al. Genomic landscapes of canine splenic angiosarcoma (hemangiosarcoma) contain extensive heterogeneity within and between patients. PLoS ONE 2022, 17, e0264986. [Google Scholar] [CrossRef]
- Kapturska, K.M.; Pawlak, A. New molecular targets in canine hemangiosarcoma—Comparative review and future of the precision medicine. Vet. Comp. Oncol. 2023, 21, 357–377. [Google Scholar] [CrossRef]
- Farver, C.; Crain, D.; Cernat, M.; Ally, A.; Karlan, B.Y.; Schilero, C.; von Deimling, A.; Tcaciuc, D.; Moran, C.; Wilmott, J.; et al. Genomic, Pathway Network, and Immunologic Features Distinguishing Squamous Carcinomas. Cell Rep. 2018, 23, 194–212.e6. [Google Scholar] [CrossRef]
- Wong, K.; Ludwig, L.; Krijgsman, O.; Adams, D.J.; Wood, G.A.; Van Der Weyden, L. Comparison of the oncogenomic landscape of canine and feline hemangiosarcoma shows novel parallels with human angiosarcoma. Dis. Model. Mech. 2021, 14, dmm049044. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, M.; Maloneyhuss, M.A.; Wojcik, J.; Durham, A.C.; Mason, N.J.; Roth, D.B. Actionable mutations in canine hemangiosarcoma. PLoS ONE 2017, 12, e0188667. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.S.; Rassnick, K.M.; Frimberger, A.E. Evaluation of clinical and histologic factors associated with survival time in dogs with stage II splenic hemangiosarcoma treated by splenectomy and adjuvant chemotherapy: 30 cases (2011–2014). J. Am. Vet. Med. Assoc. 2017, 251, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, G.K.; Powers, B.E.; Mallinckrodt, C.H.; Withrow, S.J. Surgery and doxorubicin in dogs with hemangiosarcoma. J. Vet. Intern. Med. 1996, 10, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Avallone, G.; Rasotto, R.; Chambers, J.K.; Miller, A.D.; Behling-Kelly, E.; Monti, P.; Berlato, D.; Valenti, P.; Roccabianca, P. Review of Histological Grading Systems in Veterinary Medicine. Vet. Pathol. 2021, 58, 809–828. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.-T.; Nekouei, O.; Sandy, J.R. Histopathological Grading, Clinical Staging and CD 31 Expression of Canine Splenic Hemangiosarcoma. Vet. Sci. 2023, 10, 190. [Google Scholar] [CrossRef]
- Batschinski, K.; Nobre, A.; Vargas-Mendez, E.; Tedardi, M.V.; Cirillo, J.; Cestari, G.; Ubukata, R.; Dagli, M.L.Z. Canine visceral hemangiosarcoma treated with surgery alone or surgery and doxorubicin: 37 cases (2005–2014). Can. Vet. J. 2018, 59, 967. [Google Scholar]
- Wendelburg, K.M.; Price, L.L.; Burgess, K.E.; Lyons, J.A.; Lew, F.H.; Berg, J. Survival time of dogs with splenic hemangiosarcoma treated by splenectomy with or without adjuvant chemotherapy: 208 cases (2001–2012). J. Am. Vet. Med. Assoc. 2015, 247, 393–403. [Google Scholar] [CrossRef]
- Johnson, K.A.; Powers, B.E.; Withrow, S.J.; Sheetz, M.J.; Curtis, C.R.; Wrigley, R.H. Splenomegaly in dogs. Predictors of neoplasia and survival after splenectomy. J. Vet. Intern. Med. 1989, 3, 160–166. [Google Scholar] [CrossRef]
- Wood, C.A.; Moore, A.S.; Gliatto, J.M.; Ablin, L.A.; Berg, R.J.; Rand, W.M. Prognosis for dogs with stage I or II splenic hemangiosarcoma treated by splenectomy alone: 32 cases (1991–1993). J. Am. Anim. Hosp. Assoc. 1998, 34, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Patnaik, A.; MacEwen, E. Canine hemangiosarcoma: Retrospective analysis of 104 cases. J. Am. Vet. Med. Assoc. 1985, 186, 56–58. [Google Scholar]
- Loh, J.W.; Lee, J.Y.; Lim, A.H.; Guan, P.; Lim, B.Y.; Kannan, B.; Lee, E.C.Y.; Gu, N.X.; Ko, T.K.; Ng, C.C.Y.; et al. Spatial transcriptomics reveal topological immune landscapes of Asian head and neck angiosarcoma. Commun. Biol. 2023, 6, 461. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, V.B.; Klahn, S.; LeRoith, T.; Huckle, W.R. Canine melanoma: A review of diagnostics and comparative mechanisms of disease and immunotolerance in the era of the immunotherapies. Front. Vet. Sci. 2023, 9, 1046636. [Google Scholar] [CrossRef]
- Bongiovanni, L.; Sparger, E.; Dow, S.; Tarone, L.; Giacobino, D.; Camerino, M.; Ferrone, S.; Buracco, P.; Cavallo, F.; Riccardo, F. Canine Melanoma Immunology and Immunotherapy: Relevance of Translational Research. Front. Vet. Sci. 2022, 9, 803093. [Google Scholar] [CrossRef]
- Nicholls, P.K.; Moore, P.F.; Anderson, D.M.; Moore, R.A.; Parry, N.R.; Gough, G.W.; Stanley, M.A. Regression of Canine Oral Papillomas Is Associated with Infiltration of CD4+ and CD8+ Lymphocytes. Virology 2001, 283, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kaim, U.; Moritz, A.; Failing, K.; Baumgärtner, W. The regression of a canine Langerhans cell tumour is associated with increased expression of IL-2, TNF-α, IFN-γ and iNOS mRNA. Immunology 2006, 118, 472. [Google Scholar] [CrossRef]
- Yoon, H.H.; Shi, Q.; Heying, E.N.; Muranyi, A.; Bredno, J.; Ough, F.; Djalilvand, A.; Clements, J.; Bowermaster, R.; Liu, W.W.; et al. Intertumoral Heterogeneity of CD3+ and CD8+ T-Cell Densities in the Microenvironment of DNA Mismatch-Repair-Deficient Colon Cancers: Implications for Prognosis. Clin. Cancer Res. 2019, 25, 125–133. [Google Scholar] [CrossRef]
- Wouters, M.C.A.; Nelson, B.H. Prognostic Significance of Tumor-Infiltrating B Cells and Plasma Cells in Human Cancer. Clin. Cancer Res. 2018, 24, 6125–6135. [Google Scholar] [CrossRef]
- Saltz, J.; Gupta, R.; Hou, L.; Kurc, T.; Singh, P.; Nguyen, V.; Samaras, D.; Shroyer, K.R.; Zhao, T.; Batiste, R.; et al. Spatial Organization and Molecular Correlation of Tumor-Infiltrating Lymphocytes Using Deep Learning on Pathology Images. Cell Rep. 2018, 23, 181–193.e7. [Google Scholar] [CrossRef]
- Lo, C.S.; Sanii, S.; Kroeger, D.R.; Milne, K.; Talhouk, A.; Chiu, D.S.; Rahimi, K.; Shaw, P.A.; Clarke, B.A.; Nelson, B.H. Neoadjuvant Chemotherapy of Ovarian Cancer Results in Three Patterns of Tumor-Infiltrating Lymphocyte Response with Distinct Implications for Immunotherapy. Clin. Cancer Res. 2017, 23, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.M.; Simon, C.M. Tumor Microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y. The Function of NK Cells in Tumor Metastasis and NK Cell-Based Immunotherapy. Cancers 2023, 15, 2323. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ye, Z.; Song, F.; He, Y.; Liu, J. The Role of TAMs in Tumor Microenvironment and New Research Progress. Stem Cells Int. 2022, 2022, 5775696. [Google Scholar] [CrossRef]
- Saleh, R.; Elkord, E. FoxP3+ T regulatory cells in cancer: Prognostic biomarkers and therapeutic targets. Cancer Lett. 2020, 490, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Lindau, D.; Gielen, P.; Kroesen, M.; Wesseling, P.; Adema, G.J. The immunosuppressive tumour network: Myeloid-derived suppressor cells, regulatory T cells and natural killer T cells. Immunology 2013, 138, 105. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Koim, S.; Wrobel, M.; Käfferlein, H.U.; Brüning, T.; Stockfleth, E.; Becker, J.C.; Lang, K. Expression of Programmed Cell Death Proteins in Kaposi Sarcoma and Cutaneous Angiosarcoma. J. Immunother. 2020, 43, 169–174. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.; Guth, A.; Biller, B.; Elmslie, R.; Dow, S. Changes in Regulatory T Cells in Dogs with Cancer and Associations with Tumor Type. J. Vet. Intern. Med. 2009, 23, 875–881. [Google Scholar] [CrossRef]
- Laumont, C.M.; Banville, A.C.; Gilardi, M.; Hollern, D.P.; Nelson, B.H. Tumour-infiltrating B cells: Immunological mechanisms, clinical impact and therapeutic opportunities. Nat. Rev. Cancer 2022, 22, 414–430. [Google Scholar] [CrossRef]
- Zhang, E.; Ding, C.; Li, S.; Zhou, X.; Aikemu, B.; Fan, X.; Sun, J.; Zheng, M.; Yang, X. Roles and mechanisms of tumour-infiltrating B cells in human cancer: A new force in immunotherapy. Biomark. Res. 2023, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Vogelzang, A.; Miyajima, M.; Sugiura, Y.; Wu, Y.; Chamoto, K.; Nakano, R.; Hatae, R.; Menzies, R.J.; Sonomura, K.; et al. B cell-derived GABA elicits IL-10+ macrophages to limit anti-tumour immunity. Nature 2021, 599, 471–476. [Google Scholar] [CrossRef]
- Kuol, N.; Stojanovska, L.; Nurgali, K.; Apostolopoulos, V. The mechanisms tumor cells utilize to evade the host’s immune system. Maturitas 2017, 105, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Cornel, A.M.; Mimpen, I.L.; Nierkens, S. MHC Class I Downregulation in Cancer: Underlying Mechanisms and Potential Targets for Cancer Immunotherapy. Cancers 2020, 12, 1760. [Google Scholar] [CrossRef] [PubMed]
- Friedman, C.F.; Postow, M.A. Emerging Tissue and Blood-Based Biomarkers that may Predict Response to Immune Checkpoint Inhibition. Curr. Oncol. Rep. 2016, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Taube, J.M.; Anders, R.A.; Pardoll, D.M. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat. Rev. Cancer 2016, 16, 275–287. [Google Scholar] [CrossRef]
- Le, D.T.; Uram, J.N.; Wang, H.; Bartlett, B.R.; Kemberling, H.; Eyring, A.D.; Skora, A.D.; Luber, B.S.; Azad, N.S.; Laheru, D.; et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 372, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- André, T.; Shiu, K.-K.; Kim, T.W.; Jensen, B.V.; Jensen, L.H.; Punt, C.; Smith, D.; Garcia-Carbonero, R.; Benavides, M.; Gibbs, P.; et al. Pembrolizumab in Microsatellite-Instability–High Advanced Colorectal Cancer. N. Engl. J. Med. 2020, 383, 2207–2218. [Google Scholar] [CrossRef]
- Tumeh, P.C.; Harview, C.L.; Yearley, J.H.; Shintaku, I.P.; Taylor, E.J.M.; Robert, L.; Chmielowski, B.; Spasic, M.; Henry, G.; Ciobanu, V.; et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature 2014, 515, 568–571. [Google Scholar] [CrossRef]
- Cogdill, A.P.; Andrews, M.C.; Wargo, J.A. Hallmarks of response to immune checkpoint blockade. Br. J. Cancer 2017, 117, 1–7. [Google Scholar] [CrossRef]
- Vilain, R.E.; Menzies, A.M.; Wilmott, J.S.; Kakavand, H.; Madore, J.; Guminski, A.; Liniker, E.; Kong, B.Y.; Cooper, A.J.; Howle, J.R.; et al. Dynamic changes in PD-L1 expression and immune infiltrates early during treatment predict response to PD-1 blockade in Melanoma. Clin. Cancer Res. 2017, 23, 5024–5033. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.L.; Roh, W.; Reuben, A.; Cooper, Z.A.; Spencer, C.N.; Prieto, P.A.; Miller, J.P.; Bassett, R.L.; Gopalakrishnan, V.; Wani, K.; et al. Analysis of Immune Signatures in Longitudinal Tumor Samples Yields Insight into Biomarkers of Response and Mechanisms of Resistance to Immune Checkpoint Blockade. Cancer Discov. 2016, 6, 827–837. [Google Scholar] [CrossRef]
- Rosenberg, J.E.; Hoffman-Censits, J.; Powles, T.; Van Der Heijden, M.S.; Balar, A.V.; Necchi, A.; Dawson, N.; O’Donnell, P.H.; Balmanoukian, A.; Loriot, Y.; et al. Atezolizumab in patients with locally advanced and metastatic urothelial carcinoma who have progressed following treatment with platinum-based chemotherapy: A single-arm, multicentre, phase 2 trial. Lancet 2016, 387, 1909–1920. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.F.; Sosman, J.A.; Sznol, M.; Massard, C.; Gordon, M.S.; Hamid, O.; Powderly, J.D.; Infante, J.R.; Fassò, M.; Wang, Y.V.; et al. Atezolizumab, an Anti-Programmed Death-Ligand 1 Antibody, in Metastatic Renal Cell Carcinoma: Long-Term Safety, Clinical Activity, and Immune Correlates From a Phase Ia Study. J. Clin. Oncol. 2016, 34, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Arakawa, A.; Utsumi, D.; Sumiyoshi, S.; Yamamoto, Y.; Kitoh, A.; Ono, M.; Matsumura, Y.; Kato, M.; Konishi, K.; et al. CD8+ tumor-infiltrating lymphocytes at primary sites as a possible prognostic factor of cutaneous angiosarcoma. Int. J. Cancer 2014, 134, 2393–2402. [Google Scholar] [CrossRef]
- Lobrano, R.; Paliogiannis, P.; Zinellu, A.; Palmieri, G.; Persico, I.; Mangoni, A.A.; Cossu, A. PD-L1 Expression in Cutaneous Angiosarcomas: A Systematic Review with Meta-Analysis. Curr. Oncol. 2023, 30, 5135–5144. [Google Scholar] [CrossRef]
- Bi, Y.; Ge, L.; Ren, X.; Pang, J.; Zhao, Y.; Liang, Z. Tumor microenvironment and its clinicopathological and prognostic associations in surgically resected cutaneous angiosarcoma. Clin. Transl. Oncol. 2022, 24, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Boichard, A.; Wagner, M.J.; Kurzrock, R. Angiosarcoma heterogeneity and potential therapeutic vulnerability to immune checkpoint blockade: Insights from genomic sequencing. Genome Med. 2020, 12, 61. [Google Scholar] [CrossRef]
- Florou, V.; Rosenberg, A.E.; Wieder, E.; Komanduri, K.V.; Kolonias, D.; Uduman, M.; Castle, J.C.; Buell, J.S.; Trent, J.C.; Wilky, B.A. Angiosarcoma patients treated with immune checkpoint inhibitors: A case series of seven patients from a single institution. J. Immunother. Cancer 2019, 7, 213. [Google Scholar] [CrossRef]
- Painter, C.A.; Jain, E.; Tomson, B.N.; Dunphy, M.; Stoddard, R.E.; Thomas, B.S.; Damon, A.L.; Shah, S.; Kim, D.; Gómez Tejeda Zañudo, J.; et al. The Angiosarcoma Project: Enabling genomic and clinical discoveries in a rare cancer through patient-partnered research. Nat. Med. 2020, 26, 181–187. [Google Scholar] [CrossRef]
- Wagner, M.J.; Othus, M.; Patel, S.P.; Ryan, C.; Sangal, A.; Powers, B.; Budd, G.T.; Victor, A.I.; Hsueh, C.T.; Chugh, R.; et al. Multicenter phase II trial (SWOG S1609, cohort 51) of ipilimumab and nivolumab in metastatic or unresectable angiosarcoma: A substudy of dual anti-CTLA-4 and anti-PD-1 blockade in rare tumors (DART). J. Immunother. Cancer 2021, 9, e002990. [Google Scholar] [CrossRef]
- Tomassen, T.; Weidema, M.E.; Hillebrandt-Roeffen, M.H.S.; van der Horst, C.; Desar, I.M.E.; Flucke, U.E.; Versleijen-Jonkers, Y.M.H. Analysis of PD-1, PD-L1, and T-cell infiltration in angiosarcoma pathogenetic subgroups. Immunol. Res. 2022, 70, 256–268. [Google Scholar] [CrossRef]
- Maekawa, N.; Konnai, S.; Okagawa, T.; Nishimori, A.; Ikebuchi, R.; Izumi, Y.; Takagi, S.; Kagawa, Y.; Nakajima, C.; Suzuki, Y.; et al. Immunohistochemical analysis of PD-L1 expression in canine malignant cancers and PD-1 expression on lymphocytes in canine oral melanoma. PLoS ONE 2016, 11, e0157176. [Google Scholar] [CrossRef] [PubMed]
- Gulay, K.C.M.; Aoshima, K.; Maekawa, N.; Suzuki, T.; Konnai, S.; Kobayashi, A.; Kimura, T. Hemangiosarcoma cells induce M2 polarization and PD-L1 expression in macrophages. Sci. Rep. 2022, 12, 2124. [Google Scholar] [CrossRef] [PubMed]
- Brachelente, C.; Affolter, V.K.; Fondati, A.; Porcellato, I.; Sforna, M.; Lepri, E.; Mechelli, L.; Bongiovanni, L. CD3 and CD20 Coexpression in a Case of Canine Cutaneous Epitheliotropic T-Cell Lymphoma (Mycosis fungoides). Vet. Pathol. 2015, 53, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Porcellato, I.; Brachelente, C.; De Paolis, L.; Menchetti, L.; Silvestri, S.; Sforna, M.; Vichi, G.; Iussich, S.; Mechelli, L. FoxP3 and IDO in Canine Melanocytic Tumors. Vet. Pathol. 2019, 56, 189–199. [Google Scholar] [CrossRef]
- Porcellato, I.; Brachelente, C.; Cappelli, K.; Menchetti, L.; Silvestri, S.; Sforna, M.; Mecocci, S.; Iussich, S.; Leonardi, L.; Mechelli, L. FoxP3, CTLA-4, and IDO in Canine Melanocytic Tumors. Vet. Pathol. 2021, 58, 42–52. [Google Scholar] [CrossRef]
- Porcellato, I.; Sforna, M.; Lo Giudice, A.; Bossi, I.; Musi, A.; Tognoloni, A.; Chiaradia, E.; Mechelli, L.; Brachelente, C. Tumor-Associated Macrophages in Canine Oral and Cutaneous Melanomas and Melanocytomas: Phenotypic and Prognostic Assessment. Front. Vet. Sci. 2022, 9, 878949. [Google Scholar] [CrossRef]
- Porcellato, I.; Silvestri, S.; Menchetti, L.; Recupero, F.; Mechelli, L.; Sforna, M.; Iussich, S.; Bongiovanni, L.; Lepri, E.; Brachelente, C. Tumour-infiltrating lymphocytes in canine melanocytic tumours: An investigation on the prognostic role of CD3+ and CD20+ lymphocytic populations. Vet. Comp. Oncol. 2020, 18, 370–380. [Google Scholar] [CrossRef]
- Mizuno, T.; Suzuki, R.; Umeki, S.; Okuda, M. Crossreactivity of antibodies to canine CD25 and Foxp3 and identification of canine CD4+CD25 +Foxp3+ cells in canine peripheral blood. J. Vet. Med. Sci. 2009, 71, 1561–1568. [Google Scholar] [CrossRef]
- Meuten, D.J.; Moore, F.M.; George, J.W. Mitotic Count and the Field of View Area. Vet. Pathol. 2016, 53, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Wollina, U. Angiosarcoma: An immunogenic tumour. Br. J. Dermatol. 2018, 179, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Linden, D.; Liptak, J.M.; Vinayak, A.; Grimes, J.A.; Sandey, M.; Smiley, W.; Matz, B.M. Outcomes and prognostic variables associated with primary abdominal visceral soft tissue sarcomas in dogs: A Veterinary Society of Surgical Oncology retrospective study. Vet. Comp. Oncol. 2019, 17, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.L.; Bryan, M.E.; Atkinson, E.S.; Keeler, M.R.; Hahn, A.W.; Bryan, J.N. Neutering is associated with developing hemangiosarcoma in dogs in the Veterinary Medical Database: An age and time-period matched case-control study (1964–2003). Can. Vet. J. 2020, 61, 499. [Google Scholar]
- Cleveland, M.J.; Casale, S. Incidence of malignancy and outcomes for dogs undergoing splenectomy for incidentally detected nonruptured splenic nodules or masses: 105 cases (2009–2013). J. Am. Vet. Med. Assoc. 2016, 248, 1267–1273. [Google Scholar] [CrossRef]
- Pinello, K.; Pires, I.; Castro, A.F.; Carvalho, P.T.; Santos, A.; de Matos, A.; Queiroga, F.; Canadas-Sousa, A.; Dias-Pereira, P.; Catarino, J.; et al. Cross Species Analysis and Comparison of Tumors in Dogs and Cats, by Age, Sex, Topography and Main Morphologies. Data from Vet-OncoNet. Vet. Sci. 2022, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Carnio, A.; Eleni, C.; Cocumelli, C.; Bartolomé Del Pino, L.E.; Simeoni, S.; Spallucci, V.; Scaramozzino, P. Evaluation of intrinsic and extrinsic risk factors for dog visceral hemangiosarcoma: A retrospective case-control study register-based in Lazio region, Italy. Prev. Vet. Med. 2020, 181, 105074. [Google Scholar] [CrossRef]
- Sherwood, J.M.; Haynes, A.M.; Klocke, E.; Higginbotham, M.L.; Thomson, E.M.; Weng, H.Y.; Millard, H.A.T. Occurrence and Clinicopathologic Features of Splenic Neoplasia Based on Body Weight: 325 Dogs (2003–2013). J. Am. Anim. Hosp. Assoc. 2016, 52, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.P.; José-López, R.; Flores, F.F.; Prados, R.M.R.; Mandara, M.T.; Arús, C.; Batlle, M.P. Expression of FOXP3 in Canine Gliomas: Immunohistochemical Study of Tumor-Infiltrating Regulatory Lymphocytes. J. Neuropathol. Exp. Neurol. 2020, 79, 184–193. [Google Scholar] [CrossRef]
- Marcinowska, A.; Constantino-Casas, F.; Williams, T.; Hoather, T.; Blacklaws, B.; Dobson, J. T Lymphocytes in Histiocytic Sarcomas of Flat-Coated Retriever Dogs. Vet. Pathol. 2017, 54, 605–610. [Google Scholar] [CrossRef]
- Kim, J.H.; Hur, J.H.; Lee, S.M.; Im, K.S.; Kim, N.H.; Sur, J.H. Correlation of Foxp3 positive regulatory T cells with prognostic factors in canine mammary carcinomas. Vet. J. 2012, 193, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Biller, B.J.; Guth, A.; Burton, J.H.; Dow, S.W. Decreased ratio of CD8+ T cells to regulatory T cells associated with decreased survival in dogs with osteosarcoma. J. Vet. Intern. Med. 2010, 24, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- van Ravensteijn, S.G.; Versleijen-Jonkers, Y.M.H.; Hillebrandt-Roeffen, M.H.S.; Weidema, M.E.; Nederkoorn, M.J.L.; Bol, K.F.; Gorris, M.A.J.; Verrijp, K.; Kroeze, L.I.; de Bitter, T.J.J.; et al. Immunological and Genomic Analysis Reveals Clinically Relevant Distinctions between Angiosarcoma Subgroups. Cancers 2022, 14, 5938. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.Y.; Lim, J.Q.; Yeong, J.; Ravi, V.; Guan, P.; Boot, A.; Tay, T.K.Y.; Selvarajan, S.; Md Nasir, N.D.; Loh, J.H.; et al. Multiomic analysis and immunoprofiling reveal distinct subtypes of human angiosarcoma. J. Clin. Investig. 2020, 130, 5833–5846. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Subramaniam, A.; Zheng, J.; Amini, B.; Trinh, V.A.; Joseph, J.; Mennel, R.G.; Bishop, A.J.; Sturgis, E.M.; Goepfert, R.P.; et al. Clinical activity of checkpoint inhibitors in angiosarcoma: A retrospective cohort study. Cancer 2022, 128, 3383–3391. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, E.; Antonescu, C.R.; Smith, S.; Bradic, M.; Kashani, D.; Richards, A.L.; Donoghue, M.; Kelly, C.M.; Nacev, B.; Chan, J.E.; et al. Clinical, genomic, and transcriptomic correlates of response to immune checkpoint blockade-based therapy in a cohort of patients with angiosarcoma treated at a single center. J. Immunother. Cancer 2022, 10, e004149. [Google Scholar] [CrossRef] [PubMed]
- Yarchoan, M.; Hopkins, A.; Jaffee, E.M. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. N. Engl. J. Med. 2017, 377, 2500–2501. [Google Scholar] [CrossRef] [PubMed]
- Cristescu, R.; Mogg, R.; Ayers, M.; Albright, A.; Murphy, E.; Yearley, J.; Sher, X.; Liu, X.Q.; Lu, H.; Nebozhyn, M.; et al. Pan-tumor genomic biomarkers for PD-1 checkpoint blockade-based immunotherapy. Science 2018, 362, eaar3593. [Google Scholar] [CrossRef] [PubMed]
- Damotte, D.; Warren, S.; Arrondeau, J.; Boudou-Rouquette, P.; Mansuet-Lupo, A.; Biton, J.; Ouakrim, H.; Alifano, M.; Gervais, C.; Bellesoeur, A.; et al. The tumor inflammation signature (TIS) is associated with anti-PD-1 treatment benefit in the CERTIM pan-cancer cohort. J. Transl. Med. 2019, 17, 357. [Google Scholar] [CrossRef]
- Kerrison, W.G.; Lee, A.T.; Thway, K.; Jones, R.L.; Huang, P.H. Current Status and Future Directions of Immunotherapies in Soft Tissue Sarcomas. Biomedicines 2022, 10, 573. [Google Scholar] [CrossRef]
- Wang, J.Z.; Zhang, Y.H.; Guo, X.H.; Zhang, H.Y.; Zhang, Y. The double-edge role of B cells in mediating antitumor T-cell immunity: Pharmacological strategies for cancer immunotherapy. Int. Immunopharmacol. 2016, 36, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Kataki, A.; Scheid, P.; Piet, M.; Marie, B.; Martinet, N.; Martinet, Y.; Vignaud, J.M. Tumor infiltrating lymphocytes and macrophages have a potential dual role in lung cancer by supporting both host-defense and tumor progression. J. Lab. Clin. Med. 2002, 140, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Liu, W.; Ly, D.; Xu, H.; Qu, L.; Zhang, L. Tumor-infiltrating B cells: Their role and application in anti-tumor immunity in lung cancer. Cell. Mol. Immunol. 2019, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; de Reyniès, A.; Keung, E.Z.; Chen, T.W.W.; Sun, C.M.; Calderaro, J.; Jeng, Y.M.; Hsiao, L.P.; Lacroix, L.; Bougoüin, A.; et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, N.; Mohsenifar, Z.; Baghban, A.A.; Arjomandkhah, A. CD20+ tumor infiltrating b lymphocyte in oral squamous cell carcinoma: Correlation with clinicopathologic characteristics and heat shock protein 70 expression. Patholog. Res. Int. 2018, 2018, 4810751. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Raposo, T.; Gregório, H.; Pires, I.; Prada, J.; Queiroga, F.L. Prognostic value of tumour-associated macrophages in canine mammary tumours. Vet. Comp. Oncol. 2014, 12, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.; Gameiro, A.; Correia, J.; Ferreira, J.; Ferreira, F. The Landscape of Tumor-Infiltrating Immune Cells in Feline Mammary Carcinoma: Pathological and Clinical Implications. Cells 2022, 11, 2578. [Google Scholar] [CrossRef]
- Parisi, F.; Tesi, M.; Millanta, F.; Gnocchi, M.; Poli, A. M1 and M2 tumour-associated macrophages subsets in canine malignant mammary tumours: An immunohistochemical study. Res. Vet. Sci. 2021, 136, 32–38. [Google Scholar] [CrossRef]
- Brady, R.V.; Thamm, D.H. Tumor-associated macrophages: Prognostic and therapeutic targets for cancer in humans and dogs. Front. Immunol. 2023, 14, 1176807. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.N.; Rodrigues, M.A.; Gomes, D.A.; Salgado, B.S.; Cassali, G.D. Tumour-associated macrophages: Relation with progression and invasiveness, and assessment of M1/M2 macrophages in canine mammary tumours. Vet. J. 2018, 234, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Seung, B.J.; Lim, H.Y.; Shin, J., II; Kim, H.W.; Cho, S.H.; Kim, S.H.; Sur, J.H. CD204-Expressing Tumor-Associated Macrophages Are Associated With Malignant, High-Grade, and Hormone Receptor-Negative Canine Mammary Gland Tumors. Vet. Pathol. 2018, 55, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Kerboeuf, M.; Haugeberg, D.A.; Olsen, T.; Sørling, L.K.; Koppang, E.O.; Moe, L.; Haaland, A.H. Tumor-associated macrophages in canine visceral hemangiosarcoma. Vet. Pathol. 2023, 61, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Thaiwong, T.; Sledge, D.G.; Collins-Webb, A.; Kiupel, M. Immunohistochemical Characterization of Canine Oral Papillary Squamous Cell Carcinoma. Vet. Pathol. 2018, 55, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Nair, M.G. Macrophages in wound healing: Activation and plasticity. Immunol. Cell Biol. 2019, 97, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.L.; Klemm, F.; Akkari, L.; Tabar, V.; Gutin, P.H.; Correspondence, J.A.J. Macrophage Ontogeny Underlies Differences in Tumor-Specific Education in Brain Malignancies. Cell Rep. 2016, 17, 2445–2459. [Google Scholar] [CrossRef]
- Pierezan, F.; Mansell, J.; Ambrus, A.; Hoffmann, A.R. Immunohistochemical expression of ionized calcium binding adapter molecule 1 in cutaneous histiocytic proliferative, neoplastic and inflammatory disorders of dogs and cats. J. Comp. Pathol. 2014, 151, 347–351. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Type | Clone | Manufacturer | Antigen Retrieval | Dilution |
|---|---|---|---|---|---|
| Iba-1 | Mouse monoclonal | MABN92 | Merck Millipore (Burlington, MA USA) | HIER; Tris-EDTA; pH 9.0 | 1:100 |
| FoxP3 | Rat monoclonal | FJK-16s | eBioscience™ (# 14-5773-82) (San Diego, CA, USA) | HIER; Tris-EDTA; pH 9.0 | 1:100 |
| CTLA-4 | Mouse monoclonal | F-8 | Santa Cruz Biotechnology (Dallas, TX, USA) | HIER; Tris-EDTA; pH 9.0 | 1:100 |
| CD3 | Rabbit polyclonal | - | Dako (A0452) (Santa Clara, CA, USA) | HIER; Tris-EDTA; pH 9.0 | 1:200 |
| CD20 | Rabbit polyclonal | - | Thermo Scientific (RB-9013) (Waltham, MA, USA) | No AR | 1:200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brachelente, C.; Torrigiani, F.; Porcellato, I.; Drigo, M.; Brescia, M.; Treggiari, E.; Ferro, S.; Zappulli, V.; Sforna, M. Tumor Immune Microenvironment and Its Clinicopathological and Prognostic Associations in Canine Splenic Hemangiosarcoma. Animals 2024, 14, 1224. https://doi.org/10.3390/ani14081224
Brachelente C, Torrigiani F, Porcellato I, Drigo M, Brescia M, Treggiari E, Ferro S, Zappulli V, Sforna M. Tumor Immune Microenvironment and Its Clinicopathological and Prognostic Associations in Canine Splenic Hemangiosarcoma. Animals. 2024; 14(8):1224. https://doi.org/10.3390/ani14081224
Chicago/Turabian StyleBrachelente, Chiara, Filippo Torrigiani, Ilaria Porcellato, Michele Drigo, Martina Brescia, Elisabetta Treggiari, Silvia Ferro, Valentina Zappulli, and Monica Sforna. 2024. "Tumor Immune Microenvironment and Its Clinicopathological and Prognostic Associations in Canine Splenic Hemangiosarcoma" Animals 14, no. 8: 1224. https://doi.org/10.3390/ani14081224