
Saponin Extracts Utilization as Dietary Additive in Ruminant Nutrition: A Meta-Analysis of In Vivo Studies

 ,
,  ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
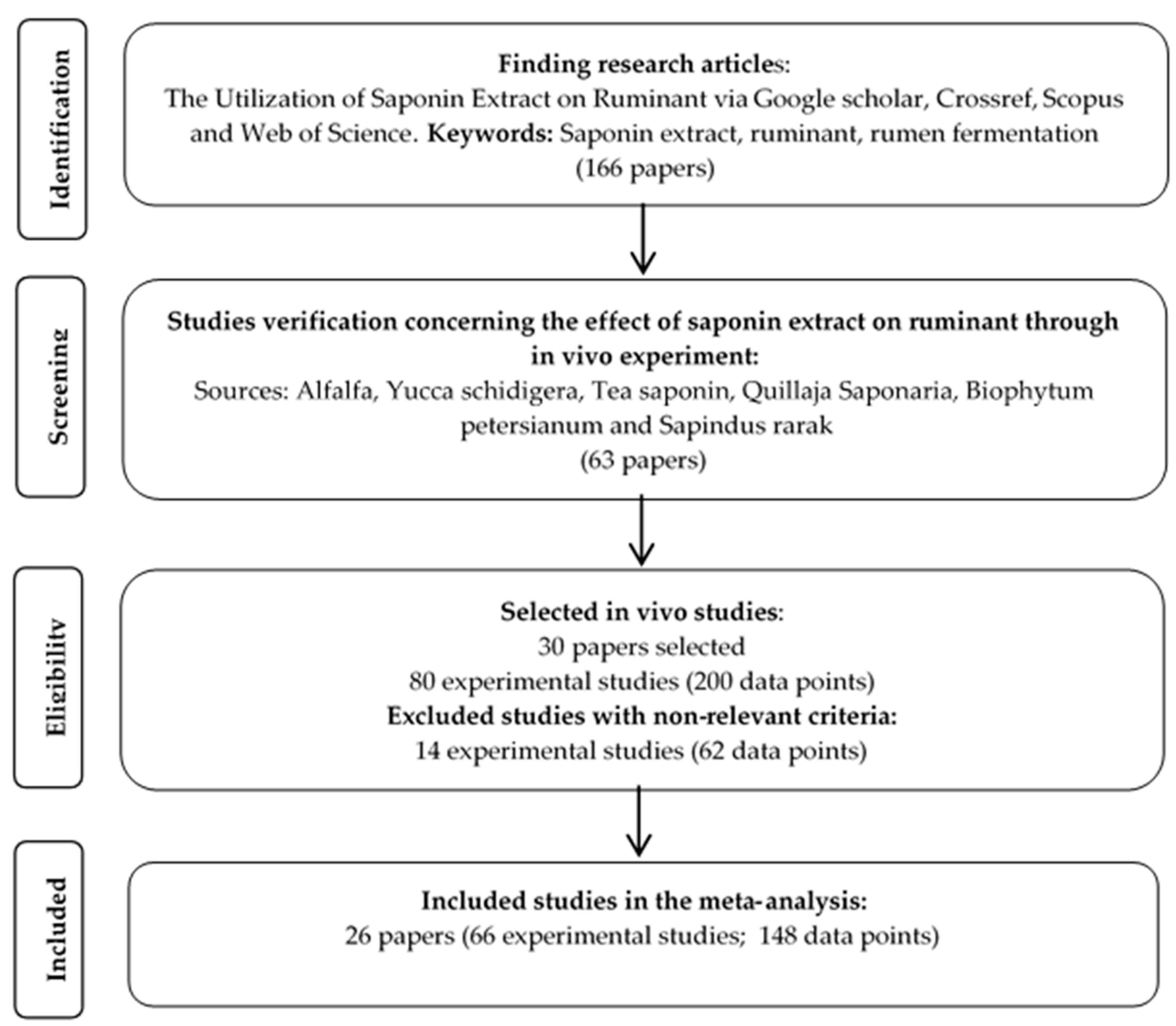
2. Materials and Methods
2.1. Database Development
2.2. Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
| No | Study | Year | Exp. | Animal | Age (Months) | Status | IBW (kg) | Sources of Saponin Extract | Level (g/kg DM) | Extracted Forms |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Valdez et al. [22] | 1986 | 1 | Dairy cows | n.d. | 1st lactation (6–10 week postpartum) | n.d. | Yucca schidigera | 0.77 | Powder |
| 2 | Lu et al. [23] | 1987 | 2–3 | Sheep | n.d. | Mature wethers | 49 | Medicago sativa | 20–40 | Powder |
| 3 | Wu et al. [24] | 1994 | 4–5 | Dairy cows | n.d. | Lactation | 650 | Yucca schidigera | 0–0.396 | Powder |
| 4 | Hussain [13] | 1995 | 6–15 | Steers | n.d. | n.d. | 574–658 | Yucca schidigera | 0.25 | Powder |
| 5 | Wilson [25] | 1998 | 16–17 | Dairy cows | Multiparous (122 d postpartum) | 640 | Yucca schidigera | 0.378 | Powder | |
| 6 | Sliwinski [26] | 2002 | 18 | Sheep | 4.02 | Castrated male lambs | 35.1 | Yucca schidigera | 0.002–0.03 | Powder |
| 7 | Eryavuz abd Dehoroti [8] | 2004 | 19–22 | Sheep | 24–108 | n.d. | 186.6 | Yucca schidigera | 5–30 | Liquid |
| 8 | Santoso et al. [27] | 2006 | 23 | Goat | n.d. | n.d. | 20.3 | Biophytum petersianum | 0.072–0.144 | Liquid |
| 9 | Wina et al. [28] | 2006 | 24–25 | Sheep | n.d. | Male sheep | 16.5 | Sapindus rarak | 20.16–30.24 | Powder |
| 10 | Lovett et al. [29] | 2006 | 26–27 | Dairy cows | n.d. | 1st and 2nd or 3rd lactation (±39 d post calving) | 585–610 | Yucca schidigera | 1.488–4.421 | Powder |
| 11 | Baah et al. [3] | 2007 | 28 | Dairy cows | n.d. | Heifers | 601 | Quillaja saponaria | 0–8 | Powder |
| 12 | Liu et al. [30] | 2007 | 29–34 | Sheep | n.d. | Male sheep | 40 | Yucca schidigera | 0.1–0.3 | Powder |
| 13 | Abdelmawla [1] | 2008 | 35 | Buffalo | n.d. | 4th and 5th lactation | 591 | Quillaja saponaria | 0.052–0.052 | Liquid |
| 14 | Benchaar et al. [6] | 2008 | 36 | Dairy cows | n.d. | Lactation (87 DIM) | 730 | Yucca schidigera | 2.752 | Powder |
| 15 | Singer et al. [31] | 2008 | 37 | Dairy cows | n.d. | Late lactation (298 DIM) | 810 | Yucca schidigera | 2.01–6.23 | Powder |
| 16 | Selcuk & Tuncer [32] | 2010 | 38 | Sheep | 2–2.5 | Male lamb | 20.87–21.69 | Yucca schidigera | 0.2–0.4 | Powder |
| 17 | Li et al. [16] | 2011 | 39 | Sheep | n.d. | Male sheep | 40 | Yucca schidigera | 0.1–0.3 | Powder |
| 18 | Nasri et al. [33] | 2011 | 40–43 | Sheep | 5–6 | Male lamb | 17.8–18.8 | Quillaja saponaria | 0.1–0.09 | Powder |
| 19 | Nasri et al. [34] | 2012 | 44–55 | Sheep | 5–6 | Female lamb | 23.9–28.9 | Quillaja saponaria | 0.12–0.36 | Liquid |
| Agave americana | 0.12–0.36 | Powder | ||||||||
| 20 | Guyader et al. [9] | 2015 | 56–57 | Dairy cows | n.d. | Multiparous nonlactating | 658 | Camellia sinensis | 5 | Powder |
| 21 | Guyader et al. [10] | 2017 | 58 | Dairy cows | Primiparous & | 617 | Camellia sinensis | 7.6 | Powder | |
| Multiparous (106 DIM) | ||||||||||
| 22 | Baheg et al. [4] | 2017 | 59–60 | Sheep | 43 | Ewes | 33.76 | Yucca schidigera | 0.2 | Powder |
| 23 | Kumar et al. [14] | 2017 | 61–63 | Goat | 7.03 | Male kids | 19.43–19.96 | Camellia sinensis | 4 | Powder |
| 25 | Zhang et al. [35] | 2021 | 65 | Sheep | 12 | Male castrated sheep | 48.37 | Camellia sinensis | 5–20 | Powder |
| 24 | Yi et al. [36] | 2022 | 64 | Steers | n.d. | Steers | 510.5 | Yucca schidigera | 0.198 | Powder |
| 26 | Alsubait et al. [2] | 2023 | 66 | Sheep | 3 and 4 | Male lambs | 26.26–26.97 | Yucca schidigera | 0.3–0.6 | Powder |
3. Results
3.1. Datasets
3.2. Relationship between Dietary Saponins Levels on Observed Parameters
3.3. Comparative Analysis of Saponins’ Sources
4. Discussion
4.1. Influence of Saponin Extract Utilization on Ruminant Performance, Digestibility, Rumen Fermentation and Health Parameters
4.2. The Relationship between Ruminant Production Health and Metabolism by the Divergence Sources of Saponin Extract
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdelmawla, S. Impacts of Natural Saponin Plant Extract without or with Fresh Baker’s Yeast in Buffaloe Heifers Rations on Nutrients Digestibility, Some Ruminal Fermentation and Blood Serum Parameters. J. Anim. Poult. Prod. 2008, 33, 3303–3315. [Google Scholar] [CrossRef]
- Alsubait, I.S.; Alhidary, I.A.; Al-Haidary, A.A. Effects of Different Levels of Yucca Supplementation on Growth Rates, Metabolic Profiles, Fecal Odor Emissions, and Carcass Traits of Growing Lambs. Animals. 2023, 13, 755. [Google Scholar] [CrossRef] [PubMed]
- Baah, J.; Ivan, M.; Hristov, A.N.; Koenig, K.M.; Rode, L.M.; McAllister, T.A. Effects of potential dietary antiprotozoal supplements on rumen fermentation and digestibility in heifers. Anim. Feed Sci. Technol. 2007, 137, 126–137. [Google Scholar] [CrossRef]
- Baheg, R.; El-Bahrawy, K.; El-Azrak, K.; Samak, M.; Sallam, S. Effect of Condensed Tannins and Saponin Supplementation on Reproductive Performance in Barki Ewes. Egypt J. Nutr. Feeds. 2017, 20, 197–210. [Google Scholar] [CrossRef]
- Benchaar, C.; Chouinard, P.Y. Short communication: Assessment of the potential of cinnamaldehyde, condensed tannins, and saponins to modify milk fatty acid composition of dairy cows. J. Dairy Sci. 2009, 92, 3392–3396. [Google Scholar] [CrossRef] [PubMed]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, ruminal fermentation, ciliate protozoal populations, and milk production from dairy cows fed cinnamaldehyde, quebracho condensed tannin, or Yucca schidigera saponin extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef]
- Cieślak, A.; Szumacher-Strabel, M.; Stochmal, A.; Oleszek, W. Plant components with specific activities against rumen methanogens. Animal 2013, 7, 253–265. [Google Scholar] [CrossRef]
- Eryavuz, A.; Dehority, B.A. Effect of Yucca schidigera extract on the concentration of rumen microorganisms in sheep. Anim. Feed Sci. Technol. 2004, 117, 215–222. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Doreau, M.; Morgavi, D.P.; Gérard, C.; Loncke, C.; Martin, C. Nitrate but not tea saponin feed additives reduce methane in dairy cows. J. Anim. Sci. 2015, 93, 5367–5377. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Doreau, M.; Morgavi, D.P.; Gérard, C.; Martin, C. Tea saponin reduced methanogenesis in vitro but increased methane yield in lactating dairy cows. J. Dairy Sci. 2017, 100, 1845–1855. [Google Scholar] [CrossRef]
- Gunun, P.; Cherdthong, A.; Khejornsart, P.; Wanapat, M.; Polyorach, S.; Kang, S.; Kaewwongsa, W.; Gunun, N. The Effect of Phytonutrients in Terminalia chebula Retz. on Rumen Fermentation Efficiency, Nitrogen Utilization, and Protozoal Population in Goats. Animals 2022, 12, 2022. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.L.; Liu, J.X.; Ye, J.A.; Wu, Y.M.; Guo, Y.Q. Effect of tea saponin on rumen fermentation in vitro. Anim. Feed Sci. Technol. 2005, 120, 333–339. [Google Scholar] [CrossRef]
- Hussain, I.; Cheeke, P.R. Effect of dietary Yucca schidigera extract on rumen and blood profiles of steers fed concentrate- or roughage-based diets. Anim. Feed Sci. Technol. 1995, 51, 231–242. [Google Scholar] [CrossRef]
- Jayanegara, A.; Sujamoko, T.U.P.; Ridla, M.; Kondo, M.; Kreuzer, M. Silage quality as influenced by concentration and type of tannins present in the material ensiled: A meta-analysis. J. Anim. Physiol. Anim. Nutr. 2019, 103, 456–465. [Google Scholar] [CrossRef]
- Kumar, M.; Kannan, A.; Bhar, R.; Gulati, A.; Gaurav, A.; Sharma, V.K. Nutrient intake, digestibility and performance of Gaddi kids supplemented with tea seed or tea seed saponin extract. Asian-Australas. J. Anim. Sci. 2017, 30, 486–494. [Google Scholar] [CrossRef]
- Li, Z.; Liu, D.; Ma, H.; Fu, B.; Wang, L.; Gou, Z.; Sun, J.; He, X.; Liu, C. Effects of Yucca on the sheep ruminal fermentation and some serum parameters. In Proceedings of the ITME 2011—IEEE International Symposium on IT in Medicine and Education, Cuangzhou, China, 9–11 December 2011; Volume 2, pp. 541–545. [Google Scholar] [CrossRef]
- Yanza, Y.R.; Mahmudah, N.; Hidayat, C.; Jayanegara, A.; Fitri, A.; Ramadhani, F.; Syarifah, H.; Basri Jumin, H.; Andrian, D.; Melissa; et al. Tea Saponin Modulates In Vitro Rumen Fermentation Profile and Reduces Methane Production: A Meta-Analysis. Biotropia 2023, 30, 82–90. [Google Scholar] [CrossRef]
- Respati, A.N.; Yanza, Y.R.; Yano, A.A.; Astuti, D.; Ningsih, N.; Triswanto; Purnamayanti, L.; Gading, B.M.; Wardani, W.W.; Jayanegara, A.; et al. Meta-analysis of the effects of dietary sources of selenium on lactational performance and oxidative status of dairy cows. Anim. Feed Sci. Technol. 2023, 305, 115782. [Google Scholar] [CrossRef]
- Yanza, Y.R.; Fitri, A.; Suwignyo, B.; Elfahmi; Hidayatik, N.; Kumalasari, N.R.; Irawan, A.; Jayanegara, A. The Utilisation of Tannin Extract as a Dietary Additive in Ruminant Nutrition: A Meta-Analysis. Animals 2021, 11, 3317. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, N.R. Invited review: Integrating quantitative findings from multiple studies using mixed model methodology. J. Dairy Sci. 2001, 84, 741–755. [Google Scholar] [CrossRef]
- Yanza, Y.R.; Szumacher-Strabel, M.; Jayanegara, A.; Kasenta, A.M.; Gao, M.; Huang, H.; Patra, A.K.; Warzych, E.; Cieslak, A. The Effects of Dietary Medium-Chain Fatty Acids on Ruminal Methanogenesis and Fermentation In Vitro and In Vivo: A Meta-analysis. J. Anim. Physiol. Anim. Nutr. 2021, 105, 874–889. [Google Scholar] [CrossRef]
- Valdez, F.R.; Bush, L.J.; Goetsch, A.L.; Owens, F.N. Effect of steroidal sapogenins on ruminal fermentation and on production of lactating dairy cows. J. Dairy Sci. 1986, 69, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.D.; Jorgensen, N.A. Alfafa saponins affect site and extent of nutrient digestion in ruminants. J. Nutr. 1987, 117, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sadik, M.; Sleiman, F.T.; Simas, J.M.; Pessarakli, M.; Hubers, J.T. Influence of yucca extract on ruminal metabolism in cows. J. Anim. Sci. 1994, 72, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.C.; Overton, T.R.; Clark, J.H. Effects of Yucca shidigera extract and soluble protein on performance of cows and concentrations of urea nitrogen in plasma and milk. J. Dairy Sci. 1998, 81, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Sliwiński, B.J.; Kreuzer, M.; Wettstein, H.R.; Machmuller, A. Rumen fermentation and nitrogen balance of lambs fed diets containing plant extracts rich tannins and saponins, and associated emissions of nitrogen and methane. Anim. Nutr. 2002, 56, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Santoso, B.; Kilmaskossu, A.; Sambodo, P. Effects of saponin from Biophytum petersianum Klotzsch on ruminal fermentation, microbial protein synthesis and nitrogen utilization in goats. Anim. Feed Sci. Technol. 2006, 137, 58–68. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. Effects of daily and interval feeding of Sapindus rarak saponins on protozoa, rumen fermentation parameters and digestibility in sheep. Asian-Australas. J. Anim. Sci. 2006, 19, 1580–1587. [Google Scholar] [CrossRef]
- Lovett, D.K.; Stack, L.; Lovell, S.; Callan, J.; Flynn, B.; Hawkins, M.; Mara, F.P.O. Effect of feeding Yucca schidigera extract on performance of lactating dairy cows and ruminal fermentation parameters in steers. Livest. Sci. 2006, 102, 23–32. [Google Scholar] [CrossRef]
- Liu, C.; Li, Z.; Juan, D.; An-shan, S. The Effect of Yucca schidigera extract on ruminal fermentation and parameters traits in sheep. J. Agr. Sci. 2007, 6, 121–128. [Google Scholar] [CrossRef]
- Singer, M.D.; Robinson, P.H.; Salem, A.Z.M.; Depeters, E.J. Impacts of rumen fluid modified by feeding Yucca schidigera to lactating dairy cows on in vitro gas production of 11 common dairy feedstuffs, as well as animal performance. Anim. Feed Sci. Technol. 2008, 146, 242–258. [Google Scholar] [CrossRef]
- Selcuk, Z.; Tuncer, S.D. The effects of different levels of Yucca schidigera added to the lamb’s diets containing urea on growth performance, carcass characteristic, some rumen, and blood parameters. Anim. Veter Adv. 2010, 9, 654–660. [Google Scholar] [CrossRef]
- Nasri, S.; Ben Salem, H.; Vasta, V.; Abidi, S.; Makkar, H.; Priolo, A. Effect of increasing levels of Quillaja saponaria on digestion, growth, meat quality of barbarine lamb. Anim. Feed Sci. Technol. 2011, 164, 71–78. [Google Scholar] [CrossRef]
- Nasri, S.; Ben Salem, H. Effect of oral administration of Agave americana or Quillaja saponaria extracts on digestion and growth of barbarine female lamb. Livest. Sci. 2012, 147, 59–65. [Google Scholar] [CrossRef]
- Zhang, F.; Li, B.; Ban, Z.; Liang, H.; Li, L.; Zhao, W.; Yan, X. Evaluation of origanum oil, hydrolysable tannins and tea saponin in mitigating ruminant methane: In vitro and In vivo methods. Anim. Physiol. Anim. Nutr. 2021, 105, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Wu, B.; Ma, J.; Cui, X.; Deng, Z.; Hu, S.; Li, W.; Runa, A.; Li, X.; Meng, Q.; et al. Effects of dietary capsaicin and Yucca schidigera extracts as feed additives on rumen fermentation and microflora of beef cattle fed with a moderate-energy diet. J. Ferment. 2022, 9, 30. [Google Scholar] [CrossRef]
- Wang, J.K.; Ye, J.A.; Liu, J.X. Effects of tea saponins on rumen microbiota, rumen fermentation, methane production and growth performance—A review. Trop. Anim. Health Prod. 2012, 44, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Pen, B.; Sar, C.; Mwenya, B.; Kuwaki, K.; Morikawa, R.; Takahashi, J. Effects of Yucca schidigera and Quillaja saponaria extracts on in vitro ruminal fermentation and methane emission. Anim. Feed Sci. Technol. 2006, 129, 175–186. [Google Scholar] [CrossRef]
- Wang, B.; Tu, Y.; Zhao, S.P.; Hao, Y.H.; Liu, J.X.; Liu, F.H.; Xiong, B.H.; Jiang, L.S. Effect of tea saponins on milk performance, milk fatty acids, and immune function in dairy cow. J. Dairy Sci. 2017, 100, 8043–8052. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The impact of saponins or saponin-containing plant materials on ruminant production--a review. J. Agric. Food Chem. 2005, 53, 8093–8105. [Google Scholar] [CrossRef]
- Zhou, L.; Shi, Y.; Guo, R.; Liang, M.; Zhu, X.; Wang, C. Digital Gene-Expression Profiling Analysis of the Cholesterol-Lowering Effects of Alfalfa Saponin Extract on Laying Hens. PLoS ONE 2014, 9, e98578. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. “Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rumin. Res. 2015, 64, 157–166. [Google Scholar] [CrossRef]
- Kamra, D.N. Rumen microbial ecosystem. Curr. Sci. 2011, 101, 845–858. Available online: http://www.jstor.org/stable/24110438 (accessed on 29 March 2024).
- Clauss, M.; Hume, I.D.; Hummel, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 2010, 8, 979–992. [Google Scholar] [CrossRef]
- Mao, H.L.; Wang, J.K.; Zhou, Y.Y.; Liu, J.X. Effects of addition of tea saponins and soybean oil on milk performance, milk fatty acid composition, and nutrient digestibility in dairy cows. Livest. Sci. 2010, 129, 56–62. [Google Scholar] [CrossRef]
- Tavendale, M.H.; Meagher, L.P.; Pacheco, D.; Walker, N.; Attwood, G.T.; Sivakumaran, S. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Technol. 2005, 123–124, 403–419. [Google Scholar] [CrossRef]
- Sophie Comtet-Marre, N.; Parisot, P.; Lepercq, F.; Chaucheyras-Durand, P.; Mosoni, E.; Peyretaillade, A.; Bayat, K.; Shingfield, P.P.; Forano, E. Metatranscriptomics Reveals the Active Bacterial and Eukaryotic Fibrolytic Communities in the Rumen of Dairy Cow Fed a Mixed Diet. Front. Microbiol. 2017, 8, 67. [Google Scholar] [CrossRef]
- Anantasook, N.; Wanapat, M.; Cherdthong, A.; Gunun, P. Effect of tannins and saponins in Samanea saman on rumen environment, milk yield and milk composition in lactating dairy cows. J. Anim. Physiol. Anim. Nutr. 2015, 99, 335–344. [Google Scholar] [CrossRef]
- Kozłowska, M.; Cieślak, A.; Jóźwik, A.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Kowalczyk, M.; Filipiak, W.; Szumacher-Strabel, M. The effect of total and individual alfalfa saponins on rumen methane production. J. Sci. Food Agric. 2020, 100, 1922–1930. [Google Scholar] [CrossRef]
- Kozłowska, M.; Cieślak, A.; Jóźwik, A.; El-Sherbiny, M.; Gogulski, M.; Lechniak, D.; Gao, M.; Yanza, Y.R.; Vazirigohar, M.; Szumacher-Strabel, M. Effects of partially replacing grass silage by lucerne silage cultivars in a high-forage diet on ruminal fermentation, methane production, and fatty acid composition in the rumen and milk of dairy cows. Anim. Feed Sci. Technol. 2021, 277, 114959. [Google Scholar] [CrossRef]
- Patra, A.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef]
- Lu, C.; Fan, G.; Wang, D. Akebia Saponin D ameliorated kidney injury and exerted anti-inflammatory and anti-apoptotic effects in diabetic nephropathy by activation of NRF2/HO-1 and inhibition of NF-KB pathway. Int. Immunopharmacol. 2020, 84, 106467. [Google Scholar] [CrossRef]
- Du, Y.; Wang, L.; Qian, J.; Zhang, K.; Chai, K. Panax notoginseng saponins protect kidney from diabetes by up-regulating silent information regulator 1 and activating antioxidant proteins in rats. Chin. J. Integr. Med. 2016, 22, 910–917. [Google Scholar] [CrossRef]
- Chen, G.; Wu, S.; Na, S.; Li, L. Effect of total saponin of dioscorea on uric acid excretion indicators in chronic hyperuricemia rats. Chin. J. Integr. Tradit. West. Med. 2014, 341, 75–80. [Google Scholar]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review. Brit. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.; Diao, Q.; Tu, Y. Saponin-Induced Shifts in the Rumen Microbiome and Metabolome of Young Cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef]
- Constable, P.D.; Hinchcliff, K.W.; Done, S.H.; Grünberg, W. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs, and Goats, 11th ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Thrall, M.A.; Weiser, G.; Allison, R.W.; Campbell, T.W. Veterinary Hematology and Clinical Chemistry; Wiley-Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]



| Parameters | Unit | N | Mean | SEM | Min | Max |
|---|---|---|---|---|---|---|
| Feed Intake | ||||||
| DMI | g/kg BW0.75 | 67 | 113.1 | 4.917 | 50.49 | 187.0 |
| OMI | g/kg BW0.75 | 12 | 80.25 | 9.159 | 48.82 | 150.2 |
| NDFI | g/kg BW0.75 | 12 | 36.22 | 3.146 | 18.12 | 56.62 |
| Gain Performance | ||||||
| ADG | g/d | 18 | 150.8 | 20.20 | 59.6 | 275.0 |
| g/kg BW0.75 | 18 | 13.18 | 2.209 | 5.25 | 27.80 | |
| Milk production and composition | ||||||
| Milk yield | kg/d | 21 | 25.23 | 2.071 | 7.01 | 33.85 |
| g/kg BW0.75 | 19 | 194.0 | 16.33 | 58.48 | 264.8 | |
| g/kg DMI | 21 | 1.20 | 0.080 | 0.46 | 1.55 | |
| Milk fat | g/kg | 21 | 44.49 | 3.103 | 31.3 | 74.6 |
| Milk protein | g/kg | 17 | 34.22 | 1.358 | 28.0 | 47.9 |
| Milk lactose | g/kg | 12 | 47.56 | 0.785 | 43.8 | 52.2 |
| Digestibility | ||||||
| DMD | g/kg | 40 | 698.3 | 8.769 | 628 | 813.0 |
| OMD | g/kg | 57 | 683.9 | 11.47 | 405 | 827.0 |
| CPD | g/kg | 44 | 636.2 | 10.46 | 531 | 793.7 |
| NDFD | g/kg | 47 | 559.0 | 14.89 | 295 | 730.3 |
| ADFD | g/kg | 27 | 489.9 | 15.79 | 323 | 660.1 |
| Rumen fermentation parameters | ||||||
| pH | 148 | 6.30 | 0.030 | 5.51 | 7.1 | |
| NH3 | mg/dL | 146 | 18.77 | 1.028 | 4.12 | 81.74 |
| Total VFA | mmol/L | 74 | 103.7 | 14.21 | 48.9 | 125.0 |
| Acetate | % | 71 | 62.23 | 1.305 | 5.73 | 78.4 |
| Propionate | % | 71 | 22.52 | 0.835 | 3.14 | 41.74 |
| Butyrate | % | 71 | 11.65 | 0.432 | 1.17 | 19.3 |
| Valerate | % | 29 | 1.27 | 0.141 | 0.16 | 3.68 |
| A:P ratio | 71 | 3.10 | 0.151 | 0.79 | 7.47 | |
| Protozoa | log10/mL | 91 | 5.42 | 0.044 | 4.19 | 6.52 |
| N Balance | ||||||
| Urine N/MBW | g/kg BW0.75 | 35 | 585.4 | 51.2 | 174.7 | 1099 |
| Fecal N/MBW | g/kg BW0.75 | 35 | 550.2 | 46.5 | 199.8 | 1494 |
| N retention/MBW | g/kg BW0.75 | 23 | 463.6 | 49.8 | 52.3 | 759.8 |
| Blood parameters | ||||||
| Plasma NH3 | µg/dL | 40 | 1.90 | 0.386 | 0.64 | 10.85 |
| Total protein | g/dL | 20 | 6.97 | 0.255 | 5.32 | 10.31 |
| Albumin | g/dL | 13 | 3.01 | 0.105 | 2.55 | 3.84 |
| Globulin | g/dL | 10 | 3.72 | 0.158 | 2.97 | 4.66 |
| PUN | mg/dL | 72 | 17.22 | 1.20 | 3.59 | 42.0 |
| Cholesterol | g/dL | 21 | 75.43 | 8.087 | 28.0 | 153.7 |
| Creatinine | μmol/L | 28 | 80.0 | 3.814 | 35.4 | 106.7 |
| Glucose | mg/dL | 20 | 55.17 | 3.524 | 30.96 | 86.0 |
| ALP | IU/L | 10 | 146.6 | 19.837 | 103.7 | 272.9 |
| Estimated Variables | Unit | N | Model | Intercept | SEintercept | Slope | SEslope | p-Value | RMSE | AIC | L × Animal | L × Source |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Feed Intake | ||||||||||||
| DMI | g/kg BW0.75 | 67 | L | 112.1 | 7.822 | −0.015 | 0.103 | 0.889 | 3.676 | 519.5 | 0.016 | 0.660 |
| OMI | g/kg BW0.75 | 12 | L | 80.85 | 13.59 | −0.216 | 0.725 | 0.778 | 4.728 | 97.8 | 0.079 | 0.005 |
| NDFI | g/kg BW0.75 | 12 | L | 36.32 | 4.690 | −0.036 | 0.288 | 0.905 | 1.878 | 77.98 | 0.032 | 0.021 |
| Gain performance | ||||||||||||
| ADG | g/d | 18 | L | 170.8 | 54.36 | 0.183 | 0.538 | 0.739 | 96.11 | 143 | 0.739 | 0.021 |
| g/kg BW0.75 | 18 | L | 12.01 | 4.198 | 0.009 | 0.061 | 0.880 | 0.705 | 83.1 | 0.009 | 0.854 | |
| Milk production and composition | ||||||||||||
| Milk yield | kg/d | 21 | L | 25.98 | 3.932 | −0.364 | 0.109 | 0.005 | 11.31 | 93.9 | 0.016 | 0.001 |
| g/kg BW0.75 | 19 | L | 211.6 | 29.18 | −2.938 | 0.912 | 0.008 | 5.339 | 158.4 | <0.001 | 0.295 | |
| g/kg DMI | 21 | L | 1.253 | 0.134 | −0.002 | 0.005 | 0.698 | 0.030 | −21.1 | <0.001 | 0.220 | |
| Milk fat | g/kg | 21 | L | 42.48 | 5.761 | 0.257 | 0.233 | 0.290 | 77.10 | 118 | 0.001 | 0.001 |
| Milk protein | g/kg | 17 | L | 33.70 | 2.485 | −0.091 | 0.384 | 0.817 | 79.74 | 92.5 | 0.035 | 0.082 |
| Milk lactose | g/kg | 12 | L | 47.59 | 1.567 | 0.005 | 0.225 | 0.981 | 36.44 | 56 | 0.197 | 0.341 |
| Digestibility | ||||||||||||
| DMD | g/kg | 40 | L | 702.0 | 17.07 | 0.973 | 0.473 | 0.048 | 31.45 | 382 | 0.015 | 0.425 |
| OMD | g/kg | 57 | L | 687.2 | 26.60 | 0.373 | 0.371 | 0.320 | 59.26 | 543 | 0.002 | 0.359 |
| CPD | g/kg | 44 | L | 640.2 | 24.20 | 2.017 | 1.546 | 0.201 | 36.40 | 439 | 0.206 | 0.568 |
| NDFD | g/kg | 47 | L | 564.8 | 32.90 | −1.345 | 0.806 | 0.104 | 48.43 | 471 | 0.064 | 0.795 |
| ADFD | g/kg | 27 | L | 500.7 | 33.10 | −3.977 | 1.764 | 0.036 | 39.99 | 275 | 0.005 | 0.235 |
| Rumen Fermentation Profile | ||||||||||||
| pH | 148 | L | 6.301 | 0.068 | −0.0004 | 0.005 | 0.945 | 0.176 | 114 | 0.793 | 0.394 | |
| NH3 | mg/dL | 146 | L | 23.11 | 3.102 | −0.116 | 0.110 | 0.293 | 10.46 | 982 | 0.361 | 0.653 |
| Total VFA | mmol/L | 74 | Q | 107.9 | 25.50 | 18.44 | 6.010 | 0.003 | 78.17 | 897 | <0.001 | <0.001 |
| −0.440 | 0.150 | |||||||||||
| Acetate | % | 71 | Q | 62.59 | 3.010 | −0.640 | 0.320 | 0.049 | 6.880 | 494 | 0.001 | <0.001 |
| 0.010 | 0.008 | |||||||||||
| Propionate | % | 71 | L | 21.17 | 1.848 | −0.059 | 0.067 | 0.382 | 4.440 | 416 | 0.001 | <0.001 |
| Butyrate | % | 71 | L | 11.28 | 0.943 | −0.031 | 0.042 | 0.463 | 2.014 | 348 | 0.038 | 0.001 |
| Valerate | % | 29 | Q | 1.370 | 0.260 | 0.190 | 0.080 | 0.001 | 0.355 | 32.5 | <0.001 | 0.001 |
| L | −0.040 | 0.010 | ||||||||||
| A:P ratio | 71 | L | 3.321 | 0.358 | 0.002 | 0.008 | 0.826 | 0.796 | 140 | 0.949 | 0.786 | |
| Protozoa | log10/mL | 91 | Q | 5.390 | 0.130 | 0.020 | 0.010 | 0.030 | 0.939 | 38.6 | 0.689 | <0.001 |
| −0.001 | 0.000 | |||||||||||
| N Partitioning | ||||||||||||
| Urine N | g/kg BW0.75 | 35 | Q | 614.1 | 83.99 | −5.524 | 3.315 | 0.057 | 38.02 | 430.9 | 0.371 | 0.934 |
| 0.173 | 0.085 | |||||||||||
| Fecal N | g/kg BW0.75 | 35 | L | 573.96 | 70.65 | −0.462 | 0.878 | 0.605 | 31.28 | 417.7 | 0.009 | 0.959 |
| N retention | g/kg BW0.75 | 23 | L | 476.43 | 83.87 | −0.914 | 2.207 | 0.686 | 40.00 | 272 | - | 0.976 |
| Blood parameters | ||||||||||||
| Total protein | g/dL | 20 | L | 6.790 | 0.498 | −0.027 | 0.255 | 0.915 | 1.261 | 59.2 | 0.562 | 0.993 |
| Albumin | g/dL | 13 | L | 2.976 | 0.222 | 0.044 | 0.050 | 0.400 | 3.484 | 6.8 | 0.156 | 0.329 |
| Globulin | g/dL | 10 | L | 3.756 | 0.358 | −0.057 | 0.096 | 0.576 | 7.530 | 15.7 | 0.243 | 0.226 |
| PUN | mg/dL | 72 | L | 18.23 | 2.317 | 0.105 | 0.559 | 0.852 | 1.426 | 378.5 | 0.156 | 0.535 |
| Cholesterol | mg/dL | 21 | L | 86.18 | 23.49 | −1.849 | 3.145 | 0.565 | 94.65 | 160 | 0.679 | 0.055 |
| Creatinine | μmol/L | 28 | Q | 77.54 | 7.510 | 51.85 | 19.62 | 0.014 | 58.79 | 218 | 0.884 | <0.001 |
| −13.37 | 4.960 | |||||||||||
| Glucose | mg/dL | 20 | L | 49.85 | 8.310 | −1.027 | 3.707 | 0.785 | 115.6 | 153 | 0.785 | 0.083 |
| ALP | IU/L | 10 | L | 166.8 | 37.75 | 0.514 | 2.830 | 0.861 | 98.59 | 76.9 | 0.861 | 0.065 |
| Estimated Variables | Unit | n | Experimental Groups | SEM | p-Value | Source × Animal | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | MS | QS | CS | YS | AA | BP | SR | ||||||
| Feed Intake | |||||||||||||
| DMI | g/kg BW0.75 | 67 | 112.8 | 113.6 | 111.1 | 112.4 | 111.2 | - | - | - | 4.92 | 0.892 | 0.015 |
| OMI | g/kg BW0.75 | 12 | 84.92 | - | 57.26 | 84.77 | - | - | - | - | 9.16 | 0.776 | 0.043 |
| NDFI | g/kg BW0.75 | 12 | 39.53 | - | 18.54 | 39.99 | - | - | - | - | 3.15 | 0.142 | 0.017 |
| Gain performance | |||||||||||||
| ADG | g/d | 18 | 171.1 ab | - | 188.0 a | 169.9 ab | 164.8 b | 189.4 a | - | - | 44.43 | 0.005 | - |
| g/kg BW0.75 | 18 | 11.707 b | - | 13.25 a | 11.63 b | 10.97 c | 13.58 a | - | - | 2.21 | 0.025 | - | |
| Milk production and composition | |||||||||||||
| Milk yield | kg/d | 21 | 25.80 a | - | 26.72 a | 20.81 b | 25.55 a | - | - | - | 2.97 | 0.001 | - |
| g/kg BW0.75 | 19 | 210.4 a | - | 218.4 a | 170.0 b | 208.0 a | - | - | - | 16.33 | 0.001 | - | |
| g/kg DMI | 21 | 1.247 | - | 1.311 | 1.155 | 1.255 | - | - | - | 0.08 | 0.146 | - | |
| Milk fat | g/kg | 21 | 41.52 b | - | 48.22 a | 44.23 ab | 42.54 b | - | - | - | 3.48 | <0.001 | - |
| Milk protein | g/kg | 17 | 32.97 | - | 39.84 | 31.84 | 32.55 | - | - | - | 3.08 | 0.084 | - |
| Milk lactose | g/kg | 12 | 46.92 | - | 50.11 | 48.08 | 46.64 | - | - | - | 2.19 | 0.339 | - |
| Digestibility | |||||||||||||
| DMD | g/kg | 40 | 700.4 | 730.5 | 717.02 | 705.7 | 697 | - | 683.4 | - | 29.41 | 0.491 | 0.014 |
| OMD | g/kg | 57 | 687.5 | 717 | 699.1 | 692.9 | 682.8 | 703.2 | 677.9 | 663.8 | 33.11 | 0.408 | 0.003 |
| CPD | g/kg | 44 | 644.7 | - | 638.1 | 645.5 | 647.3 | 673.8 | 628.2 | 38.59 | 0.658 | 0.321 | |
| NDFD | g/kg | 47 | 563.2 | - | 568.3 | 550.6 | 560.8 | 569.7 | 551.3 | 547.6 | 47.46 | 0.972 | 0.321 |
| ADFD | g/kg | 27 | 494.9 | - | - | 484 | 493.8 | - | 496 | - | 48.29 | 0.956 | 0.269 |
| Rumen fermentation parameters | |||||||||||||
| pH | 148 | 6.28 | 6.09 | 6.26 | 6.11 | 6.31 | 6.14 | 6.8 | - | 0.26 | 0.341 | 0.440 | |
| NH3 | mg/dL | 146 | 23.6 | 27.19 | 19.14 | 21.83 | 23.26 | 23.66 | 17.42 | 19.78 | 6.64 | 0.386 | 0.224 |
| Total VFA | mmol/L | 74 | 104.9 a | 60.12 b | 116.5 a | 106.9 a | 111.1 a | - | 63.79 b | - | 57.83 | <0.001 | 0.224 |
| Acetate | % | 71 | 63.34 | 64.14 | 29.99 | 66.45 | 63.75 | - | 60.36 | 66.32 | 10.27 | 0.106 | 0.798 |
| Propionate | % | 71 | 20.41 | - | 17.52 | 19.57 | 21.47 | - | 26.58 | 21.66 | - | 0.665 | 0.156 |
| Butyrate | % | 71 | 11.96 | 13.59 | 7.41 | 10.84 | 11.5 | - | 9.01 | 8.87 | 2.95 | 0.295 | 0.541 |
| Valerate | % | 29 | 1.34 | - | 0.9 | - | 1.44 | - | 1,17 | - | 0.42 | 0.782 | 0.126 |
| A:P ratio | 71 | 3.53 | 3.48 | 1.76 | 3.58 | 3.49 | - | 2.67 | 3.22 | 1.47 | 0.521 | 0.240 | |
| Protozoa | log10/mL | 91 | 5.48 a | 5.17 b | 5.29 b | 5.63 a | 5.63 a | 5.33 b | 5.22 b | 5.02 b | 0.25 | 0.009 | 0.287 |
| N Partitioning | |||||||||||||
| Urine N | g/kg BW0.75 | 35 | 619.9 | 622.8 | 521.9 | 637.1 | 615.0 | 597.5 | - | 570.84 | 51.20 | 0.614 | 0.245 |
| Fecal N | g/kg BW0.75 | 35 | 572.6 | 557.2 | 590.1 | 567.9 | 574.8 | 584.8 | - | 554.91 | 46.47 | 0.997 | 0.001 |
| N retention | g/kg BW0.75 | 23 | 463.2 | - | 537.3 | 368.5 | 486.2 | 523.5 | - | 459.46 | 49.82 | 0.188 | - |
| Blood parameters | |||||||||||||
| Total protein | g/dL | 20 | 6.66 | - | 7.4 | 6.63 | 6.2 | 7.85 | - | - | 0.95 | 0.279 | 0.689 |
| Albumin | g/dL | 13 | 2.92 | - | 3.24 | 3.1 | 2.91 | - | - | - | 0.22 | 0.308 | - |
| Globulin | g/dL | 10 | 3.64 | - | 4.20 | 3.51 | 3.32 | - | - | - | 0.35 | 0.228 | - |
| PUN | mg/dL | 72 | 18.07 b | - | 17.1335 b | 19.40 b | 17.90 b | 23.04 a | - | - | 1.20 | 0.001 | 0.345 |
| Cholesterol | mg/dL | 21 | 87.87 | - | 81.65 | 84.14 | 91.55 | 73.57 | - | - | 27.63 | 0.287 | 0.711 |
| Creatinine | μmol/L | 28 | 80.16 b | - | 87.67 b | 72.35 b | 80.14 b | 122.0 a | - | - | 12.29 | <0.001 | - |
| Glucose | mg/dL | 20 | 53.65 a | - | 31.03 b | 53.56 a | 49.90 a | 31.62 b | - | - | 13.02 | 0.005 | - |
| ALP | IU/L | 10 | 170.8 | - | - | 175.9 | 161.9 | - | - | - | 32.69 | 0.388 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanza, Y.R.; Irawan, A.; Jayanegara, A.; Ramadhani, F.; Respati, A.N.; Fitri, A.; Hidayat, C.; Niderkorn, V.; Cieslak, A.; Szumacher-Strabel, M.; et al. Saponin Extracts Utilization as Dietary Additive in Ruminant Nutrition: A Meta-Analysis of In Vivo Studies. Animals 2024, 14, 1231. https://doi.org/10.3390/ani14081231
Yanza YR, Irawan A, Jayanegara A, Ramadhani F, Respati AN, Fitri A, Hidayat C, Niderkorn V, Cieslak A, Szumacher-Strabel M, et al. Saponin Extracts Utilization as Dietary Additive in Ruminant Nutrition: A Meta-Analysis of In Vivo Studies. Animals. 2024; 14(8):1231. https://doi.org/10.3390/ani14081231
Chicago/Turabian StyleYanza, Yulianri Rizki, Agung Irawan, Anuraga Jayanegara, Fitri Ramadhani, Adib Norma Respati, Ainissya Fitri, Cecep Hidayat, Vincent Niderkorn, Adam Cieslak, Malgorzata Szumacher-Strabel, and et al. 2024. "Saponin Extracts Utilization as Dietary Additive in Ruminant Nutrition: A Meta-Analysis of In Vivo Studies" Animals 14, no. 8: 1231. https://doi.org/10.3390/ani14081231