Bacterial Community Influences the Effects of Lactobacillus acidophilus on Lipid Metabolism, Immune Response, and Antioxidant Capacity in Dogs

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. L. acidophilus
2.2. Experimental Animals and Study Design
2.3. Sample Collection
2.4. Measurement of Body Weight
2.5. Measurement of Nutrient Apparent Digestibility
2.6. Measurement of Serum Indices
2.7. Bacterial Community Analysis
2.8. Statistical Analysis
3. Results
3.1. Effects of Lactobacillus acidophilus on Body Weight in Chinese Rural Dogs
3.2. Effect of Lactobacillus acidophilus on Apparent Digestibility of Nutrients in Chinese Rural Dogs
3.3. Effect of Lactobacillus acidophilus on Serum Indices in Chinese Rural Dogs
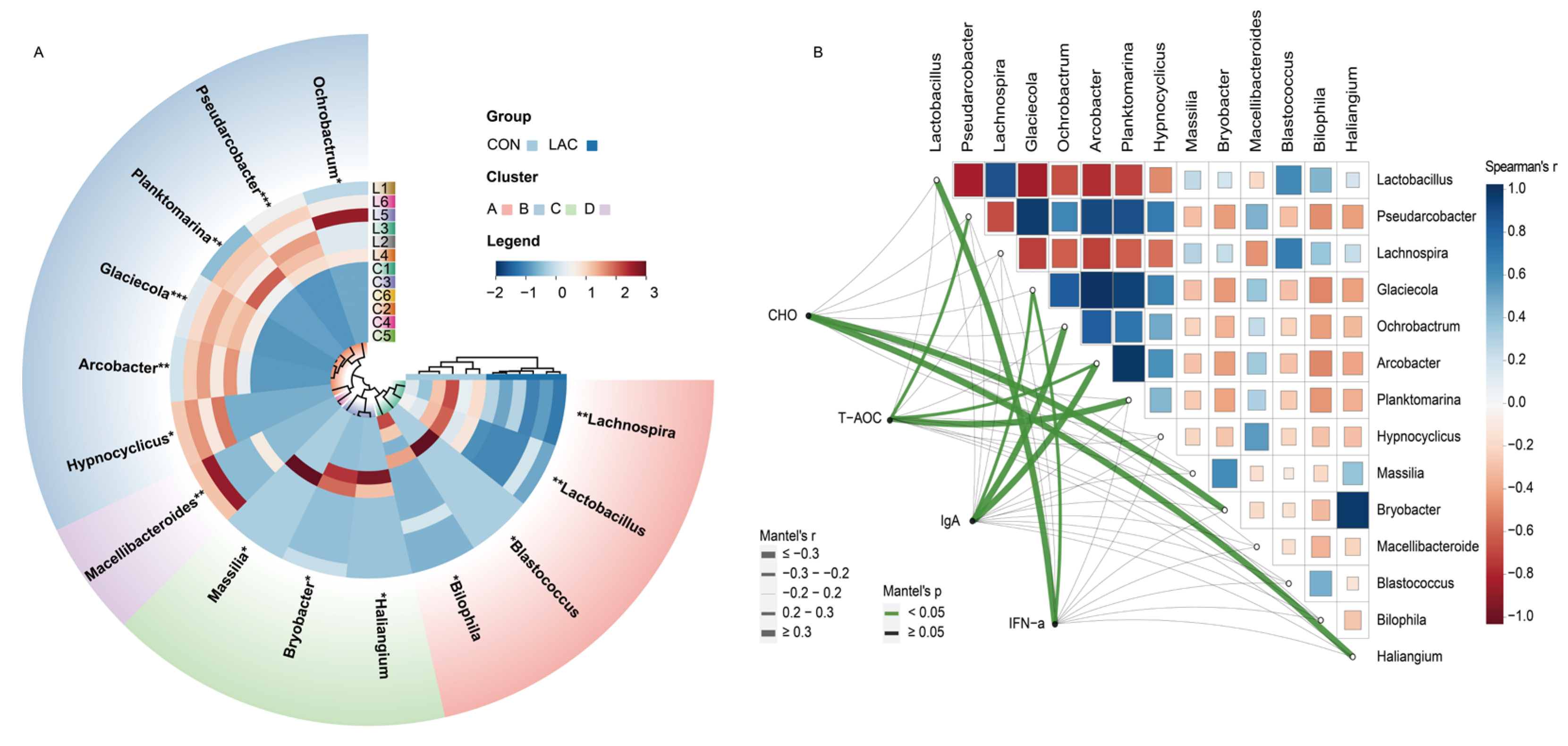
3.4. Effects of Lactobacillus acidophilus on the Bacterial Community in Chinese Rural Dogs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, H.; Li, X.; Chen, X.; Hai, D.; Wei, C.; Zhang, L.; Li, P. The Functional Roles of Lactobacillus acidophilus in Different Physiological and Pathological Processes. J. Microbiol. Biotechnol. 2022, 32, 1226–1233. [Google Scholar] [CrossRef]
- Mital, B.K.; Garg, S.K. Anticarcinogenic, hypocholesterolemic, and antagonistic activities of Lactobacillus acidophilus. Crit. Rev. Microbiol. 1995, 21, 175–214. [Google Scholar] [CrossRef]
- Theilmann, M.C.; Goh, Y.J.; Nielsen, K.F.; Klaenhammer, T.R.; Barrangou, R.; Abou Hachem, M. Lactobacillus acidophilus Metabolizes Dietary Plant Glucosides and Externalizes Their Bioactive Phytochemicals. mBio 2017, 8, 10–1128. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, B.; Zhou, X.; Wang, Y.; Wang, H.; Jia, S.; Zhang, Z.; Chu, C.; Mu, J. Combined Lowering Effects of Rosuvastatin and L. acidophilus on Cholesterol Levels in Rat. J. Microbiol. Biotechnol. 2019, 29, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.; Fontecha, J.; Cuesta, P. Combined effect of Lactobacillus acidophilus and β-cyclodextrin on serum cholesterol in pigs. Br. J. Nutr. 2016, 115, 1–5. [Google Scholar] [CrossRef]
- Lee, N.Y.; Shin, M.J.; Youn, G.S.; Yoon, S.J.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Han, S.H.; Kim, B.K.; Lee, D.Y.; et al. Lactobacillus attenuates progression of nonalcoholic fatty liver disease by lowering cholesterol and steatosis. Clin. Mol. Hepatol. 2021, 27, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Kang, X.; Yang, H.; Liu, H.; Yang, X.; Liu, Q.; Tian, H.; Xue, Y.; Ren, P.; Kuang, X.; et al. Lactobacillus acidophilus ameliorates obesity in mice through modulation of gut microbiota dysbiosis and intestinal permeability. Pharmacol. Res. 2022, 175, 106020. [Google Scholar] [CrossRef]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef]
- Sandøe, P.; Palmer, C.; Corr, S.; Astrup, A.; Bjørnvad, C.R. Canine and feline obesity: A One Health perspective. Vet. Rec. 2014, 175, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Hamper, B. Current Topics in Canine and Feline Obesity. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 785–795. [Google Scholar] [CrossRef]
- Clark, M.; Hoenig, M. Metabolic Effects of Obesity and Its Interaction with Endocrine Diseases. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 797–815. [Google Scholar] [CrossRef]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the dog and human gut microbiomes in gene content and response to diet. Microbiome 2018, 19, 72. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Endo, A.; Beasley, S.; Salminen, S. Microbiota and probiotics in canine and feline welfare. Anaerobe 2015, 34, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Tvarijonaviciute, A.; Barić-Rafaj, R.; Horvatic, A.; Muñoz-Prieto, A.; Guillemin, N.; Lamy, E.; Tumpa, A.; Ceron, J.J.; Martinez-Subiela, S.; Mrljak, V. Identification of changes in serum analytes and possible metabolic pathways associated with canine obesity-related metabolic dysfunction. Vet. J. 2019, 244, 51–59. [Google Scholar] [CrossRef]
- Chikamune, T.; Katamoto, H.; Ohashi, F.; Shimada, Y. Serum lipid and lipoprotein concentrations in obese dogs. J. Vet. Med. Sci. 1995, 57, 595–598. [Google Scholar] [CrossRef]
- Pasupathy, K.; Sahoo, A.; Pathak, N.N. Effect of lactobacillus supplementation on growth and nutrient utilization in mongrel pups. Arch. Tierernahr. 2001, 55, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Baillon, M.-L.A.; Marshall-Jones, Z.V.; Butterwick, R.F. Effects of probiotic Lactobacillus acidophilus strain DSM13241 in healthy adult dogs. Am. J. Vet. Res. 2004, 65, 338–343. [Google Scholar] [CrossRef]
- Marshall-Jones, Z.V.; Baillon, M.L.; Croft, J.M.; Butterwick, R.F. Effects of Lactobacillus acidophilus DSM13241 as a probiotic in healthy adult cats. Am. J. Vet. Res. 2006, 67, 1005–1012. [Google Scholar] [CrossRef]
- Marelli, S.P.; Fusi, E.; Giardini, A.; Martino, P.A.; Polli, M.; Bruni, N.; Rizzi, R. Effects of probiotic Lactobacillus acidophilus D2/CSL (CECT 4529) on the nutritional and health status of boxer dogs. Vet. Rec. 2020, 187, e28. [Google Scholar] [CrossRef]
- National Research Council, Division on Earth, Life Studies, Committee on Animal Nutrition, Subcommittee on Dog, & Cat Nutrition. Research Council. Nutrient Requirements of Dogs and Cats; The National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Animal Feeding Stuffs: Methods of Sampling and Analysis—Detection and Enumeration of Lactobacillus spp. Used as Feed Additive. UNE EN 15787:2022. Available online: https://standards.iteh.ai/catalog/standards/cen/cd3bf18a-7ac3-47b2-bc7e-c9571525c94f/en-15787-2021 (accessed on 13 March 2024).
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists; AOAC: Arlington, VI, USA, 2003. [Google Scholar]
- Berg, J.; Brandt, K.K.; Al-Soud, W.A.; Holm, P.E.; Hansen, L.H.; Sørensen, S.J.; Nybroe, O. Selection for Cu-tolerant bacterial communities with altered composition, but unaltered richness, via long-term Cu exposure. Appl. Environ. Microbiol. 2012, 78, 7438–7446. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Alaqil, A.A.; Abbas, A.O.; El-Beltagi, H.S.; El-Atty, H.K.A.; Mehaisen, G.M.K.; Moustafa, E.S. Dietary Supplementation of Probiotic Lactobacillus acidophilus Modulates Cholesterol Levels, Immune Response, and Productive Performance of Laying Hens. Animals 2020, 10, 1588. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Sulek, K.; Skov, K.; Frandsen, H.L.; Smedsgaard, J.; Wilcks, A.; Skov, T.H.; Villas-Boas, S.G.; Licht, T.R. Lactobacillus acidophilus NCFM affects vitamin E acetate metabolism and intestinal bile acid signature in monocolonized mice. Gut Microbes 2014, 5, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Ostrowski, M.; Ferreira, D.M.T.P.; Rosado, E.L.; Soares-Mota, M. Effect of Lactobacillus on body weight and body fat in overweight subjects: A systematic review of randomized controlled clinical trials. Int. J. Obes. 2017, 41, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, K.E.; Burnett, J.R.; Hooper, A.J.; van Bockxmeer, F.M. Familial hypercholesterolemia: Epidemiology, Neolithic origins and modern geographic distribution. Crit. Rev. Clin. Lab. Sci. 2011, 48, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Abifadel, M.; Rabès, J.P.; Devillers, M.; Munnich, A.; Erlich, D.; Junien, C.; Varret, M.; Boileau, C. Mutations and polymorphisms in the proprotein convertase subtilisin kexin 9 (PCSK9) gene in cholesterol metabolism and disease. Hum. Mutat. 2009, 30, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Dubuc, G.; Seidah, N.G. The influence of PCSK9 polymorphisms on serum low-density lipoprotein cholesterol and risk of atherosclerosis. Curr. Atheroscler. Rep. 2010, 12, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Seidah, N.G.; Awan, Z.; Chrétien, M.; Mbikay, M. PCSK9: A key modulator of cardiovascular health. Circ. Res. 2014, 114, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Park, S.; Lee, H.; Min, B.; Jung, S.; Park, S.; Kim, E.; Oh, S. Effect of Lactobacillus acidophilus NS1 on plasma cholesterol levels in diet-induced obese mice. J. Dairy Sci. 2015, 98, 1492–1501. [Google Scholar] [CrossRef]
- Park, S.S.; Lee, Y.J.; Song, S.; Kim, B.; Kang, H.; Oh, S.; Kim, E. Lactobacillus acidophilus NS1 attenuates diet-induced obesity and fatty liver. J. Endocrinol. 2018, 237, 87–100. [Google Scholar] [CrossRef]
- Janda, A.; Bowen, A.; Greenspan, N.S.; Casadevall, A. Ig Constant Region Effects on Variable Region Structure and Function. Front. Microbiol. 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.A. Canine IgA and IgA deficiency: Implications for immunization against respiratory pathogens. Can. Vet. J. 2019, 60, 1305–1311. [Google Scholar] [PubMed]
- Maurer, M.A.; Meyer, L.; Bianchi, M.; Turner, H.L.; Le, N.P.L.; Steck, M.; Wyrzucki, A.; Orlowski, V.; Ward, A.B.; Crispin, M.; et al. Glycosylation of Human IgA Directly Inhibits Influenza A and Other Sialic-Acid-Binding Viruses. Cell Rep. 2018, 23, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Herr, A.B.; Ballister, E.R.; Bjorkman, P.J. Insights into IgA-mediated immune responses from the crystal structures of human FcalphaRI and its complex with IgA1-Fc. Nature 2003, 423, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Breedveld, A.; van Egmond, M. IgA and FcαRI: Pathological Roles and Therapeutic Opportunities. Front. Immunol. 2019, 10, 553. [Google Scholar] [CrossRef]
- De Sousa-Pereira, P.; Woof, J.M. IgA: Structure, Function, and Developability. Antibodies 2019, 8, 57. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Kwaa, A.K.R.; Talana, C.A.G.; Blankson, J.N. Interferon Alpha Enhances NK Cell Function and the Suppressive Capacity of HIV-Specific CD8+ T Cells. J. Virol. 2019, 93, e01541-18. [Google Scholar] [CrossRef]
- Malek, T.R. The main function of IL-2 is to promote the development of T regulatory cells. J. Leukoc. Biol. 2003, 74, 961–965. [Google Scholar] [CrossRef]
- Chen, L.; Liu, W.; Li, Y.; Luo, S.; Liu, Q.; Zhong, Y.; Jian, Z.; Bao, M. Lactobacillus acidophilus ATCC 4356 attenuates the atherosclerotic progression through modulation of oxidative stress and inflammatory process. Int. Immunopharmacol. 2013, 17, 108–115. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Natella, F.; Scaccini, C. Total antioxidant capacity as a tool to assess redox status: Critical view and experimental data. Free Radic. Biol. Med. 2000, 29, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Handl, S.; Dowd, S.E.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol. Ecol. 2011, 76, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.S. Companion animals symposium: Microbes and gastrointestinal health of dogs and cats. J. Anim. Sci. 2011, 89, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, A.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items | Content % |
|---|---|
| Ingredients | |
| Corn | 19.00 |
| Soybean meal | 8.00 |
| Beef powder | 4.00 |
| Sweet potato granules | 4.00 |
| Broken rice | 13.00 |
| Beet pulp | 8.00 |
| Broken wall hemoglobulin powder | 2.50 |
| Fish meal | 1.00 |
| Bone meal | 5.00 |
| Chicken powder | 10.00 |
| Saccharomyces cerevisiae | 5.00 |
| Peameal | 3.00 |
| Chicken oil | 11.00 |
| Taste enhancer | 1.70 |
| Methionine | 0.30 |
| Chicken liver powder | 2.50 |
| Premix 1 | 2.00 |
| Total | 100.00 |
| Nutrient levels 2 | |
| CP | 28.88 |
| EE | 8.85 |
| Ash | 6.24 |
| Ca | 0.89 |
| P | 0.66 |
| Items | CON | LAC | p-Value |
|---|---|---|---|
| Body weight on day 7/kg | 4.97 ± 0.52 | 5.23 ± 0.27 | 0.668 |
| Body weight on day 14/kg | 5.03 ± 0.53 | 5.38 ± 0.27 | 0.580 |
| Body weight on day 21/kg | 4.89 ± 0.61 | 5.22 ± 0.41 | 0.668 |
| Body weight on day 28/kg | 5.46 ± 0.57 | 5.73 ± 0.34 | 0.694 |
| ADG (g/d) | 30.95 ± 7.64 | 34.23 ± 6.85 | 0.756 |
| WGR | 0.20 ± 0.06 | 0.20 ± 0.04 | 0.984 |
| TWG (kg) | 0.87 ± 0.21 | 0.96 ± 0.19 | 0.756 |
| Items | CON | LAC | p-Value |
|---|---|---|---|
| DM | 77.69 ± 0.35 | 78.60 ± 1.42 | 0.559 |
| CP | 72.46 ± 0.96 | 72.24 ± 2.22 | 0.930 |
| EE | 95.95 ± 0.33 | 95.97 ± 0.30 | 0.965 |
| Carbohydrates | 83.31 ± 0.39 | 84.21 ± 1.17 | 0.491 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, A.; Chen, X.; Zhao, W.; Nan, W.; Huang, Y.; Sun, Y.; Zhang, H.; Xu, C. Bacterial Community Influences the Effects of Lactobacillus acidophilus on Lipid Metabolism, Immune Response, and Antioxidant Capacity in Dogs. Animals 2024, 14, 1257. https://doi.org/10.3390/ani14091257
Mao A, Chen X, Zhao W, Nan W, Huang Y, Sun Y, Zhang H, Xu C. Bacterial Community Influences the Effects of Lactobacillus acidophilus on Lipid Metabolism, Immune Response, and Antioxidant Capacity in Dogs. Animals. 2024; 14(9):1257. https://doi.org/10.3390/ani14091257
Chicago/Turabian StyleMao, Aipeng, Xiaoli Chen, Weigang Zhao, Weixiao Nan, Yao Huang, Yalong Sun, Haihua Zhang, and Chao Xu. 2024. "Bacterial Community Influences the Effects of Lactobacillus acidophilus on Lipid Metabolism, Immune Response, and Antioxidant Capacity in Dogs" Animals 14, no. 9: 1257. https://doi.org/10.3390/ani14091257