

Effect of Dietary Eicosapentaenoic and Docosahexaenoic Fatty Acid Supplementation during the Last Month of Gestation on Fatty Acid Metabolism and Oxidative Status in Charolais Cows and Calves
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Fatty Acids Analysis
2.3. Determination of Thiobarbituric Acid-Reactive Substances (TBARS)
2.4. Determination of Total Phenols in Milk
2.5. Determination of Cholesterol in Milk
2.6. Determination of FASN and ACACA Gene Expression in Whole Blood
ΔΔCt = ΔCT (a target sample) − ΔCT (a reference sample)
R = 2ΔΔCt
2.7. Statistical Analysis
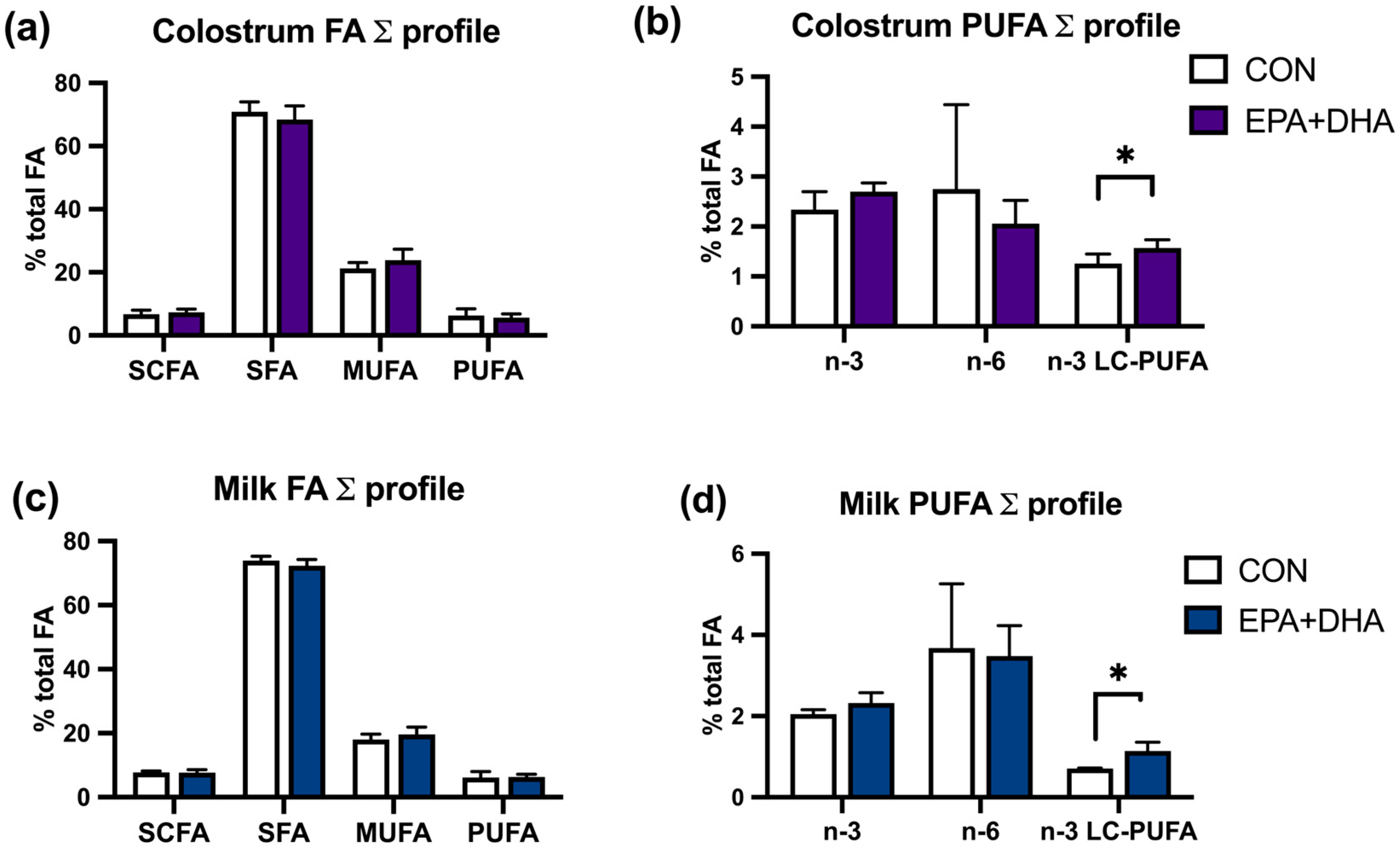
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koletzko, B.; Larque, E.; Demmelmair, H. Placental transfer of long-chain polyunsaturated fatty acids (LC-PUFA). J. Perinat. Med. 2007, 35 (Suppl. S1), S5–S11. [Google Scholar] [CrossRef]
- Gnott, M.; Vogel, L.; Kröger-Koch, C.; Dannenberger, D.; Tuchscherer, A.; Tröscher, A.; Trevisi, E.; Stefaniak, T.; Bajzert, J.; Starke, A.; et al. Changes in fatty acids in plasma and association with the inflammatory response in dairy cows abomasally infused with essential fatty acids and conjugated linoleic acid during late and early lactation. J. Dairy Sci. 2020, 103, 11889–11910. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Greco, L.; Favoreto, M.; Marsola, R.; Martins, L.; Bisinotto, R.; Shin, J.; Lock, A.; Block, E.; Thatcher, W. Effect of supplementing fat to pregnant nonlactating cows on colostral fatty acid profile and passive immunity of the newborn calf. J. Dairy Sci. 2014, 97, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef] [PubMed]
- Campbell, F.M.; Gordon, M.J.; Dutta-Roy, A.K. Plasma membrane fatty acid-binding protein (FABPpm) of the sheep placenta. Biochim. Biophys. Acta 1994, 1214, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Roque-Jiménez, J.A.; Rosa-Velázquez, M.; Pinos-Rodríguez, J.M.; Vicente-Martínez, J.G.; Mendoza-Cervantes, G.; Flores-Primo, A.; Lee-Rangel, H.A.; Relling, A.E. Role of Long Chain Fatty Acids in Developmental Programming in Ruminants. Animals 2021, 11, 762. [Google Scholar] [CrossRef]
- Desantadina, R.; Quintana, S.; Recavarren, M.I.; Relling, A.E. Effect of time of gestation on fatty acid transporters mRNA expression in bovine placenta. Biosci. J. 2017, 34, 180–185. [Google Scholar] [CrossRef]
- Hanebutt, F.L.; Demmelmair, H.; Schiessl, B.; Larqué, E.; Koletzko, B. Long-chain polyunsaturated fatty acid (LC-PUFA) transfer across the placenta. Clin. Nutr. 2008, 27, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.K.; Weber, W.J.; Moore, C.E.; Bauman, D.E.; Hansen, L.B.; Chester-Jones, H.; Crooker, B.A.; Baumgard, L.H. Effects of week of lactation and genetic selection for milk yield on milk fatty acid composition in Holstein cows. J. Dairy Sci. 2005, 88, 3886–3893. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Masson, L.L.; Lock, A.L.; Mottram, T.T. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cows. J. Dairy Sci. 2006, 89, 1604–1612. [Google Scholar] [CrossRef]
- Bernard, L.; Leroux, C.; Chilliard, Y. Expression and nutritional regulation of lipogenic genes in the ruminant lactating mammary gland. Adv. Exp. Med. Biol. 2008, 606, 67–108. [Google Scholar] [CrossRef] [PubMed]
- Stoop, W.; Bovenhuis, H.; Heck, J.; Van Arendonk, J. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L. Milk Fat: Origin of Fatty Acids and Influence of Nutritional Factors Thereon. In Advanced Dairy Chemistry Volume 2 Lipids; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: Boston, MA, USA, 2006; pp. 43–92. [Google Scholar]
- Lenihan-Geels, G.; Bishop, K.S.; Ferguson, L.R. Alternative sources of omega-3 fats: Can we find a sustainable substitute for fish? Nutrients 2013, 5, 1301–1315. [Google Scholar] [CrossRef]
- Lock, A.L.; Bauman, D.E. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Castaneda-Gutierrez, E.; de Veth, M.J.; Lock, A.L.; Dwyer, D.A.; Murphy, K.D.; Bauman, D.E. Effect of supplementation with calcium salts of fish oil on n-3 fatty acids in milk fat. J. Dairy Sci. 2007, 90, 4149–4156. [Google Scholar] [CrossRef] [PubMed]
- Boeckaert, C.; Vlaeminck, B.; Dijkstra, J.; Issa-Zacharia, A.; Van Nespen, T.; Van Straalen, W.; Fievez, V. Effect of dietary starch or micro algae supplementation on rumen fermentation and milk fatty acid composition of dairy cows. J. Dairy Sci. 2008, 91, 4714–4727. [Google Scholar] [CrossRef] [PubMed]
- Abughazaleh, A.A.; Potu, R.B.; Ibrahim, S. Short communication: The effect of substituting fish oil in dairy cow diets with docosahexaenoic acid-micro algae on milk composition and fatty acids profile. J. Dairy Sci. 2009, 92, 6156–6159. [Google Scholar] [CrossRef]
- Glover, K.E.; Budge, S.; Rose, M.; Rupasinghe, H.P.; Maclaren, L.; Green-Johnson, J.; Fredeen, A.H. Effect of feeding fresh forage and marine algae on the fatty acid composition and oxidation of milk and butter. J. Dairy Sci. 2012, 95, 2797–2809. [Google Scholar] [CrossRef] [PubMed]
- Brookes, K.J.; Chen, W.; Xu, X.; Taylor, E.; Asherson, P. Association of fatty acid desaturase genes with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2006, 60, 1053–1061. [Google Scholar] [CrossRef]
- Amminger, G.P.; Berger, G.E.; Schafer, M.R.; Klier, C.; Friedrich, M.H.; Feucht, M. Omega-3 fatty acids supplementation in children with autism: A double-blind randomized, placebo-controlled pilot study. Biol. Psychiatry 2007, 61, 551–553. [Google Scholar] [CrossRef]
- Bauman, D.E.; Mather, I.H.; Wall, R.J.; Lock, A.L. Major advances associated with the biosynthesis of milk. J. Dairy Sci. 2006, 89, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Mašek, T.; Krstulović, L.; Brozić, D.; Vranić, M.; Maurić, M.; Bajić, M.; Starčević, K. Cow colostrum and early milk enriched with eicosapentaenoic and docosahexaenoic fatty acid. Eur. Food Res. Technol. 2014, 238, 635–640. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Doreau, M. Effect of different types of forages, animal fat or marine oils in cow’s diet on milk fat secretion and composition, especially conjugated linoleic acid (CLA) and polyunsaturated fatty acids. Livest. Prod. Sci. 2001, 70, 31–48. [Google Scholar] [CrossRef]
- Mach Casellas, N.; Jacobs, A.A.A.; Kruijt, L.; Van Baal, J.; Smits, M.A. Alteration of gene expression in mammary gland tissue of dairy cows in response to dietary unsaturated fatty acids. Animal 2011, 5, 1217–1230. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W.; Latimer, G., Jr. AOAC International Official Methods of Analysis; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Grotto, D.; Santa Maria, L.; Boeira, S.; Valentini, J.; Charão, M.; Moro, A.; Nascimento, P.; Pomblum, V.; Garcia, S. Rapid quantification of malondialdehyde in plasma by high performance liquid chromatography–visible detection. J. Pharm. Biomed. Anal. 2007, 43, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Ough, C.S.; Amerine, M.A. Methods for Analysis of Musts and Wines; John Wiley & Sons: Hoboken, NJ, USA, 1988. [Google Scholar]
- Sreeramulu, D.; Raghunath, M. Antioxidant activity and phenolic content of roots, tubers and vegetables commonly consumed in India. Food Res. Int. 2010, 43, 1017–1020. [Google Scholar] [CrossRef]
- Fletouris, D.; Botsoglou, N.; Psomas, I.; Mantis, A. Rapid determination of cholesterol in milk and milk products by direct saponification and capillary gas chromatography. J. Dairy Sci. 1998, 81, 2833–2840. [Google Scholar] [CrossRef]
- Robinson, T.; Sutherland, I.; Sutherland, J. Validation of candidate bovine reference genes for use with real-time PCR. Vet. Immunol. Immunopathol. 2007, 115, 160–165. [Google Scholar] [CrossRef]
- Alessandri, J.-M.; Extier, A.; Al-Gubory, K.H.; Langelier, B.; Baudry, C.; LePoupon, C.; Lavialle, M.; Guesnet, P. Ovariectomy and 17β-estradiol alter transcription of lipid metabolism genes and proportions of neo-formed n-3 and n-6 long-chain polyunsaturated fatty acids differently in brain and liver. J. Nutr. Biochem. 2011, 22, 820–827. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vahmani, P.; Glover, K.; Fredeen, A. Effects of pasture versus confinement and marine oil supplementation on the expression of genes involved in lipid metabolism in mammary, liver, and adipose tissues of lactating dairy cows. J. Dairy Sci. 2014, 97, 4174–4183. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Inada, S.; Kobayashi, E.; Abe, T.; Hasebe, H.; Sasazaki, S.; Oyama, K.; Mannen, H. Identification of SNPs in the FASN gene and their effect on fatty acid milk composition in Holstein cattle. Livest. Sci. 2012, 144, 281–284. [Google Scholar] [CrossRef]
- Doreau, M.; Chilliard, Y. Effects of ruminal or postruminal fish oil supplementation on intake and digestion in dairy cows. Reprod. Nutr. Dev. 1997, 37, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Scollan, N.D.; Dhanoa, M.S.; Choi, N.J.; Maeng, W.J.; Enser, M.; Wood, J.D. Biohydrogenation and digestion of long chain fatty acids in steers fed on different sources of lipid. J. Agric. Sci. 2001, 136, 345–355. [Google Scholar] [CrossRef]
- Shingfield, K.; Ahvenjarvi, S.; Toivonen, V.; Arola, A.; Nurmela, K.; Huhtanen, P.; Griinari, J.M. Effect of dietary fish oil on biohydrogenation of fatty acids and milk fatty acid content in cows. Anim. Sci. 2003, 77, 165–180. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Mansbridge, R.M.; Doreau, M. Ruminant milk fat plasticity: Nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 2000, 49, 181–205. [Google Scholar] [CrossRef]
- Gulati, S.K.; Ashes, J.R.; Scott, T.W. Hydrogenation of eicosapentaenoic and docosahexaenoic acids and their incorporation into milk fat. Anim. Feed. Sci. Technol. 1999, 79, 57–64. [Google Scholar] [CrossRef]
- Wright, T.C.; Holub, B.J.; McBride, B.W. Apparent transfer efficiency of docosahexaenoic acid from diet to milk in dairy cows. Canadian J. Anim. Sci. 1999, 79, 565–568. [Google Scholar] [CrossRef]
- Harvatine, K.J.; Boisclair, Y.R.; Bauman, D.E. Recent advances in the regulation of milk fat synthesis. Animal 2009, 3, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Shingfield, K.J.; Bernard, L.; Leroux, C.; Chilliard, Y. Role of trans fatty acids in the nutritional regulation of mammary lipogenesis in ruminants. Animal 2010, 4, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Harvatine, K.J.; Bauman, D.E. SREBP1 and thyroid hormone responsive spot 14 (S14) are involved in the regulation of bovine mammary lipid synthesis during diet-induced milk fat depression and treatment with CLA. J. Nutr. 2006, 136, 2468–2474. [Google Scholar] [CrossRef] [PubMed]
- Capper, J.L.; Wilkinson, R.G.; Mackenzie, A.M.; Sinclair, L.A. The effect of fish oil supplementation of pregnant and lactating ewes on milk production and lamb performance. Animal 2007, 1, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M.; Urrutia, N.L.; Vásconez-Montúfar, C.; Morales, R. Factors associated with the content of mammary-synthesized fatty acids in milk fat: A meta-analysis. J. Dairy Sci. 2019, 102, 4105–4117. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.W.; Schroeder, J.W.; Park, C.S.; Keller, W.L.; Schimek, D.E. Altering the Fatty Acids in Milk Fat by Including Canola Seed in Dairy Cattle Diets. J. Dairy Sci. 2005, 88, 3084–3094. [Google Scholar] [CrossRef] [PubMed]
- Ballou, M.; DePeters, E. Supplementing milk replacer with omega-3 fatty acids from fish oil on immunocompetence and health of Jersey calves. J. Dairy Sci. 2008, 91, 3488–3500. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B. Fats for brains. Eur. J. Clin. Nutr. 1992, 46 (Suppl. S1), S51–S62. [Google Scholar] [PubMed]
- Pickard, R.M.; Beard, A.P.; Seal, C.J.; Edwards, S.A. Lamb Vigour is Affected by DHA Supplementation of Ewe Diets during Late Pregnancy. Proceedings EAAP, 312. Available online: https://docplayer.net/20716119-Lamb-vigour-is-affected-by-dha-supplementation-of-ewe-diets-during-late-pregnancy.html (accessed on 18 April 2024).
- Capper, J.; Wilkinson, R.; Sinclair, L.; Pattinson, S.; Mackenzie, A. The effect of long-chain polyunsaturated fatty acid and vitamin E supplementation of ewes on neonatal lamb vigour, lamb growth and colostrum parameters. In Proceedings of the British Society of Animal Science; Cambridge University Press: Cambridge, UK, 2002; p. 7. [Google Scholar]
- Burns, P.; Bonnette, T.; Engle, T.; Whittier, J. Effects of fishmeal supplementation on fertility and plasma Ω-3 fatty acid profiles in primiparous, lactating beef cows. Prof. Anim. Sci. 2002, 18, 373–379. [Google Scholar] [CrossRef]
- Childs, S.; Hennessy, A.; Sreenan, J.; Wathes, D.; Cheng, Z.; Stanton, C.; Diskin, M.; Kenny, D. Effect of level of dietary n-3 polyunsaturated fatty acid supplementation on systemic and tissue fatty acid concentrations and on selected reproductive variables in cattle. Theriogenology 2008, 70, 595–611. [Google Scholar] [CrossRef]
- Hill, T.; VandeHaar, M.; Sordillo, L.; Catherman, D.; Bateman Ii, H.; Schlotterbeck, R. Fatty acid intake alters growth and immunity in milk-fed calves. J. Dairy Sci. 2011, 94, 3936–3948. [Google Scholar] [CrossRef]
- Moallem, U.; Zachut, M. The effects of supplementation of various n-3 fatty acids to late-pregnant dairy cows on plasma fatty acid composition of the newborn calves. J. Dairy Sci. 2012, 95, 4055–4058. [Google Scholar] [CrossRef]
- Marques, R.S.; Cooke, R.F.; Rodrigues, M.C.; Brandão, A.P.; Schubach, K.M.; Lippolis, K.D.; Moriel, P.; Perry, G.A.; Lock, A.; Bohnert, D.W. Effects of supplementing calcium salts of polyunsaturated fatty acids to late-gestating beef cows on performance and physiological responses of the offspring. J. Anim. Sci. 2017, 95, 5347–5357. [Google Scholar] [CrossRef]
- Gibson, R.A.; Muhlhausler, B.; Makrides, M. Conversion of linoleic acid and alpha-linolenic acid to long-chain polyunsaturated fatty acids (LCPUFAs), with a focus on pregnancy, lactation and the first 2 years of life. Matern. Child Nutr. 2011, 7, 17–26. [Google Scholar] [CrossRef]
- Basak, S.; Mallick, R.; Duttaroy, A.K. Maternal Docosahexaenoic Acid Status during Pregnancy and Its Impact on Infant Neurodevelopment. Nutrients 2020, 12, 3615. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.N.; Rivera-Acevedo, K.C.; Relling, A.E. Prepartum fatty acid supplementation in sheep I. Eicosapentaenoic and docosahexaenoic acid supplementation do not modify ewe and lamb metabolic status and performance through weaning. J. Anim. Sci. 2018, 96, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.N.; Murphy, K.D.; Relling, A.E. Prepartum fatty acid supplementation in sheep. II. Supplementation of eicosapentaenoic acid and docosahexaenoic acid during late gestation alters the fatty acid profile of plasma, colostrum, milk and adipose tissue, and increases lipogenic gene expression of adipose tissue. J. Anim. Sci. 2018, 96, 1181–1204. [Google Scholar] [PubMed]
- Uken, K.L.; Schäff, C.T.; Vogel, L.; Gnott, M.; Dannenberger, D.; Görs, S.; Tuchscherer, A.; Tröscher, A.; Liermann, W.; Hammon, H.M. Modulation of colostrum composition and fatty acid status in neonatal calves by maternal supplementation with essential fatty acids and conjugated linoleic acid starting in late lactation. J. Dairy Sci. 2021, 104, 4950–4969. [Google Scholar] [CrossRef]
- Vogel, L.; Gnott, M.; Kröger-Koch, C.; Dannenberger, D.; Tuchscherer, A.; Tröscher, A.; Kienberger, H.; Rychlik, M.; Starke, A.; Bachmann, L.; et al. Effects of abomasal infusion of essential fatty acids together with conjugated linoleic acid in late and early lactation on performance, milk and body composition, and plasma metabolites in dairy cows. J. Dairy Sci. 2020, 103, 7431–7450. [Google Scholar] [CrossRef]
- Zeng, X.; Li, S.; Liu, L.; Cai, S.; Ye, Q.; Xue, B.; Wang, X.; Zhang, S.; Chen, F.; Cai, C. Role of functional fatty acids in modulation of reproductive potential in livestock. J. Anim. Sci. Biotechnol. 2023, 14, 24. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 532. [Google Scholar] [CrossRef] [PubMed]
- Wullepit, N.; Hostens, M.; Ginneberge, C.; Fievez, V.; Opsomer, G.; Fremaut, D.; De Smet, S. Influence of a marine algae supplementation on the oxidative status of plasma in dairy cows during the periparturient period. Prev. Vet. Med. 2012, 103, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Scislowski, V.; Bauchart, D.; Gruffat, D.; Laplaud, P.-M.; Durand, D. Effect of dietary n − 6 and n − 3 polyunsaturated fatty acids on peroxidizability of lipoproteins in steers. Lipids 2005, 40, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Lopreiato, V.; Mezzetti, M.; Cattaneo, L.; Ferronato, G.; Minuti, A.; Trevisi, E. Role of nutraceuticals during the transition period of dairy cows: A review. J. Anim. Sci. Biotechnol. 2020, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Pi, Y.; Gao, S.; Ma, L.; Zhu, Y.; Wang, J.; Zhang, J.; Xu, J.; Bu, D. Effectiveness of rubber seed oil and flaxseed oil to enhance the α-linolenic acid content in milk from dairy cows. J. Dairy Sci. 2016, 99, 5719–5730. [Google Scholar] [CrossRef] [PubMed]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed. Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Opgenorth, J.; Sordillo, L.M.; VandeHaar, M.J. Colostrum supplementation with n-3 fatty acids and α-tocopherol alters plasma polyunsaturated fatty acid profile and decreases an indicator of oxidative stress in newborn calves. J. Dairy Sci. 2020, 103, 3545–3553. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Martini, S. Increasing omega fatty acid content in cow’s milk through diet manipulation: Effect on milk flavor. J. Dairy Sci. 2009, 92, 1378–1386. [Google Scholar] [CrossRef]
- Puppel, K.; Nałęcz-Tarwacka, T.; Kuczyńska, B.; Gołębiewski, M.; Kordyasz, M. Effect of different fat supplements on the antioxidant capacity of cow’s milk. Arch. Anim. Breed. 2013, 56, 178–190. [Google Scholar] [CrossRef]
- Baer, R.; Ryali, J.; Schingoethe, D.; Kasperson, K.; Donovan, D.; Hippen, A.; Franklin, S. Composition and properties of milk and butter from cows fed fish oil. J. Dairy Sci. 2001, 84, 345–353. [Google Scholar] [CrossRef]
- Ahnadi, C.E.; Beswick, N.; Delbecchi, L.; Kennelly, J.J.; Lacasse, P. Addition of fish oil to diets for dairy cows. II. Effects on milk fat and gene expression of mammary lipogenic enzymes. J. Dairy Res. 2002, 69, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Piperova, L.S.; Teter, B.B.; Bruckental, I.; Sampugna, J.; Mills, S.E.; Yurawecz, M.P.; Fritsche, J.; Ku, K.; Erdman, R.A. Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat–depressing diet. J. Nutr. 2000, 130, 2568–2574. [Google Scholar] [CrossRef]
- Peterson, D.G.; Matitashvili, E.A.; Bauman, D.E. Diet-induced milk fat depression in dairy cows results in increased trans-10, cis-12 CLA in milk fat and coordinate suppression of mRNA abundance for mammary enzymes involved in milk fat synthesis. J. Nutr. 2003, 133, 3098–3102. [Google Scholar] [CrossRef]
- Loor, J.; Piperova, L.; Everts, R.; Rodriguez-Zas, S.; Drackley, J.; Erdman, R.; Lewin, H. Mammary gene expression profiling in cows fed a milk fat-depressing diet using a bovine 13,000 oligonucleotide microarray. J. Dairy Sci. 2005, 88, 120. [Google Scholar]
- Donovan, D.C.; Schingoethe, D.J.; Baer, R.J.; Ryali, J.; Hippen, A.R.; Franklin, S.T. Influence of dietary fish oil on conjugated linoleic acid and other fatty acids in milk fat from lactating dairy cows. J. Dairy Sci. 2000, 83, 2620–2628. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.; Christie, W. Lipid metabolism in the mammary gland of ruminant animals. In Lipid Metabolism in Ruminant Animals; Elsevier: Amsterdam, The Netherlands, 1981; pp. 227–277. [Google Scholar]
- Barber, M.C.; Clegg, R.A.; Travers, M.T.; Vernon, R.G. Lipid metabolism in the lactating mammary gland. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1997, 1347, 101–126. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Corl, B.A.; Dwyer, D.A.; Sæbø, A.; Bauman, D.E. Identification of the conjugated linoleic acid isomer that inhibits milk fat synthesis. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2000, 278, R179–R184. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.; Matitashvili, E.; Corl, B.; Dwyer, D.; Bauman, D. Trans-10, cis-12 conjugated linoleic acid decreases lipogenic rates and expression of genes involved in milk lipid synthesis in dairy cows. J. Dairy Sci. 2002, 85, 2155–2163. [Google Scholar] [CrossRef]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Peterson, D.G.; Matitashvili, E.A.; Bauman, D.E. The inhibitory effect of trans-10, cis-12 CLA on lipid synthesis in bovine mammary epithelial cells involves reduced proteolytic activation of the transcription factor SREBP-1. J. Nutr. 2004, 134, 2523–2527. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, P.S.; De Mendoza, D. Control of fatty acid desaturation: A mechanism conserved from bacteria to humans. Mol. Microbiol. 2006, 62, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Duplus, E.; Forest, C. Is there a single mechanism for fatty acid regulation of gene transcription? Biochem. Pharmacol. 2002, 64, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Kadegowda, A.K.; Bionaz, M.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Peroxisome proliferator-activated receptor-gamma activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Perfield, J.W.; Harvatine, K.J.; Baumgard, L.H. Regulation of fat synthesis by conjugated linoleic acid: Lactation and the ruminant model. J. Nutr. 2008, 138, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Loor, J.J.; Herbein, J.H. Trans 10, c is 12–18: 2 Is a More Potent Inhibitor of De Novo Fatty Acid Synthesis and Desaturation than cis 9, trans 11–18: 2 in the Mammary Gland of Lactating Mice. J. Nutr. 2004, 134, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Harvatine, K.J.; Boisclair, Y.R.; Bauman, D.E. Supplementation with fish oil as a source of n–3 fatty acids does not downregulate mammary lipogenesis in lactating mice. J. Nutr. 2013, 143, 1913–1919. [Google Scholar]
- Murrieta, C.; Hess, B.; Scholljegerdes, E.; Engle, T.; Hossner, K.; Moss, G.; Rule, D. Evaluation of milk somatic cells as a source of mRNA for study of lipogenesis in the mammary gland of lactating beef cows supplemented with dietary high-linoleate safflower seeds. J. Anim. Sci. 2006, 84, 2399–2405. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % on As-Fed Basis | Alfalfa Haylage | Corn, Ground | Fatmix-65 | |
|---|---|---|---|---|
| DM | 51.2 | 88 | 95 | |
| Ash | 4.1 | 1.3 | 30 | |
| Ether extract (EE) | 1.5 | 3.7 | 65 | |
| Crude protein (CP) | 9.4 | 8.1 | / | |
| Crude fiber (CF) | 15.4 | 2.7 | / | |
| Neutral detergent fiber (NDF) | 22.2 | 12.8 | / | |
| Acid detergent fiber (ADF) | 7.1 | 3.5 | / | |
| NSC 1 | 20.8 | 72.2 | / | |
| EPA (g/100 g) | 0.00 | 0.00 | 9.1 | |
| DHA (g/100 g) | 0.00 | 0.00 | 7.8 | |
| estimated daily intake/cow | CON | ad libitum | 1 kg | / |
| EPA + DHA | ad libitum | 1 kg | 100 g | |
| % on DM Basis | CON | EPA + DHA |
|---|---|---|
| Palmitic acid (16:00) | 0.709 | 0.873 |
| Palmitoleic acid (16:1) | 0.011 | 0.040 |
| Stearic acid (18:00) | 0.088 | 0.109 |
| Oleic acid (18:1) | 0.169 | 0.299 |
| Linoleic acid (18:2n6) | 0.737 | 0.773 |
| Linolenic acid (18:3n3) | 1.139 | 1.140 |
| EPA (20:5n-3) | 0.000 | 0.083 |
| DHA (22:6n3) | 0.000 | 0.071 |
| Gene | Primers | Reference |
|---|---|---|
| ACACA | F: (5′–CAT CTT GTC CGA AAC GTC GAT–3′) | [35] |
| R: (5′–CCC TTC GAA CAT ACA CCT CCA–3′) | ||
| FASN | F: (5′–CTA CCA AGC CAG GCA GGT C–3′) | [36] |
| R: (5′–GCC ATT GTA CTT GGG CTT GT–3′) | ||
| GADPH | F: (5′–CGT GTC TGT TGT GGA TCT GAC CTG–3′) | [36] |
| R: (5′–CAA CCT GGT CCT CAG TGT AGC CT–3′) |
| FA (% of Total FA) | Colostrum | 7th Day in Milk | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | EPA + DHA | p-Value | CON | EPA + DHA | p-Value | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| C4:0 | 4.16 | 1.50 | 3.94 | 1.00 | 0.778 | 1.66 | 0.28 | 1.83 | 0.29 | 0.437 |
| C6:0 | 0.84 | 0.27 | 1.21 | 0.23 | 0.036 | 1.83 | 0.11 | 1.89 | 0.19 | 0.614 |
| C8:0 | 0.50 | 0.11 | 0.70 | 0.15 | 0.043 | 1.29 | 0.09 | 1.30 | 0.18 | 0.929 |
| C10:0 | 1.21 | 0.19 | 1.48 | 0.26 | 0.094 | 2.95 | 0.31 | 2.68 | 0.34 | 0.309 |
| C11:0 | 0.04 | 0.02 | 0.03 | 0.04 | 0.679 | 0.05 | 0.01 | 0.04 | 0.01 | 0.267 |
| C12:0 | 2.44 | 0.18 | 2.33 | 0.30 | 0.495 | 3.57 | 0.38 | 2.86 | 0.37 | 0.056 |
| C14:0 | 13.22 | 1.12 | 11.44 | 1.22 | 0.036 | 1.46 | 0.27 | 10.79 | 0.68 | 0.104 |
| C14:1 | 0.99 | 0.50 | 0.96 | 0.89 | 0.957 | 0.71 | 0.11 | 0.52 | 0.13 | 0.075 |
| C15:0 | 1.33 | 0.28 | 1.30 | 0.22 | 0.849 | 1.74 | 0.50 | 1.80 | 0.16 | 0.860 |
| C15:1 | 0.02 | 0.02 | 0.02 | 0.02 | 0.824 | 0.05 | 0.06 | 0.04 | 0.03 | 0.881 |
| C16:0 | 38.98 | 2.80 | 34.57 | 3.78 | 0.066 | 37.47 | 1.44 | 33.97 | 2.65 | 0.052 |
| C16:1 | 4.17 | 2.54 | 3.46 | 2.42 | 0.648 | 2.09 | 0.42 | 1.71 | 0.26 | 0.251 |
| C17:0 | 0.77 | 0.17 | 0.90 | 0.23 | 0.347 | 0.76 | 0.09 | 0.84 | 0.12 | 0.304 |
| C17:1 | 0.28 | 0.14 | 0.27 | 0.07 | 0.977 | 0.19 | 0.04 | 0.31 | 0.36 | 0.513 |
| C18:0 | 7.20 | 1.80 | 10.21 | 2.24 | 0.043 | 10.86 | 1.30 | 13.71 | 1.62 | 0.040 |
| C18:1n9 | 15.63 | 2.66 | 19.08 | 0.74 | 0.023 | 14.74 | 1.24 | 16.76 | 2.32 | 0.159 |
| C18:2n6 | 2.29 | 1.71 | 1.58 | 0.40 | 0.364 | 3.40 | 1.55 | 3.15 | 0.77 | 0.812 |
| C18:3n6 | 0.09 | 0.07 | 0.05 | 0.06 | 0.407 | 0.08 | 0.06 | 0.12 | 0.11 | 0.613 |
| C18:3n3 | 0.92 | 0.18 | 0.90 | 0.14 | 0.876 | 0.86 | 0.06 | 0.79 | 0.10 | 0.268 |
| C18:4n3 | 0.12 | 0.16 | 0.18 | 0.15 | 0.579 | 0.36 | 0.08 | 0.29 | 0.16 | 0.416 |
| C20:0 | 0.16 | 0.04 | 0.21 | 0.08 | 0.262 | 0.26 | 0.04 | 0.43 | 0.29 | 0.257 |
| C20:1 | 0.04 | 0.01 | 0.05 | 0.01 | 0.240 | 0.10 | 0.09 | 0.07 | 0.06 | 0.605 |
| C20:2 | 0.11 | 0.11 | 0.13 | 0.24 | 0.877 | 0.16 | 0.25 | 0.05 | 0.05 | 0.497 |
| C20:4n6 | 0.37 | 0.04 | 0.42 | 0.10 | 0.296 | 0.20 | 0.01 | 0.21 | 0.03 | 0.396 |
| C20:3n3 | 0.04 | 0.03 | 0.05 | 0.03 | 0.820 | 0.11 | 0.13 | 0.10 | 0.09 | 0.919 |
| C20:4n3 | 0.22 | 0.06 | 0.19 | 0.08 | 0.643 | 0.09 | 0.02 | 0.09 | 0.03 | 0.955 |
| C20:5n3 | 0.31 | 0.03 | 0.40 | 0.04 | 0.003 | 0.40 | 0.04 | 0.76 | 0.19 | 0.012 |
| C22:0 | 0.04 | 0.03 | 0.05 | 0.04 | 0.532 | 0.07 | 0.01 | 0.06 | 0.01 | 0.475 |
| C22:1 | 0.02 | 0.03 | 0.01 | 0.01 | 0.447 | 0.03 | 0.01 | 0.03 | 0.03 | 0.757 |
| C22:2 | 1.25 | 1.30 | 0.84 | 0.85 | 0.543 | 0.40 | 0.21 | 0.53 | 0.23 | 0.457 |
| C22:5n3 | 0.61 | 0.11 | 0.83 | 0.08 | 0.005 | 0.18 | 0.01 | 0.22 | 0.03 | 0.038 |
| C24:0 | 0.02 | 0.03 | 0.04 | 0.03 | 0.366 | 0.04 | 0.03 | 0.12 | 0.14 | 0.238 |
| C22:6n3 | 0.12 | 0.01 | 0.15 | 0.01 | 0.001 | 0.04 | 0.04 | 0.07 | 0.06 | 0.428 |
| C24:1 | 0.05 | 0.02 | 0.04 | 0.04 | 0.577 | 0.08 | 0.06 | 0.18 | 0.10 | 0.128 |
| % | CON | EPA + DHA | p-Value | |||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||
| Colostrum | Milk fat | 4.18 | 1.38 | 3.94 | 2.71 | 0.40 |
| Protein | 18.10 | 3.62 | 16.54 | 1.91 | 0.78 | |
| Lactose | 2.20 | 0.14 | 2.70 | 0.19 | 0.05 | |
| Dry matter | 26.03 | 2.42 | 24.67 | 4.17 | 0.35 | |
| Fat free dry matter | 21.27 | 3.55 | 20.24 | 1.91 | 0.69 | |
| Cholesterol (μg/100 mL) | 67.46 | 32.03 | 41.61 | 18.39 | 0.08 | |
| 7th-day in milk | Milk fat | 3.22 | 0.37 | 3.16 | 1.31 | 0.92 |
| Protein | 3.49 | 0.12 | 3.77 | 0.67 | 0.36 | |
| Lactose | 4.76 | 0.17 | 4.61 | 0.26 | 0.34 | |
| Dry matter | 11.47 | 0.54 | 11.56 | 1.42 | 0.90 | |
| Fat free dry matter | 9.25 | 0.30 | 9.37 | 0.59 | 0.71 | |
| Cholesterol (μg/100 mL) | 10.67 | 2.021 | 12.83 | 4.875 | 0.49 | |
| Mmol/L | CON | EPA + DHA | p-Value | |||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||
| Cows | Glucose | 3.28 | 0.33 | 3.03 | 0.39 | 0.258 |
| Total Cholesterol | 3.21 a | 0.24 | 2.90 b | 0.14 | 0.025 | |
| HDL Cholesterol | 2.14 | 0.51 | 1.93 | 0.18 | 0.388 | |
| LDL Cholesterol | 1.24 | 0.36 | 1.05 | 0.14 | 0.269 | |
| Triacylglycerol | 0.15 | 0.06 | 0.14 | 0.04 | 0.687 | |
| Calves | Glucose | 6.47 | 0.35 | 6.70 | 0.57 | 0.457 |
| Total Cholesterol | 2.88 | 0.28 | 2.57 | 0.40 | 0.209 | |
| HDL Cholesterol | 1.95 | 0.52 | 1.58 | 0.33 | 0.347 | |
| LDL Cholesterol | 1.00 | 0.10 | 0.98 | 0.19 | 0.829 | |
| Triacylglycerol | 0.43 | 0.29 | 0.49 | 0.32 | 0.767 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brozić, D.; Starčević, K.; Vranić, M.; Bošnjak, K.; Maurić Maljković, M.; Mašek, T. Effect of Dietary Eicosapentaenoic and Docosahexaenoic Fatty Acid Supplementation during the Last Month of Gestation on Fatty Acid Metabolism and Oxidative Status in Charolais Cows and Calves. Animals 2024, 14, 1273. https://doi.org/10.3390/ani14091273
Brozić D, Starčević K, Vranić M, Bošnjak K, Maurić Maljković M, Mašek T. Effect of Dietary Eicosapentaenoic and Docosahexaenoic Fatty Acid Supplementation during the Last Month of Gestation on Fatty Acid Metabolism and Oxidative Status in Charolais Cows and Calves. Animals. 2024; 14(9):1273. https://doi.org/10.3390/ani14091273
Chicago/Turabian StyleBrozić, Diana, Kristina Starčević, Marina Vranić, Krešimir Bošnjak, Maja Maurić Maljković, and Tomislav Mašek. 2024. "Effect of Dietary Eicosapentaenoic and Docosahexaenoic Fatty Acid Supplementation during the Last Month of Gestation on Fatty Acid Metabolism and Oxidative Status in Charolais Cows and Calves" Animals 14, no. 9: 1273. https://doi.org/10.3390/ani14091273