

On Segurasaurus (Squamata: Pythonomorpha), a New Genus of Lizard from the Cenomanian (Upper Cretaceous) of Portugal
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Location and Stratigraphic and Palaeoenvironmental Contexts
3. Material and Methods
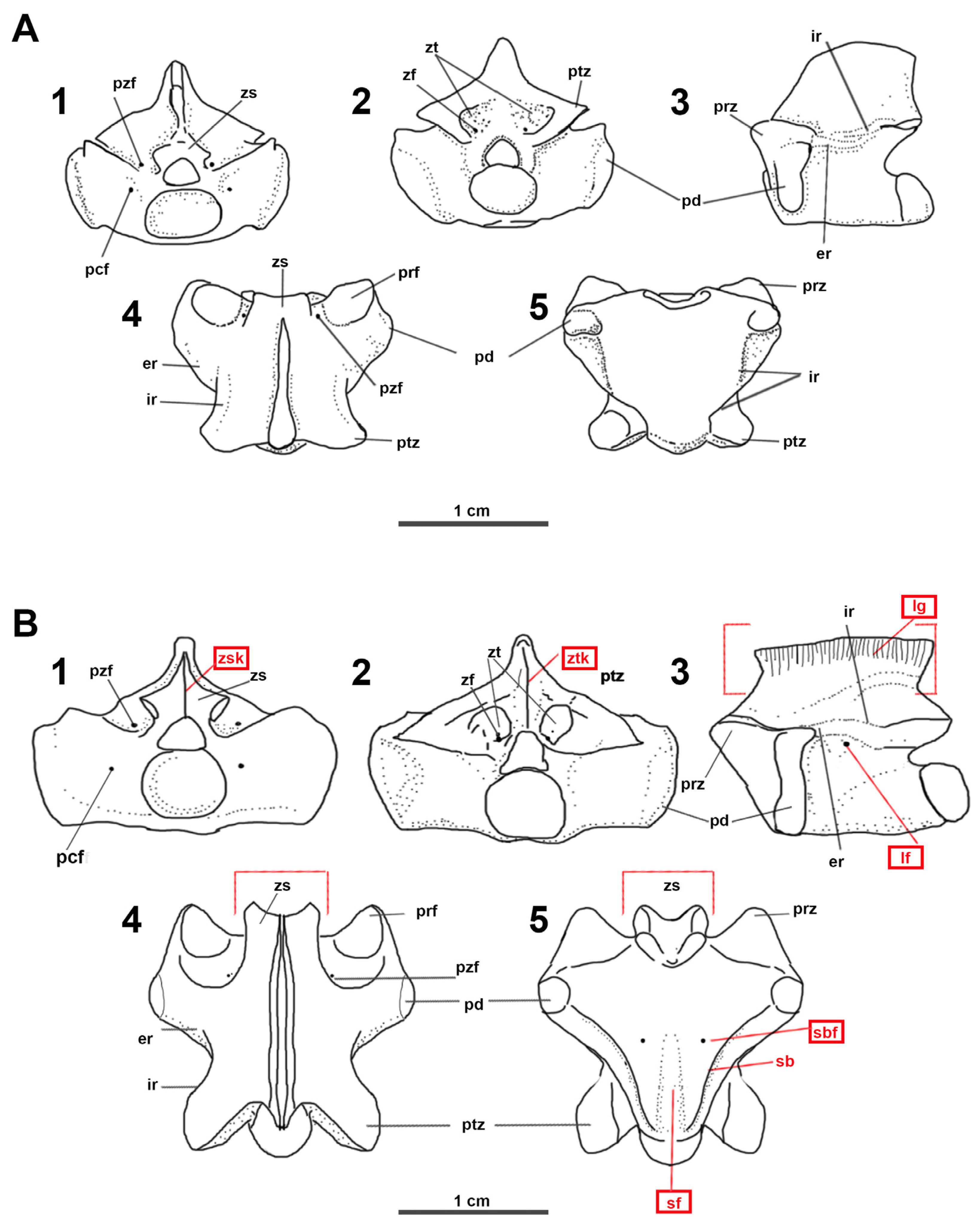
3.1. Vertebrae Measurement Abbreviations
3.2. Vertebrae Anatomical Abbreviations
4. Systematic Palaeontology (by Mélani Berrocal-Casero, Ricardo Pimentel, and Pedro M. Callapez)
4.1. Diagnosis
4.2. Distribution
4.3. Type Locality
4.4. Etymology
4.5. Material Description
Description of the New Material (DCT-CC-35)
5. Discussion
5.1. Taxonomic Remarks
5.2. Intracolumnar Position of the New Material
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rage, J.C.; Néraudeau, D. A new pachyostotic squamate reptile from the Cenomanian of France. Palaeontology 2004, 47, 1195–1210. [Google Scholar] [CrossRef]
- Pimentel, R.J.; Audije-Gil, J.; Barroso-Barcenilla, F.; Berrocal-Casero, M.; Callapez, P.M.; Ozkaya de Juanas, S.; Segura, M. On the occurrence of Carentonosaurus Rage and Néraudeau, 2004 (Squamata, Pythonomorpha) in the Iberian Cretaceous, with description of a new species from the uppermost middle Cenomanian of West Portugal. Cretac. Res. 2023, 151, 105469. [Google Scholar] [CrossRef]
- Bardet, N.; Houssaye, A.; Rage, J.C.; Pereda Suberbiola, X. The Cenomanian-Turonian (late Cretaceous) radiation of marine squamates (Reptilia): The role of the Mediterranean Tethys. Bull. Soc. Géol. Fr. 2008, 179, 605–622. [Google Scholar] [CrossRef]
- Vullo, R.; Néraudeau, D. Cenomanian vertebrate assemblages from southwestern France: A new insight into the European mid-Cretaceous continental fauna. Cret. Res. 2008, 29, 935–940. [Google Scholar] [CrossRef]
- Vullo, R.; Rage, J.C.; Néraudeau, D. Anuran and Squamate remains from the Cenomanian (Late Cretaceous) of Charentes, Western France. J. Vert. Paleont. 2011, 31, 279–291. [Google Scholar] [CrossRef]
- Vullo, R.; Bernárdez, E.; Buscalioni, A. Vertebrates from the middle?-late Cenomanian La Cabaña Formation (Asturias, northern Spain): Palaeoenvironmental and palaeobiogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 276, 120–129. [Google Scholar] [CrossRef]
- Hontecillas, D.; Knoll, F.; Arsuaga, J.L.; Laplana, C.; Pérez-González, A.; Baquedano, E. Reworked remains of mosasauroids (Pythonomorpha, Squamata) in the Pleistocene of Pinilla del Valle (Madrid, Spain). In Proceedings of the 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; Royo-Torres, R., Gascó, F., Alcalá, L., Eds.; Conjunto Paleontológico de Teruel—Dinópolis: Teruel, Spain, 2012; p. 113. [Google Scholar]
- Hontecillas, D.; Houssaye, A.; Laplana, C.; Sevilla, P.; Arsuaga, J.L.; Pérez-González, A.; Baquedano, E.; Knoll, F. Reworked marine pythonomorph (Reptilia, Squamata) remains in Late Pleistocene cave deposits in central Spain. Cret. Res. 2015, 54, 188–202. [Google Scholar] [CrossRef]
- Cabezuelo-Hernández, A.; Pérez-García, A. A new species of the Pythonomorph Carentonosaurus from the Cenomanian of Algora (Guadalajara, Central Spain). Animals 2023, 13, 1197. [Google Scholar] [CrossRef]
- Barbosa, B.; Soares, A.F.; Rocha, R.B.; Manuppella, G.; Henriques, M. Carta Geológica de Portugal, Escala 1:50 000. Notícia Explicativa da Folha 19A—Cantanhede; Serviços Geológicos de Portugal: Lisboa, Portugal, 1988; pp. 1–46. [Google Scholar]
- Callapez, P.M. Palaeobiogeographic evolution and marine faunas of the Mid-Cretaceous Western Portuguese Carbonate Platform. Thalassas 2008, 24, 29–52. [Google Scholar]
- Choffat, P.L. Recueil de Monographies Stratigraphiques sur le Système Crétacique du Portugal—Deuxième Étude—Le Crétacé Supérieur au Nord du Tage; Direction des Services Géologiques du Portugal: Lisbonne, Portugal, 1900; pp. 1–287. [Google Scholar]
- Soares, A.F. Estudo das formações pós-jurássicas das regiões de entre Sargento-Mor e Montemor-o-Velho (margem direita do Rio Mondego). Mem. Notíc. 1966, 62, 1–343. [Google Scholar]
- Soares, A.F. Contribuição para o estudo do Cretácico em Portugal (o Cretácico superior da Costa de Arnes). Mem. Notíc. 1972, 74, 1–56. [Google Scholar]
- Soares, A.F. A “Formação Carbonatada” na região do Baixo-Mondego. Com. Serv. Geol. Port. 1980, 66, 99–109. [Google Scholar]
- Callapez, P.M. Estratigrafia e Paleobiologia do Cenomaniano-Turoniano. O significado do eixo da Nazaré-Leiria-Pombal. Ph.D. Thesis, University of Coimbra, Coimbra, Portugal, 1998. [Google Scholar]
- Callapez, P.M. The Cenomanian-Turonian central West Portuguese carbonate platform. In Cretaceous and Cenozoic Events in West Iberia Margins Field Trip Guidebook 2; Dinis, J., Cunha, P.P., Eds.; FCTUC: Coimbra, Portugal, 2004; pp. 39–51. [Google Scholar]
- Barroso-Barcenilla, F.; Callapez, P.M.; Soares, A.F.; Segura, M. Cephalopod assemblages and depositional sequences from the upper Cenomanian and lower Turonian of the Iberian Peninsula (Spain and Portugal). J. Iber. Geol. 2011, 37, 9–28. [Google Scholar]
- Segura, M.; Barroso-Barcenilla, F.; Callapez, P.M.; García-Hidalgo, J.F.; Gil-Gil, J. Depositional sequences and cephalopod assemblages in the upper Cenomanian-lower Santonian of the Iberian Peninsula (Spain and Portugal). Geol. Acta 2014, 12, 19–27. [Google Scholar]
- Berthou, P.Y. Albian-Turonian stage boundaries and subdivisions in the Western Portuguese Basin, with special emphasis on the Cenomanian-Turonian boundary in the Ammonite Facies and Rudist Facies. Bull. Geol. Soc. Denmark 1984, 33, 41–45. [Google Scholar] [CrossRef]
- Lauverjat, J. Le Crétacé Supérieur dans le Nord du Bassin Occidental Portugais. Ph.D. Thesis, Université Pierre et Marie Curie, Paris, France, 1982. [Google Scholar]
- Mateus, O.; Puértolas-Pascual, E.; Callapez, P.M. A new eusuchian crocodylomorph from the Cenomanian (Late Cretaceous) of Portugal reveals novel implications on the origin of Crocodylia. Zool. J. Linn. Soc. 2018, 186, 521–528. [Google Scholar] [CrossRef]
- Auffenberg, W. The fossil snakes of Florida. Tulane Stud. Zool. 1963, 10, 131–216. [Google Scholar]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis, 10th ed.; Laurentius Salvius: Holmiae, Sweden, 1758; Volume 1, pp. 1–824. [Google Scholar]
- Oppel, M. Suite du 1er mémoire sur la classification des reptiles. Ord. II. Squammata mihi. Sect. II. Ophidii. Ord. III. Ophidii, Brongniart. Ann. Mus. Hist. Nat. 1811, 16, 376–393. [Google Scholar]
- Cope, E.D. On the reptilian orders Pythonomorpha and Streptosauria. Proc. Boston Soc. Nat. Hist. 1869, 12, 251–266. [Google Scholar]
- Tschopp, E. Nomenclature of vertebral laminae in lizards, with comments on ontogenetic and serial variation in Lacertini (Squamata, Lacertidae). PLoS ONE 2016, 11, e0149445. [Google Scholar] [CrossRef] [PubMed]
- Čerňanský, A.; Yaryhin, O.; Ciceková, J.; Werneburg, I.; Hain, M.; Klembara, J. Vertebral comparative anatomy and morphological differences in Anguine Lizards with a special reference to Pseudopus apodus. Anat. Rec. 2019, 302, 232–257. [Google Scholar] [CrossRef] [PubMed]
- Georgalis, G.L.; Scheyer, T.M. Lizards and snakes from the earliest Miocene of Saint-Gérand-le-Puy, France: An anatomical and histological approach of some of the oldest Neogene squamates from Europe. BMC Ecol. Evol. 2021, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Houssaye, A. A new aquatic pythonomorph (Reptilia, Squamata) from the Turonian (Late Cretaceous) of France. Comptes Rendus Palevol 2010, 9, 39–45. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Specimen | MH | ML | cL | ctH | ctW | cdH | cdW | naW | nsL | przD | PdD | ptzD | zsW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DCT-CC-35 | 8.91 | 12.55 | 10.26 | 3.21 | 4.08 | 2.97 | 3.82 | 7.91 | 10.62 | 9.88 | 13.06 | 9.68 | 3.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berrocal-Casero, M.; Pimentel, R.; Callapez, P.M.; Barroso-Barcenilla, F.; Ozkaya de Juanas, S. On Segurasaurus (Squamata: Pythonomorpha), a New Genus of Lizard from the Cenomanian (Upper Cretaceous) of Portugal. Geosciences 2024, 14, 84. https://doi.org/10.3390/geosciences14030084
Berrocal-Casero M, Pimentel R, Callapez PM, Barroso-Barcenilla F, Ozkaya de Juanas S. On Segurasaurus (Squamata: Pythonomorpha), a New Genus of Lizard from the Cenomanian (Upper Cretaceous) of Portugal. Geosciences. 2024; 14(3):84. https://doi.org/10.3390/geosciences14030084
Chicago/Turabian StyleBerrocal-Casero, Mélani, Ricardo Pimentel, Pedro Miguel Callapez, Fernando Barroso-Barcenilla, and Senay Ozkaya de Juanas. 2024. "On Segurasaurus (Squamata: Pythonomorpha), a New Genus of Lizard from the Cenomanian (Upper Cretaceous) of Portugal" Geosciences 14, no. 3: 84. https://doi.org/10.3390/geosciences14030084