

Foraminiferal and Palynological Records of an Abrupt Environmental Change at the Badenian/Sarmatian Boundary (Middle Miocene): A Case Study in Northeastern Central Paratethys
 ,
,
Abstract
:1. Introduction
2. Geological Setting
3. Materials and Methods
- Spiniferites morphogroup, including all the Spiniferites and Achomosphaera specimens;
- Batiacasphaera morphogroup, including B. sphaerica, Batiacasphaera sp. A, B. micropapillata, and B. hirsuta;
- Operculodinium morphogroup, including all the Operculodinium specimens;
- Systematophora morphogroup, including S. placacantha and S. ?ancyrea specimens. The latter differed from S. placacantha because of incomplete or absent proximal ridges;
- Lingulodinium morphogroup, including Lingulodinium machaerophorum and a single specimen of Lingulodinium sp. A;
- Nematosphaeropsis morphogroup, including N. labyrinthus;
- Impagidinium morphogroup, including Impagidinium sp. and I. strialatum specimens;
- Polysphaeridium morphogroup, including P. subtile and P. zoharyi.
- Black, opaque phytoclasts representing the most resistant coalified land plant remains;
- Dark-brown phytoclasts, which are land plant remains, usually equidimensional, translucent (or with translucent edges), remaining in structureless wood or cortical tissues;
- Cuticle group, including structured land plant tissues and remains with usually preserved cell structures. These are commonly the remains of the leaf epidermis;
- Pollen grains of angiosperms and gymnosperms. The most common are bisaccate pollen grains of gymnosperms;
- Pteridophyte spores;
- Dinoflagellate cysts;
- Algae and the acritarch group, including algae other than dinoflagellate cysts, such as prasinophyte (e.g., Tasmanites, Cymatiosphaera, and Leiosphaera), and incertae sedis microfossils (i.e., acritarchs);
- Foraminifera organic linings (zooclasts), which are the organic cells of foraminifera;
- Amorphous organic matter (AOM), which represents structureless particles of uncertain origins (most likely marine origin in the case of the present study materials), representing a stage in the bacterial decay of organic particles in oxygen-depleted environments.
4. Results
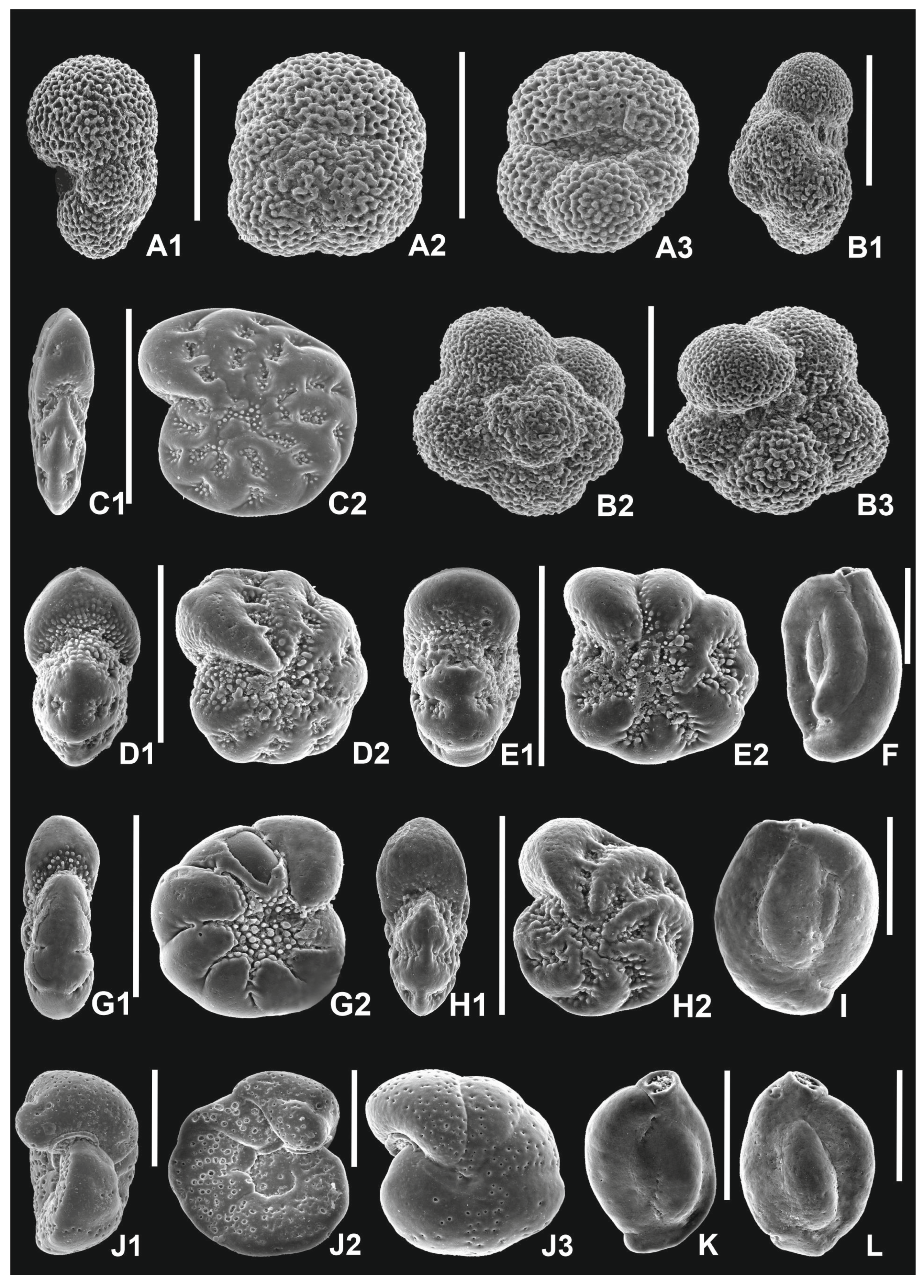
4.1. Foraminifera
4.2. Palynofacies
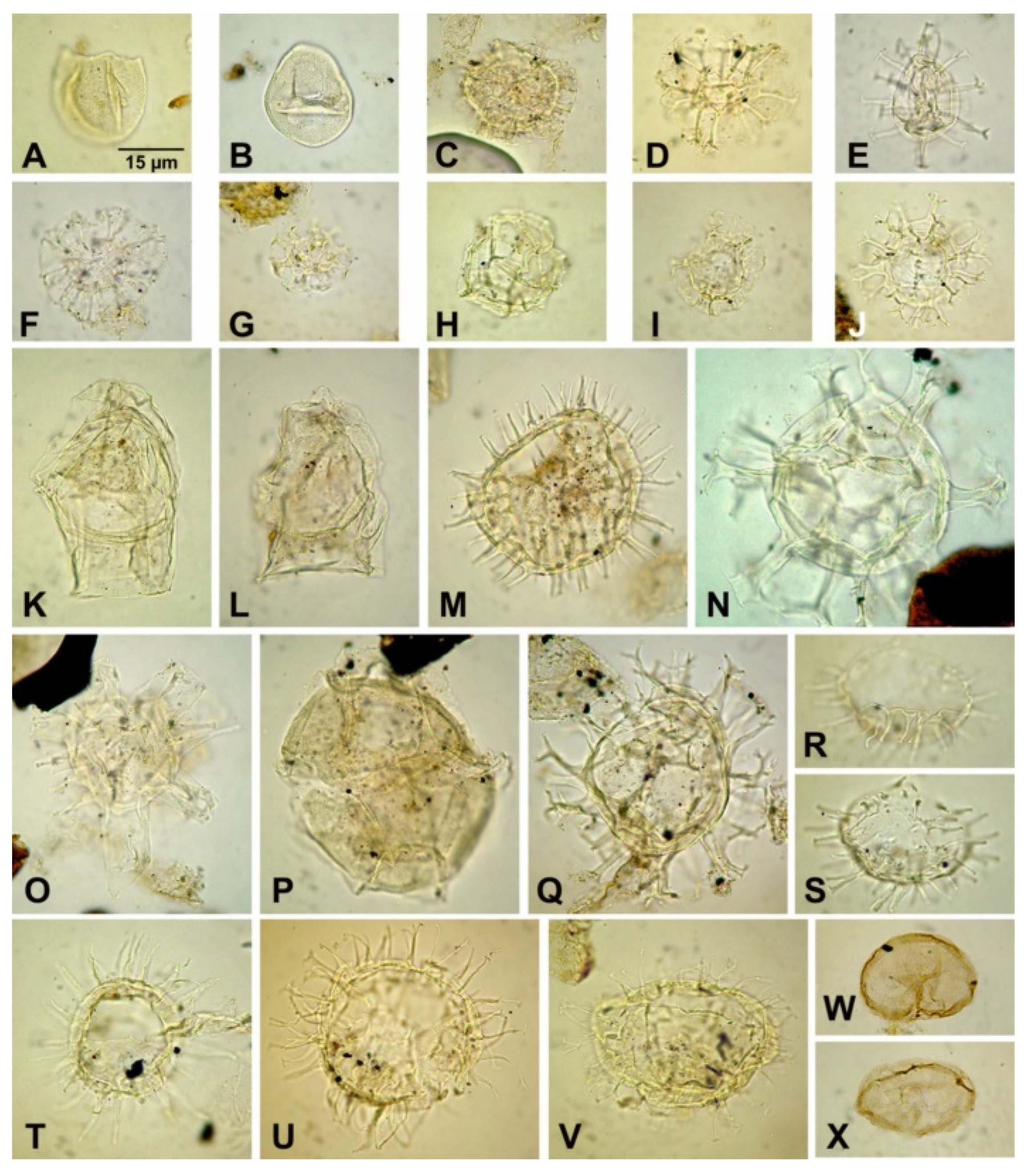
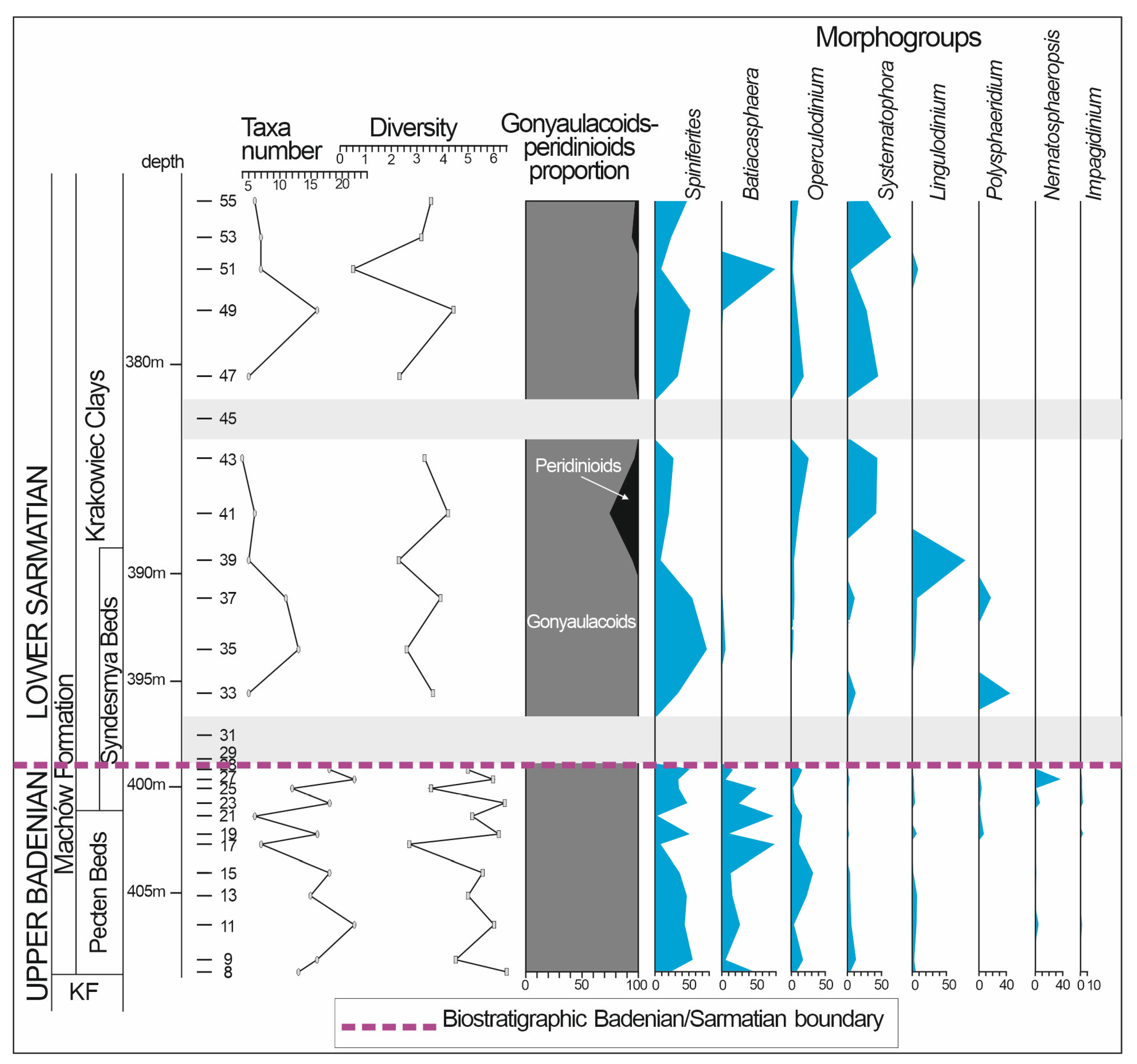
4.3. Dinoflagellate Cysts
4.4. Other Aquatic Palynomorphs
4.5. Spore–Pollen Analysis
5. Discussion
5.1. Aquatic Palaeoenvironment
5.1.1. Foraminiferal Record
5.1.2. Palynomorph Record
5.2. Terrestrial Palaeoenvironment
5.3. Implications
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Laskarev, V. Sur les equivalentes du Sarmatien supérieur en Serbie. In Recueil de Traveaux Ofert a M. Jovan Cvijic par ses amis et Collaborateurs; Vujević, P., Ed.; Državna Štamparija: Beograd, Serbia, 1924; pp. 73–85. [Google Scholar]
- Harzhauser, M.; Piller, W.E. Benchmark data of a changing sea—palaeogeography, palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 253, 8–31. [Google Scholar] [CrossRef]
- Śliwiński, M.; Bąbel, M.; Nejbert, K.; Olszewska-Nejbert, D.; Gąsiewicz, A.; Schreiber, B.C.; Be-Nowitz, J.A.; Layer, P. Badenian–Sarmatian chronostratigraphy in the Polish Carpathian Foredeep. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 326–328, 12–29. [Google Scholar] [CrossRef]
- Palcu, D.V.; Kouwenhoven, T.J.; Krijgsman, W. Reply to “Comment on the Badenian-Sarmatian extinction event in the Carpathian foredeep basin of Romania: Paleogeographic changes in the Paratethys (Palcu et al., 2015)” by Silye and Filipescu (2016). Glob. Planet. Chang. 2016, 145, 141–142. [Google Scholar] [CrossRef]
- Silye, L.; Filipescu, S. Comment on “The Badenian-Sarmatian Extinction Event in the Carpathian foredeep basin of Romania: Paleogeographic changes in the Paratethys domain” (Palcu et al., 2015). Glob. Planet. Chang. 2016, 145, 17–19. [Google Scholar] [CrossRef]
- Peryt, T.M.; Kasprzyk, A. Carbonate-evaporite sedimentary transitions in the Badenian (middle Miocene) basin of southern Poland. Sediment. Geol. 1992, 76, 257–271. [Google Scholar] [CrossRef]
- Peryt, D.; Gedl, P.; Peryt, T.M. Marine transgression(s) to evaporite basin: The case of middle Miocene (Badenian) gypsum in the Central Paratethys, SE Poland. J. Palaeogeogr. 2020, 9, 16. [Google Scholar] [CrossRef]
- Peryt, D.; Garecka, M.; Peryt, T.M. Foraminiferal and calcareous nannoplankton biostratigraphy of the upper Badenian–lower Sarmatian strata in the SE Polish Carpathian Foredeep. Geol. Quart. 2021, 65, 18. [Google Scholar] [CrossRef]
- Łuczkowska, E. Foraminiferal zones in the Miocene, south of the Holy Cross Mts. Bull. Acad. Pol. Sci. Sér. Sci. Géol. Géogr. 1963, 11, 29–34. [Google Scholar]
- Łuczkowska, E. The micropaleontological stratigraphy of the Miocene in the region of Tarnobrzeg-Chmielnik (in Polish with English summary). Pr. Geol. 1964, 20, 1–72. [Google Scholar]
- Łuczkowska, E. A new zone with Praeorbulina indigena (Foraminiferida, Globigerinidae) in the Upper Badenian (Tortonian s.s.) of Central Paratethys. Roczn. Polsk. Tow. Geol. 1971, 40, 445–448. [Google Scholar]
- Odrzywolska-Bieńkowa, E. Micropaleontological stratigraphy of the Miocene of central part of the Carpathian Foredeep (in Polish with English summary). Prz. Geol. 1975, 23, 597–603. [Google Scholar]
- Czepiec, I. Biostratigraphy and paleoenvironment of Sarmatian marginal zone of Poland (in Polish with English summary). Kwart. AGH Geologia 1996, 22, 309–338. [Google Scholar]
- Kirchner, Z. Miocene stratigraphy of the Central Carpathian Foreland based on microfaunal studies (in Polish with English summary). Acta Geol. Pol. 1956, 6, 421–450. [Google Scholar]
- Cicha, I.; Rögl, F.; Rupp, C.; Čtyroká, J. (Eds.) Oligocene-Miocene foraminifera of the Central Paratethys. Abh. Senckenberg. Naturforsch. Ges. 1998, 549, 1–325. [Google Scholar]
- Palcu, D.V.; Golovina, L.A.; Vernyhorova, Y.V.; Popov, S.V.; Krijgsman, W. Middle Miocene paleoenvironmental crises in Central Eurasia caused by changes in marine gateway configuration. Glob. Planet. Chang. 2017, 158, 57–71. [Google Scholar] [CrossRef]
- Odrzywolska-Bieńkowa, E. Porównanie wybranych głębszych profile mikrofaunistycznych rejonu świętokrzyskiego i lubelskiego (In Polish). Sprawozd. Posiedz. Kom. Nauk. PAN Kraków 1972, 16, 493–494. [Google Scholar]
- Filipescu, S. Anomalinoides dividens bioevent at the Badenian/Sarmatian boundary—a response to paleogeographic and paleoenvironmental changes. Studia Univ. Babeş-Bolyai Geologia 2004, 49, 21–26. [Google Scholar] [CrossRef]
- Dumitriu, S.D.; Loghin, S.; Dubicka, Z.; Melinte-Dobrinescu, M.C.; Paruch-Kulczycka, J.; Ionesi, V. Foraminiferal, ostracod, and calcareous nannofossil biostratigraphy of the latest Badenian—Sarmatian interval (Middle Miocene, Paratethys) from Poland, Romania and the Republic of Moldova. Geol. Carpath. 2017, 68, 419–444. [Google Scholar] [CrossRef]
- Ney, R.; Burzewski, W.; Bachleda, T.; Górecki, W.; Jakóbczak, K.; Słupczyński, K. Outline of paleogeography and evolution of lithology and facies of Neogene layers on the Carpathian Foredeep (in Polish with English summary). Prace Geol. 1974, 82, 3–65. [Google Scholar]
- Dziadzio, P.; Maksym, A.; Olszewska, B. Miocene deposition in the eastern part of the Carpathian Foredeep in Poland (In Polish with English summary). Prz. Geol. 2006, 54, 413–420. [Google Scholar]
- Alexandrowicz, S.W.; Garlicki, A.; Rutkowski, J. Podstawowe jednostki litostratygraficzne miocenu zapadliska przedkarpackiego (In Polish). Kwart. Geol. 1982, 26, 470–471. [Google Scholar]
- Peryt, T.M. The beginning, development and termination of the Middle Miocene Badenian salinity crisis in Central Paratethys. Sediment. Geol. 2006, 188–189, 379–396. [Google Scholar] [CrossRef]
- de Leeuw, A.; Tulbure, M.; Kuiper, K.F.; Melinte-Dobrinescu, M.C.; Stoica, M.; Krijgsman, W. New 40Ar/39Ar, magnetostratigraphic and biostratigraphic constraints on the termination of the Badenian Salinity Crisis: Indications for tectonic improvement of basin interconnectivity in Southern Europe. Glob. Planet. Chang. 2018, 169, 1–15. [Google Scholar] [CrossRef]
- Pawłowska, K.; Kubica, B. Karta otworu wiertniczego Babczyn-2 (in Polish; unpublished); Archive CUG: Warszawa, Poland, 1960. [Google Scholar]
- Jasionowski, M.; Peryt, T.M. Historia Badań. In Budowa geologiczna Polski, tom I. Stratygrafia, część 3a Kenozoik Paleogen Neogen; Peryt, T.M., Piwocki, M., Eds.; Polish Geological Institute: Warszawa, Poland, 2004; pp. 203–212. [Google Scholar]
- Czapowski, G. Wschodnia Część Zapadliska. In Budowa geologiczna Polski, tom I. Stratygrafia, część 3a Kenozoik Paleogen Neogen; Peryt, T.M., Piwocki, M., Eds.; Polish Geological Institute: Warszawa, Poland, 2004; pp. 239–245. [Google Scholar]
- Buzas, H.A.; Gibson, T.G. Species diversity: Benthonic Foraminifera in Western North Atlantic. Science 1969, 163, 72–75. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006; p. 426. [Google Scholar]
- Murray, J.W. Ecology and Palaeoecology of Benthic Foraminifera; Routledge: London, UK, 1991; p. 408. [Google Scholar]
- Van der Zwaan, G.J. Paleoecology of late Miocene Mediterranean Foraminifera. Utrecht Micropaleont. Bull. 1982, 25. [Google Scholar]
- Corliss, B.; Chen, C. Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implication. Geology 1988, 16, 716–719. [Google Scholar] [CrossRef]
- Culver, S. New foraminiferal depth zonation of the northwestern Gulf of Mexico. Palaios 1988, 3, 69–85. [Google Scholar] [CrossRef]
- Lutze, G.F.; Thiel, H. Epibenthic foraminifera from elevated microhabitats; Cibicidoides wuellerstorfi and Planulina ariminensis. J. Foram. Res. 1989, 19, 153–158. [Google Scholar] [CrossRef]
- Verhallen, P. Late Pliocene to early Pleistocene Mediterranean mud-dwelling foraminifera; influence of a changing environment on community structure and evolution. Utrecht Micropaleont. Bull. 1991, 40, 1–219. [Google Scholar]
- Sjoerdsma, P.G.; van der Zwaan, G.J. Simulating the effect of changing oceanic flux and oxygen content on the distribution of benthic foraminifers. Marine Micropaleont. 1992, 19, 163–180. [Google Scholar] [CrossRef]
- Langer, M. Epiphytic foraminifera. Mar. Micropaleont. 1993, 20, 235–265. [Google Scholar] [CrossRef]
- Hayward, B.W.; Hollis, C.J.; Grenfell, H.R. Recent Elphidiidae (Foraminiferida) of the South-west Pacific and fossil Elphidiidae of New Zealand. Inst. Geol. Nucl. Sci. Mon. 1997, 16, 170. [Google Scholar]
- Kaiho, K. Effect of organic carbon flux and dissolved oxygen on the benthic foraminiferal oxygen index (BFOI). Mar. Micropaleont. 1999, 37, 67–76. [Google Scholar] [CrossRef]
- Schönfeld, J. A new benthic foraminiferal proxy for near-bottom current velocities in the Gulf of Cadiz, northeastern Atlantic Ocean. Deep-Sea Res. I 2002, 49, 1853–1875. [Google Scholar] [CrossRef]
- Hohenegger, J. Estimation of environmental paleogradient values based on presence/absence data: A case study using benthic foraminifera for paleodepth estimation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 217, 115–130. [Google Scholar] [CrossRef]
- Kouwenhoven, T.J.; van der Zwaan, G.J. A reconstruction of late Miocene Mediterranean circulation patterns using benthic foraminifera. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 373–385. [Google Scholar] [CrossRef]
- Kaminski, M.A. Calibration of the Benthic Foraminiferal Oxygen Index in the Marmara Sea. Geol. Quart. 2012, 56, 757–764. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Planktic Foraminifers in the Modern Ocean; Springer: Berlin, Germany, 2017; p. 358. [Google Scholar]
- Jorissen, F.; Nardelli, M.P.; Almogi-Labin, A.; Barras, C.; Bergamin, L.; Bicchi, E.; El Kateb, A.; Ferraro, L.; McGann, M.; Morigi, C.; et al. Developing Foram-AMBI for biomonitoring in the Mediterranean: Species assignments to ecological categories. Mar. Micropaleont. 2018, 140, 33–45. [Google Scholar] [CrossRef]
- Dumitriu, S.D.; Dubicka, Z.; Loghin, S.; Melinte-Dobrinescu, M.C.; Paruch-Kulczycka, J. The evolution of the Carpathian Foredeep Basin during the latest Badenian and Sarmatian (Middle Miocene): Inferences from micropalaeontological data. Geol. Quart. 2020, 64, 1004–1022. [Google Scholar] [CrossRef]
- Cavaliere, M.; Scipioni, V.; Francescangeli, F.; Ferraro, L.; Frontalini, F. Paleoenvironmental changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A perspective from benthic foraminifera after dam construction. Water 2023, 15, 815. [Google Scholar] [CrossRef]
- Kiss, P.; Hudáčková, N.; Titschack, J.; Siccha, M.G.R.; Heřmanová, Z.; Silye, L.; Ruman, A.; Rybár, S.; Kučera, M. Convergent evolution of spherical shells in Miocene planktonic foraminifera documents the parallel emergence of a complex character in response to environmental forcing. Paleobiology 2023, 49, 454–470. [Google Scholar] [CrossRef]
- Van der Zwaan, G.J.; Duijnstee, I.A.P.; den Dulk, M.; Ernst, S.R.; Jannink, N.T.; Kouwenhoven, T.J. Benthic foraminifers: Proxies or problems? A review of paleoecological concepts. Earth-Sci. Rev. 1999, 46, 213–236. [Google Scholar] [CrossRef]
- Van Hinsbergen, D.J.J.; Kouwenhoven, T.J.; van der Zwaan, G.J. Paleobathymetry in the backstripping procedure: Correction for oxygenation effects on depth estimates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 221, 245–265. [Google Scholar] [CrossRef]
- Holcová, K.; Zágoršek, K. Bryozoa, foraminifera and calcareous nannoplankton as environmental proxies of the "bryozoan event" in the Middle Miocene of the Central Paratethys (Czech Republic). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 267, 216–234. [Google Scholar] [CrossRef]
- Hart, M.B.; Fisher, J.K.; Smart, C.W.; Speers, R.; Wall-Palmer, D. Re-colonization of hostile environments by benthic foraminifera: An example from Montserrat, Lesser Antilles Volcanic Arc. Micropaleontology 2022, 69, 1–27. [Google Scholar] [CrossRef]
- Stuchlik, L.; Ziembińska-Tworzydło, M.; Kohlman-Adamska, A.; Grabowska, I.; Ważyńska, H.; Słodkowska, B.; Sadowska, A. Atlas of Pollen and Spores of the Polish Neogene. Vol. 1—Spores; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2001; p. 158. [Google Scholar]
- Stuchlik, L.; Ziembińska-Tworzydło, M.; Kohlman-Adamska, A.; Grabowska, I.; Ważyńska, H.; Sadowska, A. Atlas of Pollen and Spores of the Polish Neogene. Vol. 2—Gymnosperms; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; p. 237. [Google Scholar]
- Stuchlik, L.; Ziembińska-Tworzydło, M.; Kohlman-Adamska, A.; Grabowska, I.; Słodkowska, B.; Ważyńska, H.; Sadowska, A. Atlas of pollen and spores of the Polish Neogene. Vol. 3—Angiosperms (1); W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2009; p. 233. [Google Scholar]
- Stuchlik, L.; Ziembińska-Tworzydło, M.; Kohlman-Adamska, A.; Grabowska, I.; Słodkowska, B.; Worobiec, E.; Durska, E. Atlas of pollen and spores of the Polish Neogene. Vol. 4—Angiosperms (2); W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2014; p. 466. [Google Scholar]
- Nalepka, D.; Walanus, A. Data processing in pollen analysis. Acta Palaeobot. 2003, 43, 125–134. [Google Scholar]
- Corliss, B. Microhabitats of benthic foraminifera within deep-sea sediments. Nature 1985, 314, 435–438. [Google Scholar] [CrossRef]
- Schiebel, R.; Bijma, J.; Hemleben, C. Population dynamics of the planktic foraminifer Globigerina bulloides from the eastern North Atlantic. Deep-Sea Res. I 1997, 44, 1701–1713. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Modern planktonic foraminifera. Paläont. Zt. 2005, 79, 135–148. [Google Scholar] [CrossRef]
- Caron, M. Cretaceous planktic foraminifera. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Nielsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 17–86. [Google Scholar]
- Hemleben, C.; Spindler, M.; Anderson, O.R. Modern Planktonic Foraminifera. Springer: New York, NY, USA, 1989; p. 363. [Google Scholar]
- Hebbeln, D.; Marchant, M.; Wefer, G. Seasonal variations of the particle flux in the Peru-Chile Current at 30°S under normal and El Nino conditions. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 2101–2128. [Google Scholar] [CrossRef]
- Petrizzo, M.R. Palaeoceanographic and palaeoclimatic inferences from Late Cretaceous planktonic foraminiferal assemblages from the Exmouth Plateau (ODP Sites 762 and 763, eastern Indian Ocean). Mar. Micropaleontol. 2002, 45, 117–150. [Google Scholar] [CrossRef]
- Kretschmer, K.; Jonkers, L.; Kucera, M.; Schulz, M. Modeling seasonal and vertical habitats of planktonic foraminifera on a global scale. Biogeosciences 2018, 15, 4405–4429. [Google Scholar] [CrossRef]
- Di Stefano, A.; Verducci, M.; Lirer, F.; Ferraro, L.; Iaccarino, S.M.; Hüsing, S.K.; Hilgen, F.J. Paleoenvironmental conditions preceding the Messinian Salinity Crisis in the Central Mediterranean: Integrated data from the Upper Miocene Trave section (Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 37–53. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Tolderlund, D.S. Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian Oceans. In The Micropaleontology of Oceans; Funnell, B.M., Riedel, W.R., Eds.; Cambridge University Press: Cambridge, UK, 1971; pp. 105–149. [Google Scholar]
- Bé, A.W.H. An ecological zoogeographic and taxonomic review of recent planktonic foraminifera. In The Micropaleontology of Oceans; Ramsay, A.T.S., Ed.; Academic Press: London, UK, 1977; pp. 1–100. [Google Scholar]
- Thunell, R.; Reynolds-Sautter, L. Planktonic foraminiferal faunal and stable isotopic indices of upwelling: A sediment trap study in the San Pedro Basin, Southern California Bight. Geol. Soc. Spec. Publ. 1992, 64, 77–91. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M.; Siccha, M.; Trommer, G.; Schulz, H.; Abdolalipour, S.; Atsushi Kurasawa, A. Genetic diversity and ecology of the planktonic foraminifers Globigerina bulloides, Turborotalita quinqueloba and Neogloboquadrina pachyderma off the Oman margin during the late SW Monsoon. Mar. Micropaleont. 2017, 137, 64–77. [Google Scholar] [CrossRef]
- Jorissen, F.J.; de Stigter, H.C.; Widmark, J.G.V. A conceptual model explaining benthic foraminiferal microhabitats. Mar. Micropaleont. 1995, 26, 3–15. [Google Scholar] [CrossRef]
- Robert, C.M. Global Sedimentology of the Ocean: An Interplay between Geodynamics and Paleoenvironment; Elsevier: Amsterdam, The Netherlands, 2008; p. 496. [Google Scholar]
- Rodrigues, A.R.; Gómez Pivel, M.A.; Schmitt, P.; de Almeida, F.K.; Bonetti, C. Infaunal and epifaunal benthic foraminifera species as proxies of organic matter paleofluxes in the Pelotas Basin, south-western Atlantic Ocean. Mar. Micropaleont. 2018, 144, 38–49. [Google Scholar] [CrossRef]
- Kováč, M.; Márton, E.; Oszczypko, N.; Vojtko, R.; Hók, J.; Králiková, S.; Plašienka, D.; Klučiar, T.; Hudáčková, N.; Oszczypko-Clowes, M. Neogene palaeogeography and basin evolution of the Western Carpathians, Northern Pannonian domain and ad joining areas. Glob. Planet. Chang. 2017, 155, 133–154. [Google Scholar] [CrossRef]
- Bukowski, K.; de Leeuw, A.; Gonera, M.; Kuiper, K.F.; Krzywiec, P.; Peryt, D. Badenian tuffite levels within the Carpathian orogenic front (Gdów–Bochnia area, Southern Poland): Radio-isotopic dating and stratigraphic position. Geol. Quart. 2010, 54, 449–464. [Google Scholar]
- Hess, S.; Kuhnt, W. Deep-sea benthic foraminiferal recolonization of the 1991 Mt Pinatubo ash layer in the South China Sea. Mar. Micropaleont. 1996, 28, 171–197. [Google Scholar] [CrossRef]
- Hess, S.; Kuhnt, W.; Hill, S.; Kaminski, M.A.; Holbourn, A.; de Leon, M. Monitoring the recolonization of the Mt Pinatubo 1991 ash layer by benthic foraminifera. Mar. Micropaleont. 2001, 43, 119–142. [Google Scholar] [CrossRef]
- Debenay, J.-P.; Geslin, E.; Eichler, B.B.; Duleba, W.; Sylvestre, F.; Eichler, P. Foraminiferal assemblages in a hypersaline lagoon, Araruama (R.J.) Brazil. J. Foram. Res. 2001, 31, 133–151. [Google Scholar] [CrossRef]
- WoRMS Editorial Board. World Register of Marine Species, 2024. Available online: https://www.marinespecies.org (accessed on 3 January 2024).
- Chaloner, W.G.; Muir, M. Spores and floras. In Coal and Coal-Bearing Strata; Murchison, D.G., Westoll, T.S., Eds.; Oliver and Boyd: Edinburgh, UK, 1968; pp. 127–146. [Google Scholar]
- Streel, M.; Richelot, C. Wind and water transport and sedimentation of miospores along two rivers subject to major floods and entering the Mediterranean Sea at Calvi (Corsica, France). In Sedimentation of Organic Particles; Traverse, A., Ed.; Cambridge University Press: Cambridge, UK, 1994; pp. 59–67. [Google Scholar]
- Brinkhuis, H. Late Eocene to Early Oligocene dinoflagellate cysts from the Priabonian type-area (northeast Italy): Biostratigraphy and palaeoenvironmental interpretation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 107, 121–163. [Google Scholar] [CrossRef]
- Stover, L.E.; Brinkhuis, H.; Damassa, S.P.; de Verteuil, L.; Helby, R.J.; Monteil, E.; Partridge, A.D.; Powell, A.J.; Riding, J.B.; Smelror, M.; et al. Mesozoic–Tertiary dinoflagellates, acritarchs and prasinophytes. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Dallas TX, USA, 1996; Volume 2, pp. 641–750. [Google Scholar]
- Sluijs, A.; Pross, J.; Brinkhuis, H. From greenhouse to icehouse; organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene. Earth-Sci. Rev. 2005, 68, 281–315. [Google Scholar] [CrossRef]
- Williams, G.L. Dinoflagellate cysts, their classification, biostratigraphy and palaeoecology. In Oceanic Micropalaeontology; Ramsay, A.T.S., Ed.; Academic Press: London, UK, 1977; pp. 1231–1325. [Google Scholar]
- Köthe, A. Paleogene dinoflagellates from northwest Germany. Geol. Jb. R. A 1990, 118, 1–111. [Google Scholar]
- Gedl, P. Middle Miocene dinoflagellate cysts from the Korytnica clays (Góry Świętokrzyskie Mountains, Poland). Ann. Soc. Geol. Pol. 1996, 66, 191–218. [Google Scholar]
- Schreck, M.; Matthiessen, J. Batiacasphaera micropapillata: Palaeobiogeographic distribution and palaeoecological implications of a critical Neogene species complex. Micropalaeont. Soc. Spec. Publ. 2013, 5, 301–314. [Google Scholar]
- Gedl, P. Palynofacies of the Miocene deposits in the Gliwice area (Upper Silesia, Poland). Bull. Pol. Acad. Sci. Earth Sci. 1997, 45, 191–201. [Google Scholar]
- Rochon, A.; de Vernal, A.; Turon, J.L.; Matthiessen, J.; Head, M.J. Distribution of recent dinoflagellate cysts in surface sediments from the North Atlantic Ocean and adjacent seas in relation to sea-surface parameters. Am. Assoc. Strat. Palynol. Found. Contrib. Ser. 1999, 35, 146. [Google Scholar]
- Schreck, M.; Meheust, M.; Stein, R.; Matthiessen, J. Response of marine palynomorphs to Neogene climate cooling in the Iceland Sea (ODP Hole 907A). Mar. Micropaleont. 2013, 101, 49–67. [Google Scholar] [CrossRef]
- Gedl, P. Palaeoenvironmental and sedimentological interpretations of the palynofacies analysis of the Miocene deposits from the Jamnica S-119 borehole (Carpathian Foredeep, Poland). Geol. Quart. 1999, 43, 479–492. [Google Scholar]
- Guy-Ohlson, D. Prasinophycean algae. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Dallas, TX, USA, 1996; Volume 1, pp. 181–189. [Google Scholar]
- Gedl, P. Dinoflagellate cysts and palynofacies from the upper Badenian (Middle Miocene) of the Roztocze area at Józefów and Żelebsko (Carpathian Foredeep Basin, Poland): Palaeoenvironmental implications. Ann. Soc. Geol. Pol. 2016, 86, 273–289. [Google Scholar] [CrossRef]
- Bradford, M.R.; Wall, D.A. The distribution of Recent organic-walled dinoflagellate cysts in the Persian Gulf, Gulf of Oman, and northeastern Arabian Sea. Palaeontogr. Abt. B 1984, 192, 16–84. [Google Scholar]
- Gedl, P.; Peryt, D. Dinoflagellate cyst, palynofacies and foraminiferal records of environmental changes related to the Late Badenian (Middle Miocene) transgression at Kudryntsi (western Ukraine). Ann. Soc. Geol. Pol. 2011, 81, 331–349. [Google Scholar]
- Peryt, D.; Gedl, P.; Peryt, T.M. Foraminiferal and palynological records of the Late Badenian (Middle Miocene) transgression in Podolia (Shchyrets near Lviv, western Ukraine). Geol. Quart. 2014, 58, 465–484. [Google Scholar] [CrossRef]
- Gedl, P.; Peryt, D.; Peryt, T.M. Foraminiferal and palynological organic matter records of the Upper Badenian (Middle Miocene) deposits at Anadoly (marginal part of the Ukrainian Carpathian Foredeep Basin). Geol. Quart. 2016, 60, 517–536. [Google Scholar] [CrossRef]
- Head, M.J. Neogene occurrences of the marine acritarch genus Nannobarbophora Habib and Knapp, 1982 emend., and the new species N. gedlii. J. Paleont. 2003, 77, 382–385. [Google Scholar] [CrossRef]
- Gedl, P. In situ and recycled dinoflagellate cysts from Middle Miocene deposits at Bęczyn, Carpathian Foredeep, Poland. Studia Geol. Pol. 2005, 124, 371–394. [Google Scholar]
- Jiménez-Moreno, G.; Head, M.J.; Harzhauser, M. Early and Middle Miocene dinoflagellate cyst stratigraphy of the Central Paratethys, Central Europe. J. Micropaleont. 2006, 25, 113–139. [Google Scholar] [CrossRef]
- Oszast, J. The Miocene vegetation of a sulphur bed at Piaseczno near Tarnobrzeg (Southern Poland) (in Polish with English summary). Acta Palaeobot. 1967, 8, 3–29. [Google Scholar]
- Sadowska, A. Sarmatian palynoflora from Jamnica near Tarnobrzeg (Carpathian Foredeep)—environmental and climatic implications. Geol. Quart. 1999, 43, 493–498. [Google Scholar]
- Schlütz, F.; Shumilovskikh, L.S. On the relation of Potamomyces armatisporus to the fossil form-type Mediaverrunites and its taxonomical and ecological implications. Fung. Ecol. 2013, 6, 309–315. [Google Scholar] [CrossRef]
- Worobiec, G.; Neumann, F.H.; Worobiec, E.; Nitz, V.; Hartkopf-Fröder, C. New fungal cephalothecoid-like fructifications from central European Neogene deposits. Fung. Biol. 2017, 121, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Worobiec, G.; Worobiec, E.; Gedl, P.; Kasiński, J.R.; Peryt, D.; Widera, M. Terrestrial-aquatic wood-inhabiting ascomycete Potamomyces from the Miocene of Poland. Acta Palaeontol. Pol. 2022, 67, 737–744. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; van Geel, B.; Wiltshire, P.E. The enigma of the Diporotheca palynomorph. Rev. Palaeobot. Palynol. 2016, 235, 94–98. [Google Scholar] [CrossRef]
- Worobiec, G.; Worobiec, E.; Gedl, P.; Kowalski, R.; Peryt, D.; Tietz, O. Fossil history of fungus host-specificity: Association of conidia of fossil Asterosporium asterospermum with macro- and microremains of Fagus. Fungal Biol. 2023, 127, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Worobiec, E.; Worobiec, G.; Gedl, P. Occurrence of fossil bamboo pollen and a fungal conidium of Tetraploa cf. aristata in Upper Miocene deposits of Józefina (Poland). Rev. Palaeobot. Palynol. 2009, 157, 211–217. [Google Scholar] [CrossRef]
- Grabowska, I.; Słodkowska, B. Katalog Profili Osadów Trzeciorzędowych Opracowanych Palinologicznie (In Polish); PIG: Warszawa, Poland, 1993; p. 80. [Google Scholar]
- Ważyńska, H. Introduction. Pr. Państw. Inst. Geol. 1998, 160, 5–8. [Google Scholar]
- Oszast, J. Pollen analysis of Tortonian clays from Stare Gliwice in Upper Silesia, Poland (In Polish with English summary). Monogr. Botan. 1960, 9, 1–47. [Google Scholar]
- Dyjor, S.; Sadowska, A. Problem of the Badenian-Sarmatian boundary at Stara Kuźnia region near Kędzierzyn (Silesia) in the light of palinological investigations (In Polish with English summary). Acta Palaeobot. 1984, 24, 27–51. [Google Scholar]
- Sadowska, A. Former palynological studies of the Tertiary in the western part of Carpathian Foredeep in Poland (In Polish with English summary). Prz. Geol. 1996, 44, 1039–1042. [Google Scholar]
- Sadowska, A. Miocene palynology in the Gliwice Region (Upper Silesia), Poland. Bull. Pol. Acad. Sci. Earth Sci. 1997, 45, 203–210. [Google Scholar]
- Durska, E. The Badenian Salinity Crisis in the palynological record: Vegetation during the evaporative event (Carpathian Foredeep, southern Poland). Ann. Soc. Geol. Pol. 2017, 87, 213–228. [Google Scholar] [CrossRef]
- Kita, Z. Palynological analysis of Tortonian deposits from the borehole Kłaj 1 (East of Kraków) (In Polish with English summary). Roczn. Polsk. Tow. Geol. 1963, 33, 517–526. [Google Scholar]
- Syabryaj, S.V.; Stuchlik, L. Palaeofloristic and palaeoclimatic reconstruction on the territories of Ukraine and Poland during the Badenian-Sarmatian. Acta Palaeobot. 2004, 44, 55–68. [Google Scholar]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Syabryaj, S.V.; Stuchlik, L. Development of flora and vegetation of the Ukrainian Eastern Carpathians and Polish Western Carpathians in the Neogene. Acta Palaeobot. 1994, 34, 165–194. [Google Scholar]
- Kvaček, Z.; Kováč, M.; Kovar-Eder, J.; Doláková, N.; Jechorek, H.; Parashiv, V.; Kováčová, M.; Sliva, L. Miocene evolution of landscape and vegetation in the Central Paratethys. Geol. Carpath. 2006, 57, 295–310. [Google Scholar]
- Hudáčková, N.; Ruman, A.; Plašienková, I.; Jamrich, M.; Halásová, E.; Kiss, P.; Kováčová, M.; Babejová-Kmecová, J.; Kováč, M. Badenian/Sarmatian foraminifera shift in the Central Paratethys: Two methods comparison—A morphogroup and species approach. In Proceedings of the Geologica Carpathica 70 Conference, Smolenice Castle, Slovakia, 9–11 October 2019; pp. 144–145. [Google Scholar]
- Palcu, D.V.; Tulbure, M.; Bartol, M.; Kouwenhoven, T.J.; Krijgsman, W. The Badenian-Sarmatian Extinction Event in the Carpathian foredeep basin of Romania: Paleogeographic changes in the Paratethys domain. Glob. Planet. Chang. 2015, 133, 346–358. [Google Scholar] [CrossRef]
- Nováková, P.; Rybár, S.; Šarinová, K.; Nagy, A.; Hudáčková, N.; Jamrich, M.; Teodoridis, V.; Kováčová, M.; Šujan, M.; Vlček, T.; et al. The late Badenian-Sarmatian (Serravallian) environmental transition calibrated by sequence stratigraphy (eastern Danube Basin, Central Paratethys). Geol. Carpath. 2020, 71, 291–313. [Google Scholar] [CrossRef]
- Popov, S.V.; Antipov, M.P.; Zastrozhnov, A.S.; Kurina, E.E.; Pinchuk, T.N. Sea-level fluctuations on the northern shelf of the Eastern Paratethys in the Oligocene–Neogene. Stratigr. Geol. Correl. 2010, 18, 200–224. [Google Scholar] [CrossRef]
- Simon, D.; Palcu, D.; Meijer, P.; Krijgsman, W. The sensitivity of middle Miocene paleoenvironments to changing marine gateways in Central Europe. Geology 2019, 47, 35–38. [Google Scholar] [CrossRef]
- Liu, S.; Krijgsman, W.; Dekkers, M.J.; Palcu, D. Early diagenetic greigite as an indicator of paleosalinity changes in the middle Miocene Paratethys Sea of central Europe. Geochem. Geophys. Geosyst. 2017, 18, 2634–2645. [Google Scholar] [CrossRef]
- Brasier, M.D. The ecology and distribution of Recent foraminifera from the reefs and shoals around Barbuda, West Indies. J. Foram. Res. 1975, 5, 193–210. [Google Scholar] [CrossRef]
- Ellegaard, M.; Lewis, J.; Harding, I. Cyst-theca relationship, life cycle, and effects of temperature and salinity on the cyst morphology of Gonyaulax baltica sp. nov. (Dinophyceae) from the Baltic Sea area. J. Phycol. 2002, 38, 775–789. [Google Scholar] [CrossRef]
- Mudie, P.J.; Marret, F.; Mertens, K.; Shumilovskikh, L.; Leroy, S.A.G. Atlas of modern dinoflagellate cyst distributions in the Black Sea Corridor: From Aegean to Aral Seas, including Marmara, Black, Azov and Caspian Seas. Mar. Micropaleont. 2017, 134, 1–152. [Google Scholar] [CrossRef]
- Piller, W.E.; Harzhauser, M. The myth of the brackish Sarmatian Sea. Terra Nova 2005, 17, 450–455. [Google Scholar] [CrossRef]
- Ivanov, D.; Worobiec, E. Middle Miocene (Badenian) vegetation and climate dynamics in Bulgaria and Poland based on pollen data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 467, 83–94. [Google Scholar] [CrossRef]
- de Leeuw, A.; Bukowski, K.; Krijgsman, W.; Kuiper, K.F. Age of the Badenian salinity crisis; impact of Miocene climate variability on the circum-Mediterranean region. Geology 2010, 38, 715–718. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Upper Badenian | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lithostratigraphy | Machów Formation | ||||||||||||
| Pecten Beds | Syndesmya Beds | ||||||||||||
| Sample | 8 | 9 | 11 | 13 | 15 | 17 | 19 | 21 | 23 | 25 | 27 | 28 | |
| Dinoflagellate Cysts | |||||||||||||
| 1. | Pentadinium laticinctum | 13 | 1 | 2 | 2 | 1 | 4 | 1 | |||||
| 2. | Batiacasphaera sphaerica | 68 | 3 | 3 | 4 | 5 | 3 | 1 | 20 | 22 | 3 | 2 | |
| 3. | Spiniferites ramosus s.l. | 81 | 162 | 135 | 120 | 117 | 5 | 138 | 2 | 153 | 65 | 109 | 128 |
| 4. | Batiacasphaera sp. A | 79 | 12 | 87 | 43 | 41 | 44 | 32 | 16 | 51 | 1 | ||
| 5. | Operculodinium centrocarpum | 27 | 51 | 5 | 59 | 96 | 5 | 37 | 9 | 18 | 3 | 25 | 29 |
| 6. | Melitasphaeridium ?pseudorecurvatum | 3 | |||||||||||
| 7. | Homotryblium tenuispinosum * | * | * | * | |||||||||
| 8. | Melitasphaeridium choanophorum | 4 | 15 | 3 | 13 | 4 | 6 | 3 | 12 | ||||
| 9. | Pyxidinopsis psilata | 7 | 3 | 1 | 1 | 2 | 1 | 12 | |||||
| 10. | Systematophora placacantha | 6 | 41 | 16 | 10 | 12 | 5 | 2 | 1 | 7 | 3 | ||
| 11. | Lingulodinium machaerophorum | 9 | 5 | 11 | 14 | 2 | 15 | 12 | 3 | 5 | |||
| 12. | Labyrinthodinium truncatum | 5 | 10 | 12 | 7 | 2 | 2 | 2 | |||||
| 13. | Reticulatosphaera actinocoronata | 1 | 3 | 5 | 1 | 6 | 18 | 6 | 2 | ||||
| 14. | Dapsilidinium sp. | 2 | |||||||||||
| 15. | Operculodinium sp. C | 2 | |||||||||||
| 16. | Spiniferites pseudofurcatus | 1 | 2 | 1 | 3 | 5 | |||||||
| 17. | Hystrichostrogylon sp. | 1 | |||||||||||
| 18. | Reticulatosphaera? sp. | 1 | |||||||||||
| 19. | Spiniferites sp. A | 3 | 4 | 4 | 2 | 2 | 3 | 3 | 6 | ||||
| 20. | Lejeunecysta sp. | 1 | 1 | 2 | |||||||||
| 21. | Hystrichosphaeropsis obscura | 1 | 4 | ||||||||||
| 22. | Pyxidinopsis? sp. | 1 | 1 | ||||||||||
| 23. | Cordosphaeridium cf. minimum | 2 | 1 | 8 | 1 | ||||||||
| 24. | Melitasphaeridium pseudorecurvatum | 1 | 1 | 1 | 1 | 1 | 4 | 1 | |||||
| 25. | Nematosphaeropsis labyrinthus | 6 | 1 | 15 | 1 | 119 | |||||||
| 26. | Cerodinium sp. * | * | * | ||||||||||
| 27. | Homotryblium plectilum * | * | * | * | |||||||||
| 28. | Impagidinium sp. | 1 | 1 | 6 | 5 | 1 | |||||||
| 29. | Hystrichokolpoma rigaudiae | 2 | 1 | 1 | 2 | ||||||||
| 30. | Cordosphaeridium minimum | 1 | 1 | 1 | 3 | ||||||||
| 31. | Operculodinium sp. B | 4 | 2 | ||||||||||
| 32. | Wetzeliella sp. * | * | * | ||||||||||
| 33. | Charlesdowniea sp. * | * | |||||||||||
| 34. | Operculodinium sp. A | 1 | 2 | 1 | |||||||||
| 35. | Areosphaeridium sp. * | * | |||||||||||
| 36. | Glaphyrocysta semitecta * | * | |||||||||||
| 37. | Areosphaeridium diktyoplokum * | * | * | * | |||||||||
| 38. | Deflandrea phosphoritica * | * | * | ||||||||||
| 39. | Lingulodinium sp. A | 1 | |||||||||||
| 40. | Cleistosphaeridium sp. | 3 | |||||||||||
| 41. | Areoligera sp. * | * | * | ||||||||||
| Age | Lower Sarmatian | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lithostratigraphy | Machów Formation | |||||||||||||
| Syndesma Beds | Krakowiec Clays | |||||||||||||
| Sample | 29 | 31 | 33 | 35 | 37 | 39 | 41 | 43 | 47 | 49 | 51 | 53 | 55 | |
| Dinoflagellate Cysts | ||||||||||||||
| 1. | Pentadinium laticinctum | 3 | 1 | 2 | 13 | 1 | ||||||||
| 2. | Batiacasphaera sphaerica | 9 | 1 | |||||||||||
| 3. | Spiniferites ramosus s.l. | 12 | 242 | 135 | 3 | 25 | 24 | 52 | 141 | 12 | 31 | 50 | ||
| 4. | Batiacasphaera sp. A | 1 | 127 | |||||||||||
| 5. | Operculodinium centrocarpum | 8 | 10 | 2 | 22 | 3 | ||||||||
| 6. | Melitasphaeridium ?pseudorecurvatum | |||||||||||||
| 7. | Homotryblium tenuispinosum * | |||||||||||||
| 8. | Melitasphaeridium choanophorum | 2 | ||||||||||||
| 9. | Pyxidinopsis psilata | |||||||||||||
| 10. | Systematophora placacantha | 4 | 24 | 3 | 71 | 72 | 3 | 78 | 31 | |||||
| 11. | Lingulodinium machaerophorum | 13 | 15 | 27 | 13 | |||||||||
| 12. | Labyrinthodinium truncatum | 1 | ||||||||||||
| 13. | Reticulatosphaera actinocoronata | 1 | 25 | |||||||||||
| 14. | Dapsilidinium sp. | 2 | 1 | |||||||||||
| 15. | Operculodinium sp. C | |||||||||||||
| 16. | Spiniferites pseudofurcatus | 1 | ||||||||||||
| 17. | Hystrichostrogylon sp. | |||||||||||||
| 18. | Reticulatosphaera? sp. | |||||||||||||
| 19. | Spiniferites sp. A | 6 | 1 | |||||||||||
| 20. | Lejeunecysta sp. | 3 | 1 | |||||||||||
| 21. | Hystrichosphaeropsis obscura | |||||||||||||
| 22. | Pyxidinopsis? sp. | |||||||||||||
| 23. | Cordosphaeridium cf. minimum | 1 | 1 | 5 | 12 | 9 | ||||||||
| 24. | Melitasphaeridium pseudorecurvatum | 1 | ||||||||||||
| 25. | Nematosphaeropsis labyrinthus | |||||||||||||
| 26. | Cerodinium sp. * | |||||||||||||
| 27. | Homotryblium plectilum * | |||||||||||||
| 28. | Impagidinium sp. | 1 | ||||||||||||
| 29. | Hystrichokolpoma rigaudiae | |||||||||||||
| 30. | Cordosphaeridium minimum | 2 | 3 | 6 | ||||||||||
| 31. | Operculodinium sp. B | |||||||||||||
| 32. | Wetzeliella sp. * | |||||||||||||
| 33. | Charlesdowniea sp. * | |||||||||||||
| 34. | Operculodinium sp. A | |||||||||||||
| 35. | Areosphaeridium sp. * | |||||||||||||
| 36. | Glaphyrocysta semitecta * | |||||||||||||
| 37. | Areosphaeridium diktyoplokum * | |||||||||||||
| 38. | Deflandrea phosphoritica * | |||||||||||||
| 39. | Lingulodinium sp. A | |||||||||||||
| 40. | Cleistosphaeridium sp. | |||||||||||||
| 41. | Areoligera sp. * | |||||||||||||
| 42. | Polysphaeridium subtile | 16 | 39 | |||||||||||
| Age | Upper Badenian | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lithostratigraphy | Machów Formation | ||||||||||||
| Pecten Beds | Syndesmya Beds | ||||||||||||
| Sample | 8 | 9 | 11 | 13 | 15 | 17 | 19 | 21 | 23 | 25 | 27 | 28 | |
| Dinoflagellate Cysts | |||||||||||||
| 42. | Polysphaeridium subtile | 19 | 1 | 5 | 5 | ||||||||
| 43. | Cordosphaeridium sp. * | * | * | ||||||||||
| 44. | Batiacasphaera micropapillata | 6 | |||||||||||
| 45. | Polysphaeridium zoharyi | 1 | 5 | ||||||||||
| 56. | Batiacasphaera hirsuta | 13 | 95 | 10 | 39 | ||||||||
| 47. | Membranophoridium aspinatum * | * | |||||||||||
| 48. | Impagidinium pallidum | 1 | 3 | 2 | |||||||||
| 49. | Areosphaeridium michoudii * | * | |||||||||||
| 50. | Operculodinium? sp. | 1 | 5 | 2 | |||||||||
| 51. | Palaeohystrichophora? sp. * | * | |||||||||||
| 52. | Cleistosphaeridium sp. A * | * | |||||||||||
| 53. | Operculodinium sp. D | 3 | |||||||||||
| 54. | Operculodinium sp. E | 1 | |||||||||||
| 55. | Operculodinium sp. F | 7 | |||||||||||
| 56. | Hystrichokolpoma sp. | 3 | |||||||||||
| 57. | Achomosphaera sp. | 21 | |||||||||||
| 58. | Pentadinium sp. A | ||||||||||||
| 59. | Selenopemphix brevispinosa? | ||||||||||||
| 60. | Hystrichosphaeropsis sp. | ||||||||||||
| 61. | Selenopemphix nephroides | ||||||||||||
| 62. | Systematophora ?ancyrea | ||||||||||||
| 63. | Operculodinium? sp. | ||||||||||||
| 64. | Surculosphaeridium longifurcatum * | ||||||||||||
| 65. | Phthanoperidinium comatum * | ||||||||||||
| 66. | Isabelidinium sp.* | ||||||||||||
| Total Counts | 305 | 298 | 321 | 266 | 313 | 60 | 289 | 54 | 319 | 185 | 314 | 276 | |
| Age | Lower Sarmatian | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lithostratigraphy | Machów Formation | |||||||||||||
| Syndesma Beds | Krakowiec Clays | |||||||||||||
| Sample | 29 | 31 | 33 | 35 | 37 | 39 | 41 | 43 | 47 | 49 | 51 | 53 | 55 | |
| Dinoflagellate Cysts | ||||||||||||||
| 43. | Cordosphaeridium sp. * | |||||||||||||
| 44. | Batiacasphaera micropapillata | |||||||||||||
| 45. | Polysphaeridium zoharyi | 3 | ||||||||||||
| 56. | Batiacasphaera hirsuta | 1 | ||||||||||||
| 47. | Membranophoridium aspinatum * | |||||||||||||
| 48. | Impagidinium pallidum | |||||||||||||
| 49. | Areosphaeridium michoudii * | |||||||||||||
| 50. | Operculodinium? sp. | |||||||||||||
| 51. | Palaeohystrichophora? sp. * | |||||||||||||
| 52. | Cleistosphaeridium sp. A * | |||||||||||||
| 53. | Operculodinium sp. D | |||||||||||||
| 54. | Operculodinium sp. E | |||||||||||||
| 55. | Operculodinium sp. F | |||||||||||||
| 56. | Hystrichokolpoma sp. | 1 | 2 | |||||||||||
| 57. | Achomosphaera sp. | |||||||||||||
| 58. | Pentadinium sp. A | 8 | 1 | |||||||||||
| 59. | Selenopemphix brevispinosa? | 2 | 2 | 3 | 1 | |||||||||
| 60. | Hystrichosphaeropsis sp. | 1 | 1 | |||||||||||
| 61. | Selenopemphix nephroides | 42 | 3 | 3 | 1 | 4 | 2 | |||||||
| 62. | Systematophora ?ancyrea | 65 | 36 | |||||||||||
| 63. | Operculodinium? sp. | 18 | 21 | 28 | 4 | 10 | ||||||||
| 64. | Surculosphaeridium longifurcatum * | * | ||||||||||||
| 65. | Phthanoperidinium comatum * | * | ||||||||||||
| 66. | Isabelidinium sp. * | * | ||||||||||||
| Total Counts | 0 | 0 | 36 | 313 | 236 | 35 | 159 | 84 | 155 | 274 | 161 | 131 | 103 | |
| Fossil taxon | Botanical affinity | Element |
| Spores of plants | ||
| Baculatisporites sp. | Osmundaceae: Osmunda | P/A |
| Camarozonosporites sp. | Lycopodiaceae: Lycopodiella sect. Campylostachys | P |
| Distverrusporis sp. | Sphagnaceae: Sphagnum | P/A |
| Laevigatosporites sp. | Polypodiaceae, Davalliaceae, and other ferns | P/A |
| Leiotriletes sp. | Lygodiaceae and other ferns | P |
| Neogenisporis sp. | Gleicheniaceae and Cyatheaceae | P1 |
| Retitriletes sp. | Lycopodiaceae: Lycopodium | A |
| Stereisporites sp. | Sphagnaceae: Sphagnum | P/A |
| Verrucatosporites sp. | Davalliaceae, Polypodiaceae, and other ferns | P/A |
| Pollen grains of gymnosperms | ||
| Abiespollenites sp. | Pinaceae: Abies | A |
| Cathayapollis sp. | Pinaceae: Cathaya | A1 |
| Cedripites sp. | Pinaceae: Cedrus | A1 |
| Inaperturopollenites concedipites (Wodehouse and Krutzsch) | Cupressaceae: Taxodium and Glyptostrobus | P2/A1 |
| Inaperturopollenites verrupapillatus Trevisan | Cupressaceae: Taxodium and Glyptostrobus | P2/A1 |
| Keteleeriapollenites sp. | Pinaceae: Keteleeria and Pseudolarix | A1 |
| Piceapollis praemarianus Krutzsch | Pinaceae: Picea | A |
| Piceapollis sp. | Pinaceae: Picea | A |
| Pinuspollenites labdacus (Potonié) Raatz | Pinaceae: Pinus sylvestris type | A |
| Pinuspollenites sp. | Pinaceae: Pinus | A |
| Sciadopityspollenites verticillatiformis (Zauer) Krutzsch | Sciadopityaceae: Sciadopitys | A1 |
| Sciadopityspollenites sp. | Sciadopityaceae: Sciadopitys | A1 |
| Sequoiapollenites sp. | Cupressaceae: Sequoia, Sequoiadendron, and Metasequoia | A1 |
| Zonalapollenites gracilis Krutzsch | Pinaceae: Tsuga | A |
| Zonalapollenites verrucatus Krutzsch | Pinaceae: Tsuga | A |
| Zonalapollenites sp. | Pinaceae: Tsuga | A |
| Pollen grains of angiosperms | ||
| Aceripollenites sp. | Sapindaceae: Acer | A1 |
| Alnipollenites verus Potonié | Betulaceae: Alnus | P2/A |
| Caprifoliipites sp. | Adoxaceae: Sambucus and Viburnum | P/A1 |
| Carpinipites carpinoides (Pflug) Nagy | Betulaceae: Carpinus | P2/A1 |
| Caryapollenites simplex (Potonié) Raatz | Juglandaceae: Carya | A1 |
| Celtipollenites sp. | Ulmaceae: Celtis | P/A1 |
| Cercidiphyllites minimireticulatus (Trevisan) Ziembińska-Tworzydło | Cercidiphyllaceae: Cercidiphyllum | A1 |
| Chenopodipollis sp. | Amaranthaceae (incl. Chenopodiaceae) | P/A |
| Cupuliferoipollenites oviformis (Potonié) Potonié | Fagaceae: Castanea, Castanopsis, and Lithocarpus | P2/A1 |
| Cupuliferoipollenites pusillus (Potonié) Potonié | Fagaceae: Castanea, Castanopsis, and Lithocarpus | P2/A1 |
| Cyperaceaepollis neogenicus Krutzsch | Cyperaceae | P/A |
| Cyrillaceaepollenites megaexactus (Potonié) Potonié | Cyrillaceae and Clethraceae | P |
| Edmundipollis sp. | Araliaceae, Cornaceae, and Mastixiaceae | P/A |
| Ericipites baculatus Nagy | Ericaceae | A |
| Ericipites ericius (Potonié) Potonié | Ericaceae | A |
| Ericipites sp. | Ericaceae | A |
| Eucommiapollis minor Menke | Eucommiaceae: Eucommia | A1 |
| Faguspollenites cf. verus Raatz | Fagaceae: Fagus | A |
| Faguspollenites sp. | Fagaceae: Fagus | A |
| Fraxinipollis oblatus Słodkowska | Oleaceae: Fraxinus | P/A |
| Graminidites sp. | Poaceae: Pooideae | P/A |
| Ilexpollenites iliacus (Potonié) Thiergart | Aquifoliaceae: Ilex | P/A1 |
| Ilexpollenites margaritatus (Potonié) Thiergart | Aquifoliaceae: Ilex | P2 |
| Intratriporopollenites sp. | Malvaceae: Tilioideae | P/A1 |
| Juglanspollenites sp. | Juglandaceae: Juglans | A1/P2 |
| Myricipites sp. | Myricaceae: Myrica | P2/A1 |
| Nyssapollenites sp. | Nyssaceae: Nyssa | P2/A1 |
| Parthenopollenites marcodurensis (Pflug and Thomson) Traverse | Vitaceae | P/A1 |
| Periporopollenites orientaliformis (Nagy) Kohlman-Adamska and Ziembińska-Tworzydło | Altingiaceae: Liquidambar | A1 |
| Periporopollenites stigmosus (Potonié) Thomson and Pflug) | Altingiaceae: Liquidambar | A1 |
| Polyatriopollenites stellatus (Potonié) Pflug | Juglandaceae: Pterocarya | A1 |
| Quercoidites henricii (Potonié) Potonié, Thomson, and Thiergart) | Fagaceae: Quercus | P2/A1 |
| Quercopollenites rubroides Kohlman-Adamska and Ziembińska-Tworzydło | Fagaceae: Quercus | A1 |
| Quercopollenites sp. | Fagaceae: Quercus | A1 |
| Salixipollenites capreaformis Planderová | Salicaceae: Salix | A |
| Salixipollenites sp. | Salicaceae: Salix | A |
| Sparganiaceaepollenites sp. | Sparganiaceae and Typhaceae | P/A |
| Symplocoipollenites vestibulum (Potonié) Potonié | Symplocaceae: Symplocos | P |
| Tricolporopollenites fallax (Potonié) Krutzsch | Fabaceae | P/A |
| Tricolporopollenites indeterminatus (Romanowicz) Ziembińska-Tworzydło | Hamamelidaceae | P2 |
| Tricolporopollenites liblarensis (Thomson) Hochuli | Fabaceae | P/A |
| Tricolporopollenites pseudocingulum (Potonié) Thomson and Pflug | Fagaceae? and Styracaceae? | P/A1 |
| Trivestibulopollenites betuloides Pflug | Betulaceae: Betula | A |
| Ulmipollenites undulosus Wolff | Ulmaceae: Ulmus | A2 |
| Ulmipollenites sp. | Ulmaceae: Ulmus | A2 |
| Zelkovaepollenites sp. | Ulmaceae: Zelkova | A1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peryt, D.; Gedl, P.; Worobiec, E.; Worobiec, G.; Peryt, T.M. Foraminiferal and Palynological Records of an Abrupt Environmental Change at the Badenian/Sarmatian Boundary (Middle Miocene): A Case Study in Northeastern Central Paratethys. Geosciences 2024, 14, 86. https://doi.org/10.3390/geosciences14030086
Peryt D, Gedl P, Worobiec E, Worobiec G, Peryt TM. Foraminiferal and Palynological Records of an Abrupt Environmental Change at the Badenian/Sarmatian Boundary (Middle Miocene): A Case Study in Northeastern Central Paratethys. Geosciences. 2024; 14(3):86. https://doi.org/10.3390/geosciences14030086
Chicago/Turabian StylePeryt, Danuta, Przemysław Gedl, Elżbieta Worobiec, Grzegorz Worobiec, and Tadeusz Marek Peryt. 2024. "Foraminiferal and Palynological Records of an Abrupt Environmental Change at the Badenian/Sarmatian Boundary (Middle Miocene): A Case Study in Northeastern Central Paratethys" Geosciences 14, no. 3: 86. https://doi.org/10.3390/geosciences14030086