OK-432 Administration Inhibits Murine Allergic Rhinitis at the Induction Phase, through the Macrophage Activation with TLR2 Signaling Pathway



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Reagents and Antibodies
2.3. Measurement of Cytokine Production by Spleen Cells
2.4. Measurement of Cytokine Production by Peritoneal Macrophages
2.5. Immunization Protocol and Treatment
2.6. Measurement of Cytokine Production by CD3+ Splenic T Cells
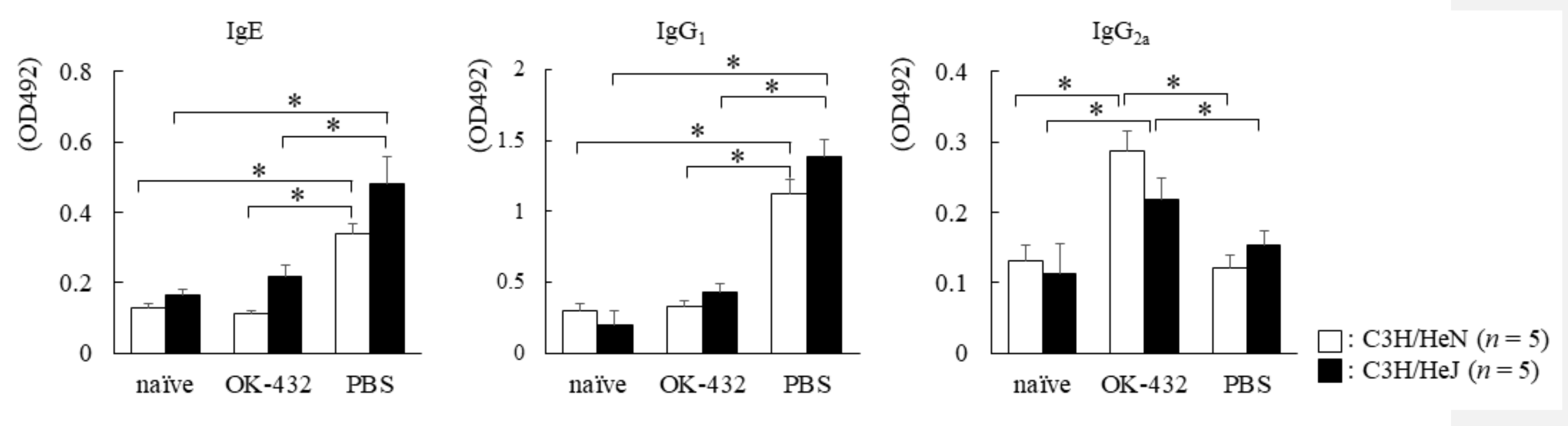
2.7. Measurement of OVA-Specific IgE, IgG1 and IgG2a
2.8. Evaluation of Nasal Symptom
2.9. Histological Examination
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
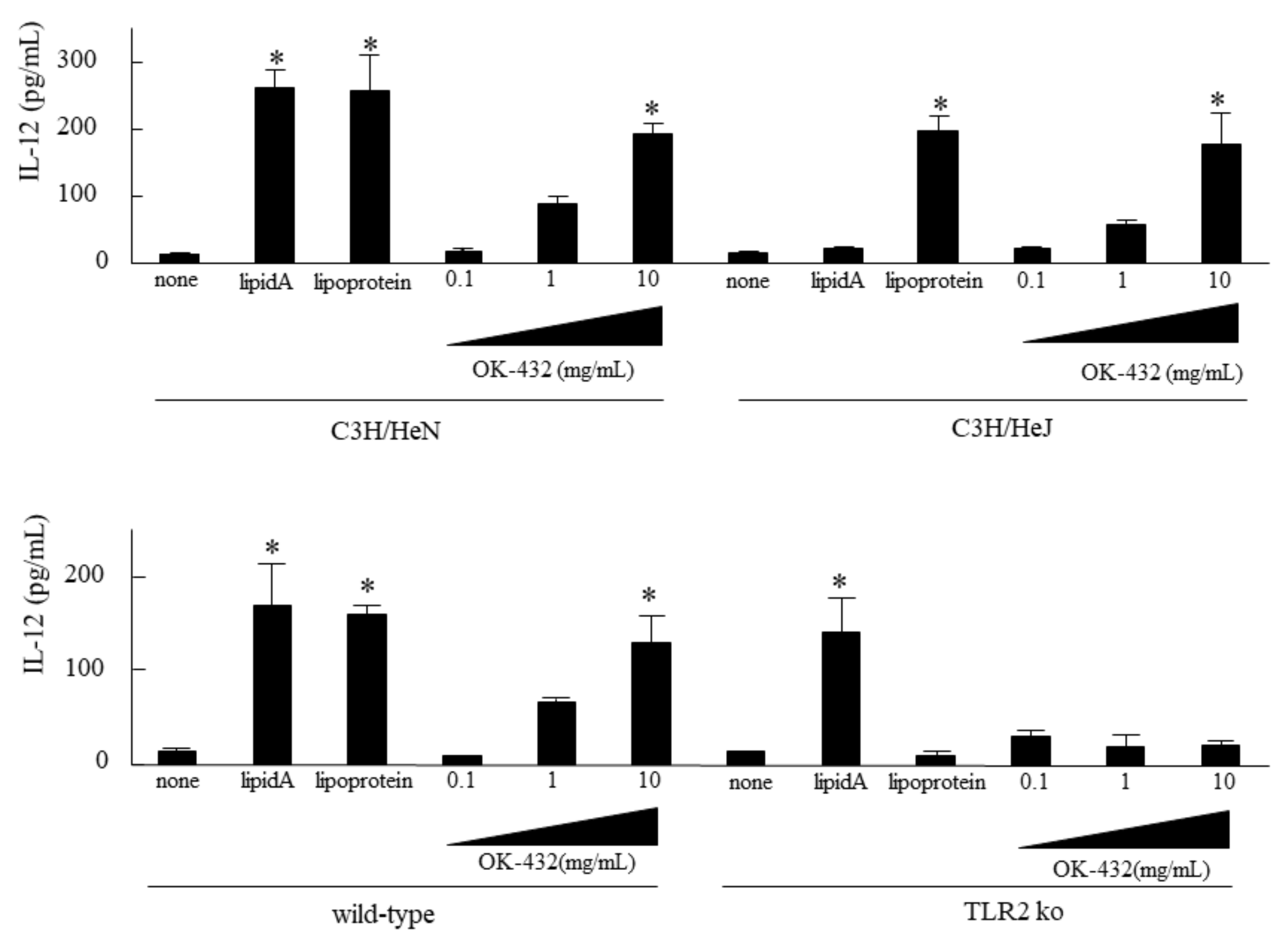
3.1. Effect of OK-432 on IL-12 Production by Splenic Macrophages Derived from C3H/HeN, C3H/HeJ, TLR2 KO and C57/BL6 Mice In Vitro
3.2. Effect of OK-432 on IL-12 Production by Murine Peritoneal Macrophages In Vivo
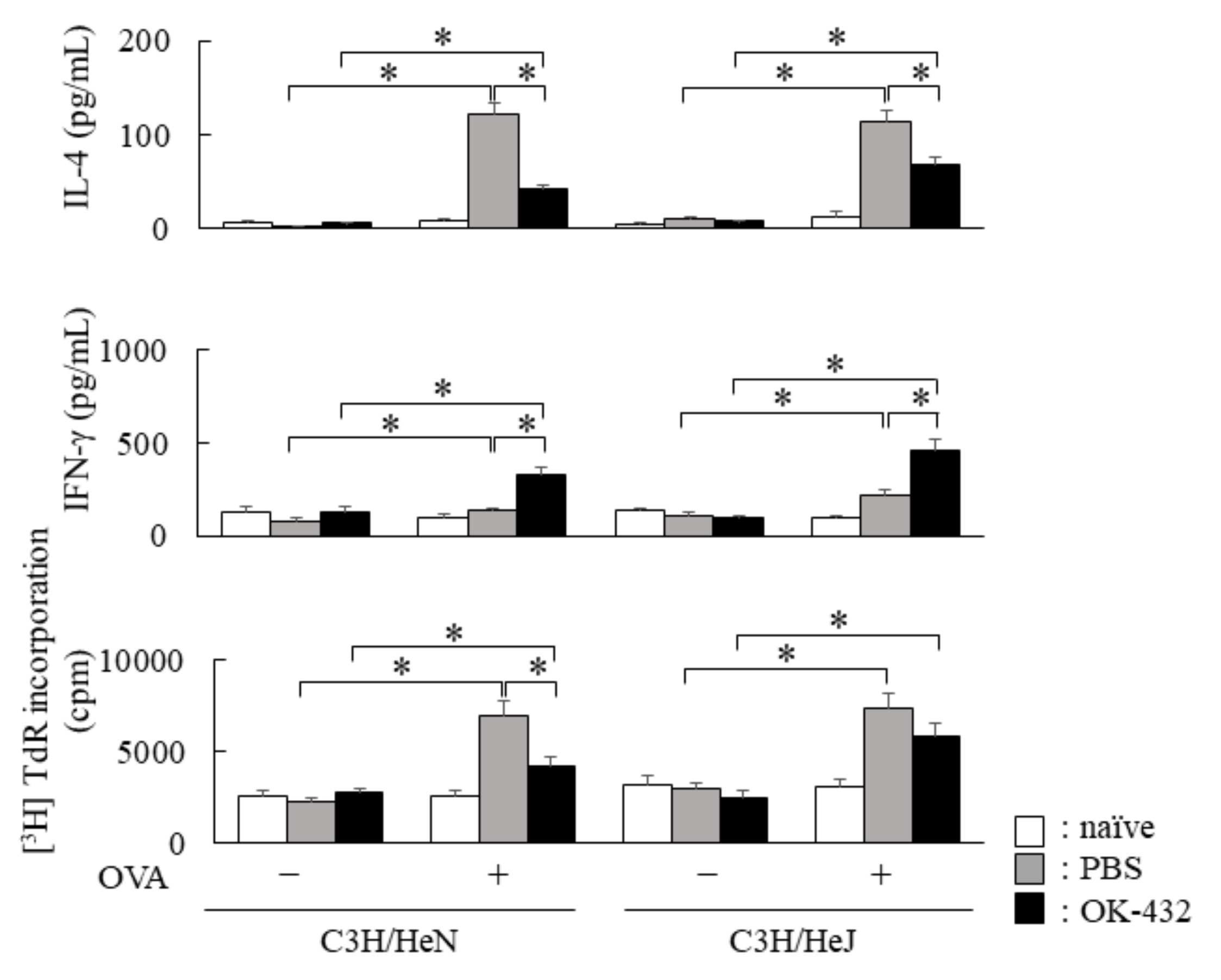
3.3. The Effect of OK-432 for Th2 Responses and Allergic Rhinitis in C3H/HeN and C3H/HeJ Mice Sensitized with OVA at Induction Phase
3.4. The Effect of OK-432 for Th2 Responses and Allergic Rhinitis in C57/BL6 and TLR2 Knock-Out Mice Sensitized with OVA at Induction Phase
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baraniuk, J.N. Mechanisms of allergic rhinitis. Curr. Allergy Asthma Rep. 2001, 1, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D.; HayGlass, K.T. T cell chemokine receptor expression in human Th1- and Th2-associated diseases. Arch. Immunol. Ther. Exp. 2000, 48, 451–456. [Google Scholar]
- Pearlman, D.S. Pathophysiology of the inflammatory response. J. Allergy Clin. Immunol. 1999, 104 Pt 1, S132–S137. [Google Scholar] [CrossRef]
- Durham, S.R. Mechanisms of mucosal inflammation in the nose and lungs. Clin Exp Allergy 1998, 28, 11–16. [Google Scholar] [PubMed]
- Baraniuk, J.N. Pathogenesis of allergic rhinitis. J. Allergy Clin. Immunol. 1997, 99, S763–S772. [Google Scholar] [CrossRef]
- Punnonen, J.; Aversa, G.; Cocks, B.G.; de Vries, J.E. Role of interleukin-4 and interleukin-13 in synthesis of IgE and expression of CD23 by human B cells. Allergy 1994, 49, 576–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, J.E.; Punnonen, J.; Cocks, B.G.; de Waal Malefyt, R.; Aversa, G. Regulation of the human IgE response by IL4 and IL13. Res. Immunol. 1993, 144, 597–601. [Google Scholar] [CrossRef]
- Fujieda, S.; Zhang, K.; Saxon, A. IL-4 plus CD40 monoclonal antibody induces human B cells gamma subclass-specific isotype switch: Switching to gamma 1, gamma 3, and gamma 4, but not gamma 2. J. Immunol. 1995, 155, 2318–2328. [Google Scholar]
- Gajewski, T.F.; Joyce, J.; Fitch, F.W. Antiproliferative effect of IFN-gamma in immune regulation: III. Differential selection of Th1 and Th2 murine helper T lymphocyte clones using recombinant IL-2 and recombinant IFN-gamma. J. Immunol. 1989, 143, 15. [Google Scholar]
- Manetti, R.; Parronchi, P.; Giudizi, M.G.; Piccini, M.P.; Maggi, E.; Trinchieri, G.; Romagnani, S. Natural killer cell stimulatory factor (interleukin 12 [IL-12]) induces T helper type 1 (Th1)-specific immune responses and inhibits the development of IL-4-producing Th cells. J. Exp. Med. 1993, 177, 1199–1204. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1993, 7, 145. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G.; Pflanz, S.; Kastelein, R.A. The IL-12 family of heterodimeric cytokines: New players in the regulation of T cell responses. Immunity 2003, 19, 641–644. [Google Scholar] [CrossRef]
- Trinchieri, G. Interleukin-12: A proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. Annu Rev Immunol. 1995, 13, 251–276. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12: A cytokine produced by antigen-presenting cells with immunoregulatory functions in the generation of T-helper cells type 1 and cytotoxic lymphocytes. Blood 1994, 84, 4008–4027. [Google Scholar]
- Wolf, S.F.; Sieburth, D.; Sypek, J. Interleukin 12: A key modulator of immune function. Stem Cells 1994, 12, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.Y.; Liao, T.N.; Chiang, B.L.; Huang, M.S.; Chen, C.C.; Chou, C.C.; Hsieh, K.H. Decreased production of IFN gamma and increased production of IL-6 by cord blood mononuclear cells of newborns with a high risk of allergy. Clin. Exp. Allergy 1996, 26, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Borres, M.P.; Einarsson, R.; Bjorksten, B. Serum levels of interleukin-4, soluble CD23 and IFN gamma in relation to the development of allergic disease during the first 18 months of life. Clin. Exp. Allergy 1995, 25, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.L.; Kemp, A.; Thorburn, S.J.; Hill, D.J. Reduced interferon-gamma secretion in neonates and subsequent atopy. Lancet 1994, 344, 983–985. [Google Scholar] [CrossRef]
- Okamoto, H.; Shoin, S.; Koshimura, S.; Shimizu, R. Studies on the anticancer and streptolysin S-forming abilities of hemolytic streptococci. Jpn. J. Microbiol. 1967, 11, 323–336. [Google Scholar] [CrossRef]
- Oshimi, K.; Kano, S.; Takaku, F.; Okumura, K. Augmentation of mouse natural killer cell activity by a streptococcal preparation, OK-432. J. Natl. Cancer Inst. 1980, 65, 1265–1269. [Google Scholar]
- Saito, M.; Ebina, T.; Koi, M.; Yamaguchi, T.; Kamade, Y.; Ishida, N. Induction of interferon-γ in mouse spleen cells by OK-432, a preparation of Streptococcus pyogenes. Cell Immunol. 1982, 68, 187–192. [Google Scholar] [CrossRef]
- Ichimura, O.; Suzuki, S.; Saito, M.; Sugawara, Y.; Ishida, N. Augmentation of interleukin 1 and interleukin 2 production by OK-432. Int. J. Immunopharmacol. 1985, 7, 263–270. [Google Scholar] [CrossRef]
- Fukui, H.; Koshihara, Y.; Nagamuta, M.; Mizutani, Y.; Uchida, A. Production of interleukin 6 by human spleen cells stimulated with streptococcal preparation OK-432. Immunol. Lett. 1989, 21, 127–130. [Google Scholar] [CrossRef]
- Misaki, T.; Watanabe, Y.; Iida, Y.; Hidaka, A.; Kasagi, K.; Fukushima, H.; Konishi, J. Recruitment of T lymphocytes and induction of tumor necrosis factor in thyroid cancer by a local immunotherapy. Cancer Immunol. Immunother. 1992, 35, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Duda, R.B.; Azilvasi, A.; Chen, X.; Mai, M.; O’Donnell, M.A. Streptococcal preparation OK-432 is a potent inducer of IL-12 and a T helper cell 1 dominant state. J. Immunol. 1997, 158, 5619–5626. [Google Scholar] [PubMed]
- Okamoto, M.; Ohe, G.; Oshikawa, T.; Nishikawa, H.; Furuichi, S.; Yoshida, H.; Matsuno, T.; Saito, M.; Sato, M. Induction of Th1-type cytokines by lipoteichoic acid-related preparation isolated from OK-432, a penicillin-killed streptococcal agent. Int. Immunopharmacol. 2000, 49, 363–376. [Google Scholar] [CrossRef]
- Zhang, G.; Ghosh, S. Toll-like receptor-mediated NF-κB activation: A phylogenetically conserved paradigm in innate immunity. J. Clin. Investig. 2001, 10, 13–19. [Google Scholar] [CrossRef]
- Takeuchi, O.; Hoshino, K.; Kawai, T.; Sanjo, H.; Takada, H.; Ogawa, T.; Takeda, K.; Akira, S. Differential Roles of TLR2 and TLR4 in Recognition of Gram-Negative and Gram-Positive Bacterial Cell Wall Components. Immunity 1999, 11, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Ishimitsu, R.; Nishimura, H.; Yajima, T.; Watase, T.; Kawauchi, H.; Yoshikai, Y. Overexpression of IL-15 in vivo enhances Tc1 response, which inhibits allergic inflammation in a murine model of asthma. J. Immunol. 2001, 166, 1991–2001. [Google Scholar] [CrossRef]
- Ishimitsu, R.; Yajima, T.; Nishimura, H.; Kawauchi, H.; Yoshikai, Y. NKT cells are dispensable in the induction of oral tolerance but are indispensable in the abrogation of oral tolerance by prostaglandin E. Eur. J. Immunol. 2003, 33, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Onozaki, K.; Suzuki, K.; Fukutomi, Y.; Yagami, K.; Hashimoto, T. Activation and augmentation of guinea pig macrophages with streptococcal preparation OK-432 and stimulated spleen cell products. J. Reticuloendothel. Soc. 1982, 32, 433–442. [Google Scholar]
- Umemura, M.; Nishimura, H.; Hirose, K.; Matsuguchi, T.; Yoshikai, Y. Overexpression of IL-15 in vivo enhances protection against Mycobacterium bovis bacillus Calmette-Guerin infection via augmentation of NK and T cytotoxic 1 responses. J. Immunol. 2001, 167, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Kawamoto, E.; Chen, Z.; Kamei, C. A new model of allergic rhinitis in rats by topical sensitization and evaluation of H1-receptor antagonists. Immunopharmacology 2000, 48, 1–7. [Google Scholar] [CrossRef]
- Usami, H.; Yamamoto, A.; Yamashita, W.; Sugawara, Y.; Hamada, S.; Yamamoto, T.; Kato, K.; Kokeguchi, S.; Ohokuni, H.; Kotani, S. Antitumour effects of streptococcal lipoteichoic acids on Meth A fibrosarcoma. Br. J. Cancer 1988, 57, 70–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seder, R.A.; Paul, W.E. Acquisition of lymphokine-producing phenotype by CD4+ T cells. Annu Rev Immunol. 1994, 12, 635–673. [Google Scholar] [CrossRef]
- Leonard, P.; Sur, S. Interleukin-12: Potential role in asthma therapy. BioDrugs 2003, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2000, 85, 9–18. [Google Scholar] [CrossRef]
- Strober, W.; Kelsall, B.; Fuss, I.; Marth, T.; Ludviksson, B.; Ehrhardt, R.; Neurath, M. Reciprocal IFN-γ and TGF-β responses regulate the occurrence of mucosal inflammation. Immunol Today 1997, 18, 61–64. [Google Scholar] [CrossRef]
- Biron, C.A.; Gazzinelli, R.T. Effects of IL-12 on immune responses to microbial infections: A key mediator in regulating disease outcome. Curr. Opin. Immunol. 1995, 7, 485–496. [Google Scholar] [CrossRef]
- O’Garra, A.; Murphy, K. Role of cytokines in determining T-lymphocyte function. Curr Opin Immunol. 1994, 6, 458–466. [Google Scholar] [CrossRef]
- Kira, J.; Kawano, Y.; Yamasaki, K. Multiple sclerosis with mite antigen-specific IgE. J. Neurol. Sci. 1998, 157, 138–142. [Google Scholar] [CrossRef]
- Allanore, Y.; Hilliquin, P.; Coste, J.; Renoux, M.; Menkes, C.J. Decreased prevalence of atopy in rheumatoid arthritis. Lancet 1998, 351, 497. [Google Scholar] [CrossRef]
- Alm, J.S.; Lilja, G.; Pershagen, G.; Scheynius, A. Early BCG vaccination and development of atopy. Lancet 1997, 350, 400–403. [Google Scholar] [CrossRef]
- Prokopakis, E.; Vardouniotis, A.; Kawauchi, H.; Scadding, G.; Georgalas, C.; Hellings, P.; Velegrakis, G.; Kalogjera, L. The pathophysiology of the hygiene hypothesis. Int. J. Pediatr. Otorhinolaryngol. 2013, 77, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Olsnes, C.; Olofsson, J.; Aarstad, H.J. MAPKs ERK and p38, but not JNK phosphorylation, modulate IL-6 and TNF-α secretion following OK-432 in vitro stimulation of purified human monocytes. Scand. J. Immunol. 2011, 74, 114–125. [Google Scholar] [CrossRef]
- Olsnes, C.; Bredholt, T.; Olofsson, J.; Aarstad, H.J. OK-432-stimulated chemokine secretion from human monocytes depends on MEK1/2, and involves p38 MAPK and NF-κB phosphorylation, in vitro. APMIS 2013, 121, 299–310. [Google Scholar] [CrossRef] [PubMed]












© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoi, N.; Morikura, I.; Fuchiwaki, T.; Yamada, T.; Prokopakis, E.; Kawauchi, H. OK-432 Administration Inhibits Murine Allergic Rhinitis at the Induction Phase, through the Macrophage Activation with TLR2 Signaling Pathway. Med. Sci. 2018, 6, 107. https://doi.org/10.3390/medsci6040107
Aoi N, Morikura I, Fuchiwaki T, Yamada T, Prokopakis E, Kawauchi H. OK-432 Administration Inhibits Murine Allergic Rhinitis at the Induction Phase, through the Macrophage Activation with TLR2 Signaling Pathway. Medical Sciences. 2018; 6(4):107. https://doi.org/10.3390/medsci6040107
Chicago/Turabian StyleAoi, Noriaki, Ichiro Morikura, Takafumi Fuchiwaki, Takaya Yamada, Emmanuel Prokopakis, and Hideyuki Kawauchi. 2018. "OK-432 Administration Inhibits Murine Allergic Rhinitis at the Induction Phase, through the Macrophage Activation with TLR2 Signaling Pathway" Medical Sciences 6, no. 4: 107. https://doi.org/10.3390/medsci6040107