Differential Expression of Tissue Transglutaminase Splice Variants in Peripheral Blood Mononuclear Cells of Primary Progressive Multiple Sclerosis Patients

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Isolation of Primary Human Peripheral Blood Mononuclear Cells
2.3. mRNA Isolation and cDNA Synthesis
2.4. Semi-Quantitative Real-Time PCR (qPCR)
2.5. Statistical Analysis
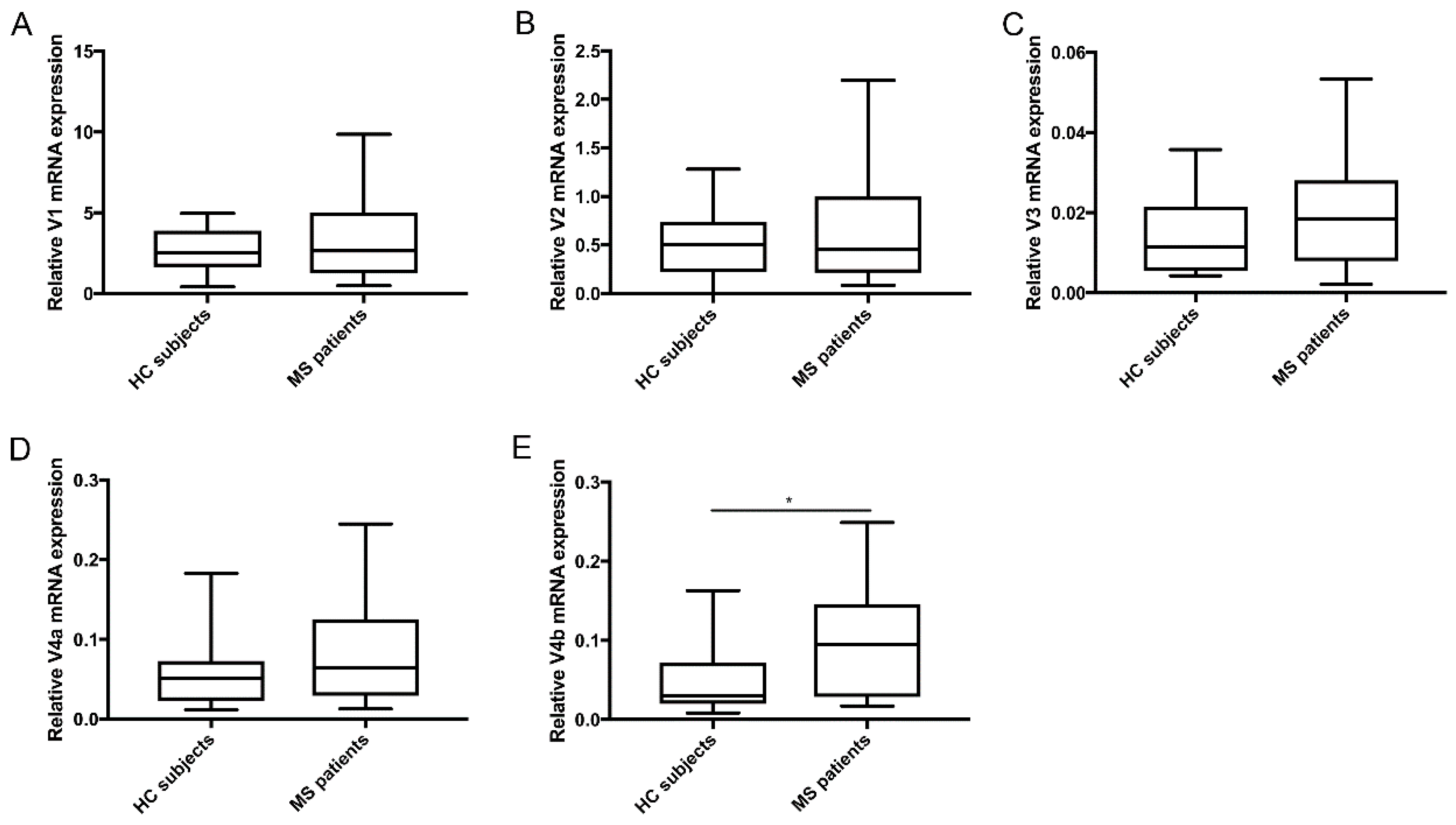
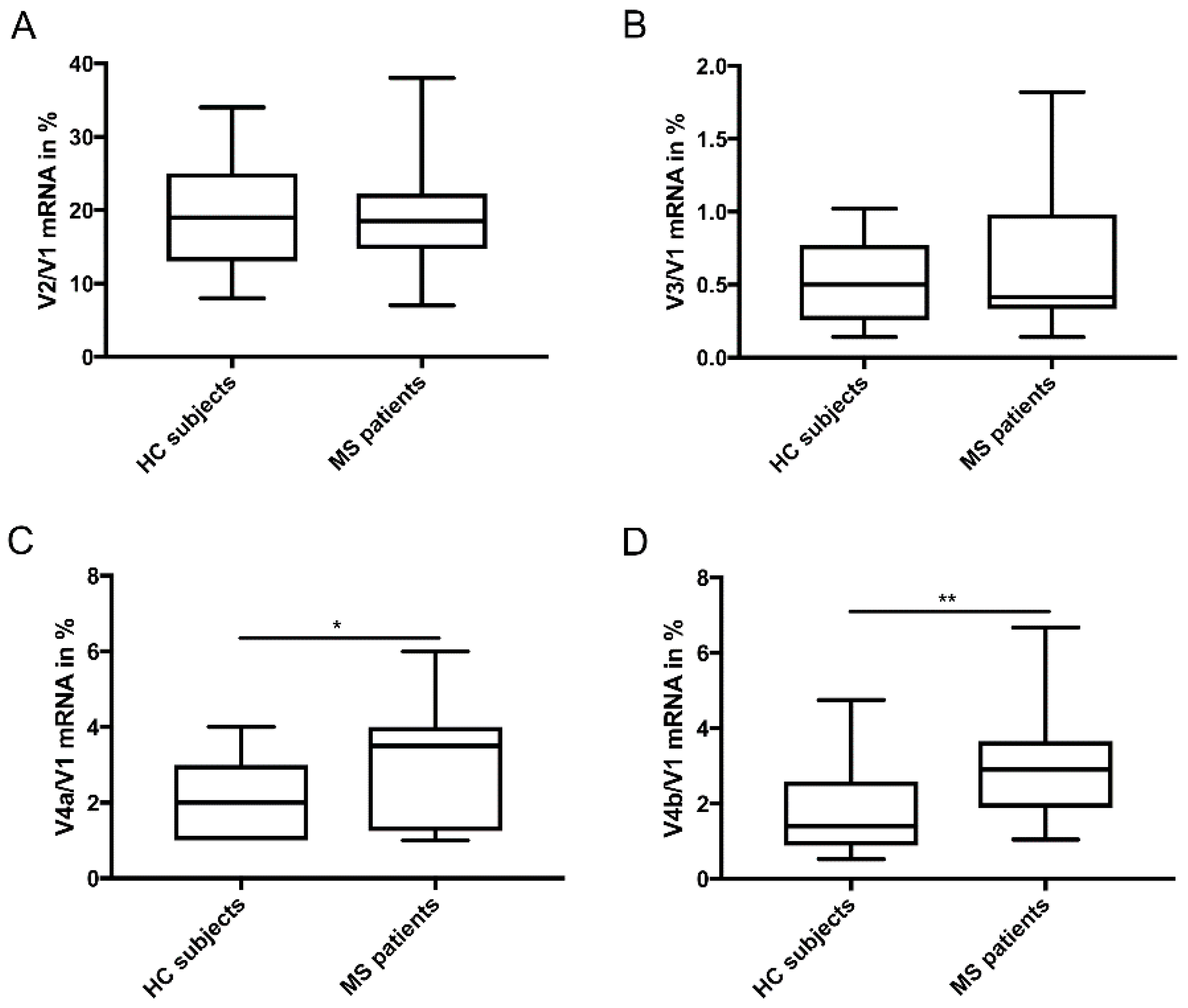
3. Results
Differential Expression of TG2 Splice Variants in PP-MS Patients Compared to HC Subjects
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Noseworthy, J.; Lucchinetti, C.; Rodriguez, M.; Weinshenker, B. Multiple sclerosis. N. Engl. J. Med. 2000, 343, 938–952. [Google Scholar] [CrossRef] [PubMed]
- Høglund, R.A.; Maghazachi, A.A. Multiple sclerosis and the role of immune cells. World J. Exp. Med. 2014, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fesus, L.; Piacentini, M. Transglutaminase 2: An enigmatic enzyme with diverse functions. Trends Biochem. Sci. 2002, 27, 534–539. [Google Scholar] [CrossRef]
- Bayardo, M.; Punzi, F.; Bondar, C.; Chopita, N.; Chirdo, F. Transglutaminase 2 expression is enhanced synergistically by interferon-γ and tumour necrosis factor-α in human small intestine. Clin. Exp. Immunol. 2012, 168, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.M.; Jeon, J.H.; Kim, C.W.; Cho, S.Y.; Lee, H.J.; Jang, G.Y.; Jeong, E.M.; Lee, D.S.; Kang, J.H.; Melino, G. TGFβ mediates activation of transglutaminase 2 in response to oxidative stress that leads to protein aggregation. FASEB J. 2008, 22, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, Y.; Hiraide, S.; Matsumoto, M.; Shimamura, K.; Togashi, H. Enhanced transglutaminase 2 expression in response to stress-related catecholamines in macrophages. Immunobiology 2014, 219, 680–686. [Google Scholar] [CrossRef]
- Phatak, V.; Croft, S.; Setty, S.R.; Scarpellini, A.; Hughes, D.; Rees, R.; McArdle, S.; Verderio, E. Expression of transglutaminase-2 isoforms in normal human tissues and cancer cell lines: Dysregulation of alternative splicing in cancer. Amino Acids 2013, 44, 33–44. [Google Scholar] [CrossRef]
- Lai, T.-S.; Greenberg, C.S. TGM2 and implications for human disease: Role of alternative splicing. Front. Biosci. (Landmark Ed.) 2013, 18, 504–519. [Google Scholar]
- Citron, B.A.; SantaCruz, K.S.; Davies, P.J.; Festoff, B.W. Intron-exon swapping of transglutaminase mRNA and neuronal Tau aggregation in Alzheimer’s disease. J. Biol. Chem. 2001, 276, 3295–3301. [Google Scholar] [CrossRef]
- Antonyak, M.A.; Jansen, J.M.; Miller, A.M.; Ly, T.K.; Endo, M.; Cerione, R.A. Two isoforms of tissue transglutaminase mediate opposing cellular fates. Proc. Natl. Acad. Sci. USA 2006, 103, 18609–18614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tee, A.E.; Marshall, G.M.; Liu, P.Y.; Xu, N.; Haber, M.; Norris, M.D.; Iismaa, S.E.; Liu, T. Opposing effects of two tissue transglutaminase protein isoforms in neuroblastoma cell differentiation. J. Biol. Chem. 2010, 285, 3561–3567. [Google Scholar] [CrossRef] [PubMed]
- Arbildi, P.; Sóñora, C.; Del Río, N.; Marqués, J.; Hernández, A. Alternative RNA splicing of leucocyte tissue transglutaminase in coeliac disease. Scand. J. Immunol. 2018, 87, e12659. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.-S.; Liu, Y.; Li, W.; Greenberg, C.S. Identification of two GTP-independent alternatively spliced forms of tissue transglutaminase in human leukocytes, vascular smooth muscle, and endothelial cells. FASEB J. 2007, 21, 4131–4143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestito, C.; Breve, J.J.P.; van Eggermond, M.; Killestein, J.; Teunissen, C.E.; van Rossum, J.; Wilhelmus, M.M.M.; Drukarch, B.; van den Elsen, P.J.; van Dam, A.M. Monocyte-derived tissue transglutaminase in multiple sclerosis patients: Reflecting an anti-inflammatory status and function of the cells? J. Neuroinflamm. 2017, 14, 257. [Google Scholar] [CrossRef]
- Sestito, C.; Steenwijk, M.; Leurs, C.; Breve, J.J.; Twisk, J.; Wilhelmus, M.M.; Drukarch, B.; Teunissen, C.E.; Killestein, J.; van Dam, A.M. Tissue transglutaminase expression in peripheral blood mononuclear cells correlates with disease progression and axonal damage in subtypes of MS patients: A cross-sectional study. Ann. Neurol. 2018. in preparation. [Google Scholar]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; Van den Hoff, M.J.B.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]
- Currò, M.; Ferlazzo, N.; Risitano, R.; Condello, S.; Vecchio, M.; Caccamo, D.; Ientile, R. Transglutaminase 2 and phospholipase A 2 interactions in the inflammatory response in human Thp-1 monocytes. Amino Acids 2014, 46, 759–766. [Google Scholar] [CrossRef]
- Kalyvas, A.; David, S. Cytosolic phospholipase A2 plays a key role in the pathogenesis of multiple sclerosis-like disease. Neuron 2004, 41, 323–335. [Google Scholar] [CrossRef]
- Wilhelmus, M.M.; de Jager, M.; Smit, A.B.; van der Loo, R.J.; Drukarch, B. Catalytically active tissue transglutaminase colocalises with Aβ pathology in Alzheimer’s disease mouse models. Sci. Rep. 2016, 6, 20569. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmus, M.M.; Grunberg, S.C.; Bol, J.G.; van Dam, A.M.; Hoozemans, J.J.; Rozemuller, A.J.; Drukarch, B. Transglutaminases and transglutaminase-catalyzed cross-links colocalize with the pathological lesions in Alzheimer’s disease brain. Brain Pathol. (Zurich Switz.) 2009, 19, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Ip, J.Y.; Tong, A.; Pan, Q.; Topp, J.D.; Blencowe, B.J.; Lynch, K.W. Global analysis of alternative splicing during T-cell activation. RNA 2007. [Google Scholar] [CrossRef] [PubMed]
- Evsyukova, I.; Somarelli, J.A.; Gregory, S.G.; Garcia-Blanco, M.A. Alternative splicing in multiple sclerosis and other autoimmune diseases. RNA Biol. 2010, 7, 462–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillett, A.; Maratou, K.; Fewings, C.; Harris, R.A.; Jagodic, M.; Aitman, T.; Olsson, T. Alternative splicing and transcriptome profiling of experimental autoimmune encephalomyelitis using genome-wide exon arrays. PLoS ONE 2009, 4, e7773. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subjects | Number | Female/Male | Age (years) ± SD | Disease Duration (years) ± SD |
|---|---|---|---|---|
| HC | 30 | 10/20 | 51.4 ± 3.3 | NA |
| PP-MS | 22 | 8/14 | 51.5 ± 7.9 | 9.1 ± 5.5 |
| Gene | Forward | Reverse |
|---|---|---|
| TGM2_v1 (full-length) * | 5′ CCTTACGGAGTCCAACCTCA 3′ | 5′ CCGTCTTCTGCTCCTCAGTC 3′ |
| TGM2_v2 | 5′ TACCCAGAGGGGTCCTCAGA 3′ | 5′ GGAACACAGGGCTTTACCAGA 3′ |
| TGM2_v3 * | 5′ GGTGAGTGGCATGGTCAACT 3′ | 5′ AGGGCTCATGACCCACATC 3′ |
| TGM2_v4a * | 5′ CCTTACGGAGTCCAACCTCA 3′ | 5′ CTGGGATGTGGAGGTGCA 3′ |
| TGM2_v4b * | 5′ CCTTACGGAGTCCAACCTCA 3′ | 5′ CACTGGTGTGGAGGTGCAGC 3′ |
| POLR2F | 5′ GAACTCAAGGCCCGAAAG 3′ | 5′ TGATGATGAGCTCGTCCAC 3′ |
| HPRT1 | 5′ AGCCCTGGCGTCGTGATTAGT 3′ | 5′ CGAGCAAGACGTTCAGTCCTGTCC 3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sestito, C.; Brevé, J.J.P.; Killestein, J.; Teunissen, C.E.; Wilhelmus, M.M.M.; Drukarch, B.; Van Dam, A.-M. Differential Expression of Tissue Transglutaminase Splice Variants in Peripheral Blood Mononuclear Cells of Primary Progressive Multiple Sclerosis Patients. Med. Sci. 2018, 6, 108. https://doi.org/10.3390/medsci6040108
Sestito C, Brevé JJP, Killestein J, Teunissen CE, Wilhelmus MMM, Drukarch B, Van Dam A-M. Differential Expression of Tissue Transglutaminase Splice Variants in Peripheral Blood Mononuclear Cells of Primary Progressive Multiple Sclerosis Patients. Medical Sciences. 2018; 6(4):108. https://doi.org/10.3390/medsci6040108
Chicago/Turabian StyleSestito, Claudia, John J. P. Brevé, Joep Killestein, Charlotte E. Teunissen, Micha M. M. Wilhelmus, Benjamin Drukarch, and Anne-Marie Van Dam. 2018. "Differential Expression of Tissue Transglutaminase Splice Variants in Peripheral Blood Mononuclear Cells of Primary Progressive Multiple Sclerosis Patients" Medical Sciences 6, no. 4: 108. https://doi.org/10.3390/medsci6040108