

The Effect of Foam-Recycle on Ammonium Removal by Aerobic Denitrification Using Alcaligenes faecalis No. 4

1
Department of Bio and Environmental Health, Dongnam Health University, Suwon 16328, Republic of Korea
2
Department of Environmental Engineering, Anyang University, Anyang 14028, Republic of Korea
*
Author to whom correspondence should be addressed.
Environments 2023, 10(10), 184; https://doi.org/10.3390/environments10100184
Submission received: 17 August 2023
/
Revised: 9 October 2023
/
Accepted: 10 October 2023
/
Published: 18 October 2023
(This article belongs to the Special Issue Advanced Technologies of Water and Wastewater Treatment)
Abstract
:Aerobic denitrifier Alcaligenes faecalis No. 4 removes ammonium-nitrogen to nitrogen gas via denitrification in a single aerobic condition. In our previous studies, factors such as ammonium removal rate, denitrification ratio, and cell growth were tested in various conditions. The removal pathway from ammonium to nitrogen gas still needs to be determined in detail. To clarify this pathway of Alcaligenes faecalis No. 4, we in this study investigated the effects of several factors on ammonium removal, such as foam-recycle, initial pH, initial ammonium concentration, and airflow rate. Denitrification ratio was improved by up to 23% through foam-recycle. The improvement of the denitrification ratio was resulted by the higher enzyme activity of hydroxylamine oxidoreductase (HAO) in the produced foam, which was about 28 times higher than that in the culture broth (i.e., without foam-recycle). The stripped ammonia was significantly high (above pH 9). The initial ammonium concentration and airflow rate also influenced the denitrification ratio.
1. Introduction
Traditionally, biochemical or biological ammonium removal from wastewater is commonly carried out using autotrophic nitrification by nitrifiers under aerobic conditions and heterotrophic denitrification by denitrifying bacteria under anaerobic or anoxic conditions [1]. The disadvantages of this approach are mainly caused by autotrophic nitrifiers. Since those are slow growers and are vulnerable to organic matter, two separate reactors or two-chambers (aerobic and anaerobic) are needed to convert NH4+-N to N2 [2,3,4,5,6]. As an alternative to this traditional approach, microorganisms that have faster growth rates than autotrophic nitrifiers and the ability to convert ammonium to N2 by simultaneously heterotrophic nitrification and aerobic denitrification have attracted significant attention [7,8,9,10,11,12,13]. These organisms assimilate a part of nitrogen and organic carbon to their cell mass, and their assimilation ratio commonly increases at high ratios of C/N, resulting in decrease in denitrification ratio [14,15]. In our previous studies, the bacterium A. faecalis No. 4 (No. 4) has such heterotrophic nitrification and aerobic denitrification abilities, as well as rapid growth rates (doubles every 2.4 h) [16]. No. 4 denitrified 40~50%, assimilated 50%, and nitrified less than 10% of removed ammonium [16]. The ammonium removal rate of No. 4 was estimated at 20~30 mg-N/L·h, which was also significantly higher than that of other bacteria with similar abilities [16,17].
Operational conditions such as initial pH, initial ammonium concentration, airflow rate, etc. may influence the performance of ammonium removal processes. Furthermore, ammonia stripping is generally affected by pH conditions in the aerated cultivation, since pH controls the concentration of free ammonia. An increase in pH increases free ammonia [18,19], which leads to increase in ammonia stripping via aeration. In aerobic operation conditions, excessive oxygen or initial ammonium and carbon concentrations may also affect heterotrophic nitrification and aerobic denitrification [20,21,22,23,24,25,26,27]. Our previous studies revealed that No. 4 characteristically produced significant foam over the aerated culture medium [16]. The viable cell number of No. 4 in the foam was relatively higher than that in the culture broth, implying that foam-recycle may be a potential or key strategy for maintaining high cell mass in the reactor. Therefore, foam-recycle would consequently promote nitrogen removal efficiency and this foam-recycle will be a key operation condition in the aerobic cultivation of No. 4.
There are several enzymes which plays important role in biochemical nitrogen cycles. In those enzymes, the hydroxylamine oxidoreductase (HAO) and nitrite reductase (NIR) are involved in the denitrification process. Traditionally in this process, HAO oxidizes hydroxylamine to nitrite then NIR denitrifies nitrite under anaerobic condition. A few past studies have demonstrated that hydroxylamine was removed through HAO generated by heterotrophic nitrifier-aerobic denitrifier species [28,29,30,31,32]. HAO produced by A. faecalis strain TUD was reported to influence the denitrification directly to N2 from ammonium under aerobic conditions [33,34,35]. A. faecalis No. 4 was shown to oxidize ammonium to hydroxylamine, nitrite and nitrate, simultaneously to denitrify ammonium to nitrogen and nitrous dioxide under aerobic conditions [16]. However, ammonium removal pathway based on enzymes mediated by No. 4 was not clarified, especially the activities of HAO and NIR are suspected to be in play in those processes.
In this study, removal characteristics of high-strength ammonium wastewater using A. faecalis No. 4 was studied under several operating conditions, such as foam-recycle, initial pH, airflow rate, and initial ammonium concentration. Ammonium removal rate and denitrification ratio were compared to evaluate the effects of those operation conditions. To further elucidate the activity of heterotrophic nitrification and aerobic denitrification by No. 4, the activities of enzymes HAO and NIR were also investigated.
2. Materials and Methods
2.1. Microorganism Used
A. faecalis No. 4 isolated from sewage sludge was stocked in 25% glycerol solution at −80 °C. The stock solution was inoculated into the culture medium, and cultivated for approximately 24 h. The resulting culture was then inoculated into a fresh medium and cultivated for 16 h before being used as the seed culture.
2.2. Medium
The basic medium formulating the synthetic wastewater was prepared by dissolving 14 g [K2HPO4], 6 g [KH2PO4], 51 g [trisodium citrate dihydrate], 6 g [(NH4)2SO4], 0.2 g [MgSO4·7H2O] and 2 mL [trace mineral solution], in 1 L distilled water. The C/N ratio was adjusted to around 10 which was optimum condition for No. 4 growth as determined in a previous study [16]. Trace mineral solution contained 57.1 g [EDTA·2Na], 3.9 g [ZnSO4·7H2O], 7 g [CaCl2·2H2O], 5.1 g [MnCl2·4H2O], 5.0 g [FeSO4·7H2O], 1.1 g [(NH4)6Mo7O24·4H2O], 1.6 g [CuSO4·5H2O], and 1.6 g [CoCl2·6H2O at pH 6.0], per volumetric liter. To determine the viable cell number of No. 4, L agar medium containing 1% peptone, 0.5% NaCl, 0.5% yeast extract, and 1.5% agar (pH 7.0) was used.
2.3. Aerated Batch Cultivation
The aerated batch reactor system shown in Figure 1 was used throughout this study [16]. A total of 360 mL of membrane filtered medium was poured into a 500 mL conical flask and an aliquot of 5% seed culture of No. 4 was inoculated and stirred with a magnetic stirrer at 200-cycles/min. Air was supplied from a compressor at the flow rate of 0.5 L/min. Samples of 2 mL each were taken every 8 or 12 h from the reactor for chemical analysis, measurement of pH and for microbial count. Ammonia stripped from the reactor by the aeration air was captured in a 35 mM H2SO4 solution. In previous studies, we had observed foam generation and accumulation in an aerated batch cultivation system, which turned out to be extremely rich in No. 4 biomass. We hypothesized that foam recycle could enhance this process, thus the effect of foam-recycle compared with no-foam-recycle was also investigated in this study. Foam was allowed to accumulate in a foam holder, and subsequently was continuously recycled back into the reactor using a peristaltic pump. Different conditions of initial pH, airflow rate, and ammonium concentrations were also tested to determine the effects of varying these factors on the denitrification of No. 4 under the condition of foam-recycle. The initial pH-conditions of the medium were sequentially set at 6, 7 and 8 using 2 N NaOH and 2 N HCl, while inlet ammonium concentration was adjusted at 1037~1122 ppm, and airflow rate was fixed at 0.5 L/min. To examine the effects of initial ammonium concentration and airflow rate, ammonium concentrations were sequentially set to 250, 1000, and 3000 ppm NH4+-N by adjusting amounts of sodium citrate and ammonium sulfate, and effect of airflow rates was test at 0.2, 0.5, and 0.8 L/min. Both the effects of ammonium concentrations and airflow rate were tested at a fixed pH condition of 7.0.
2.4. Shaking-Flask Cultivation
To verify metabolic properties of hydroxylamine and nitrite those related with HAO and NIR, hydroxylamine and sodium nitrite were used instead of ammonium sulfate. NH2OH-N and NO2−-N concentrations were adjusted to about 120~130 ppm and 120~170 ppm, respectively. The amount of citrate as a carbon source was adjusted to provide a C/N ratio of 10. A 150 mL of 0.2 μm membrane-filtered medium was set in a 500 mL shaking flask and 2% (0.3 mL) separated foam or 5% (0.75 mL) No. 4 culture broth were inoculated to start the cultivation with similar cell numbers. During incubation at 30 °C and shaking at 120 spm (strokes per minute), 2 mL of samples were taken at 1, 3, 6, and 16 h after seeding for chemical analyses.
2.5. Activities of Hydroxylamine Oxidoreductase (HAO) and Nitrite Reductase (NIR)
To determine the ability of HAO on the reduction of hydroxylamine in the foam and in the culture without foam-recycle, HAO activity was determined by the reduction of K3Fe(C/N)6 at 400 nm [31]. The molar absorption coefficient (ε) of K3Fe(C/N)6 was 1/mM·cm. The reaction mixture contained 5 mM [Tris/HCl buffer] at pH 8.0, 100 μM [K3Fe(C/N)6], 1 mM [NH2OH], and 400 μM [EDTA]. The optical density at 660nm (OD660) of the No. 4 cells of the separated foam were adjusted to about 0.9 by dilution with 5 mM [Tris/HCl buffer] and centrifuged at 10,000 × rotation per minute (rpm) for 10 min. The separated cells were suspended in Tris/HCl buffer and sonicated for 2 min, while the supernatants were used for HAO analysis. The reaction of the supernatant with K3Fe(C/N)6 was carried out for 2 min. The viable cell number was used for calculation of enzyme activity per colony forming unit (cfu).
Nitrite reductase (NIR) activity was determined using previously reported methods [34,35]. Briefly, test tubes were loaded with 0.675 mL of 100 mM sodium phosphate buffer (pH 7.0), 0.4 mL of 10 mM sodium nitrite, 0.5 mL of 3 mM methyl viologen, and 0.125 mL of separated foam or the culture. The reaction was started by adding 0.3 mL of 100 mM sodium hydrosulfite dissolved in 50 mM sodium bicarbonate. Control tests were conducted without foam or culture broth. After the test tubes were sealed with a cap, pre-incubation of the reaction mixture was performed at 37 °C for 5 min under anaerobic conditions and the reaction mixture was shaken until complete decolorization. A 0.05 mL of the reaction mixture was mixed with 2.95 mL sterilized water, and then 1 mL of 1% sulfanilamide and 1 mL of 0.02% N-1-naphthylenediamine were added. After allowing the reaction mixture to stand at room temperature for 20 min, nitrite detection was performed by UV-VIS spectrophotometer at 540 nm (UV-2400PC, Shimadzu, Kyoto, Japan).
2.6. Analytical Methods
Culture samples were centrifuged at 10,000 rpm, and filtered through 0.2 μm membrane filters. The filtrate was used for the analyses of NH4+-N and nitrification products (NH2OH, NO2−, NO3−), nitrous oxide (NO), nitrogen dioxide (NO2), and citrate. Concentrations of NH4+-N was analyzed by indophenol method [36,37,38], NH2OH was measured according to the method by Fear and Burrell using spectrophotometer [39]. The NO2−, NO3− and citrate were measured by ion chromatography (HIC-6A, Shimadzu, column: Shim-pack IC-A1, Shimadzu, at flow rate of 1.5 mL min−1, buffer solution containing 2.5 mmol phtalate and 2.4 mmol trisaminomethane, and at oven temperature of 40 °C). Optical density to check growth rate and No. 4 biomass at 660 nm (OD660nm) was measured by a spectrophotometer (UV-1200, Shimadzu). The HAO and NIR activities were measured by UV-VIS recording spectrophotometer (UV-2400PC, Shimadzu). Viable cell number was counted on L agar plate and expressed as colony forming units (cfu). Intracellular nitrogen content (mg-N/L) was calculated from viable cell number and nitrogen content (%) obtained from elemental analysis of dry cell mass. Dry cell was prepared by centrifugation of culture broth, then washing the cells with distilled water, and finally drying the washed cell at 105 °C for 24 h. Denitrification ratio was calculated by subtracting the amount of total nitrifying products (NH2OH-N, NO2−-N, NO3−-N), amount of nitrogen in dry cell mass (intracellular nitrogen), and stripped ammonia from removed ammonium. The NO and NO2 were analyzed by a NOx analyzer (AP1 200A, Riken Keiki Co., Ltd., Tokyo, Japan).
3. Results and Discussion
3.1. Effect of Foam-Recycle
In our previous studies, ammonium was oxidized to hydroxylamine, nitrite, and nitrate under aerobic condition by A. faecalis No. 4, providing these intermediate substrates for subsequent denitrification [16,17,40]. Hydroxylamine was utilized without citrate consumption [16]. Less than 10% of consumed hydroxylamine was converted to nitrite implying that most of hydroxylamine was converted to denitrified products (such as N2 and N2O, more than 80% N2 production) [16]. When No. 4 was cultivated under aerobic condition, significant foaming was observed. This foam was collected in a separate foam holder (Figure 1) and continuously recycled back into the reactor. About 10~20 mL of foam was obtained in the foam holder based on the 360 mL of culture medium. The OD660nm of the separated foam was significantly higher than that in the normal culture broth of No. 4.
The results of the aerated batch experiments performed with and without foam-recycle are shown in Figure 2 and Figure 3, respectively. In both cases, the consumption of carbon (75~83% citrate was consumed), pH increased and growth of No. 4 (2~3 × 1010 cfu) were similar. However, the production of hydroxylamine and nitrite were significantly different. In the culture without foam-recycle, No. 4 produced 82 ppm NO2−-N in 110 h, while a maximum of 7 ppm hydroxylamine was observed at 24 h point, after that it decreased (Figure 3B). However, in the culture with foam-recycle, No. 4 produced lower NO2−-N (13 ppm) in 93 h but extremely low production of hydroxylamine was observed (Figure 2B). If hydroxylamine produced was rapidly converted to denitrified products, almost no residual hydroxylamine was detected in the culture broth. Table 1 shows nitrogen mass balance. Ammonium removal rate was 22 mg-N/L·h and denitrification ratio was 39% in the culture without foam-recycle. A similar or slightly higher ammonium removal rate of 24 mg-N/L·h was observed in the culture with foam-recycle. However, denitrification ratio was considerably higher (48%); approximately 9% (23% on 39% basis in the culture) without foam-recycle higher than the culture without foam-recycle.
3.2. Consumption of Other Nitrogen Sources
Figure 4 and Figure 5 present the results of No. 4’s ability to metabolize other nitrogen sources in the culture with addition of either hydroxylamine or nitrite in the basic medium instead of ammonium source. Figure 4 and Figure 5 show the results with 2% foam-recycle and with 5% No. 4 culture broth (without foam-recycle), respectively. According to Figure 4A, hydroxylamine decreased from the beginning of incubation, and about 9 ppm nitrite was produced in 6 h after consumption of 110 ppm hydroxylamine, but no nitrate was detected. Hydroxylamine decreased at the rate of 1.6 × 10−10 mg-N/cfu × h ((74.54 mg-NH2OH-N/L) ÷ (7.8 × 107 cfu/mL) ÷ 6 h). Nitrite produced from hydroxylamine decreased slightly at 8 to 16 h, and no production in nitrate was observed. While in Figure 4B, when 116 ppm of NO2−-N was added to basic medium, only a decreased of 8 ppm of NO2−-N was observed in 16 h and neither NO3− nor hydroxylamine was detected. In the experiment using the culture broth (without foam-recycle) of Figure 5, hydroxylamine decline rate (7.1 × 10−11 mg-N/cfu × h = ((71.46 mg-NH2OH-N/L) ÷ (6.3 × 107 cfu/mL) ÷ 16 h)) was significantly slower than in the former experiment (Figure 4A). And the NO2−-N consumption in this experiment (Figure 4B and Figure 5B) was similar to that in both experiments and no consumptions of carbon and nitrogen sources were observed. These results suggest that A. faecalis No. 4 either with foam-recycle or without foam-recycle converts most of hydroxylamine to denitrified products and a small portion of hydroxylamine to nitrite. This fact can be explained that denitrification activities between the cultures with foam-recycle and without foam-recycle were different in Table 1, indicating 48% and 39%, respectively.
3.3. Enzyme Activity
Table 2 shows hydroxylamine oxidoreductase (HAO) activities between the cultures with foam-recycle and without foam-recycle. Measurements of hydroxylamine oxidoreductase (HAO) activity confirmed the existence of HAO activity in both cultures with foam-recycle and without foam-recycle. The HAO activity in the foam was significantly higher than that in the culture without foam-recycle. The activity of HAO per 1 mL of culture broth with foam-recycle was about 28 times higher than that in the culture without foam-recycle and the activity per cfu in the foam was about five times than that in the culture without foam-recycle.
Measurements of hydroxylamine oxidoreductase (HAO) and nitrite oxidoreductase (NIR) activities to evaluate their involvement in the denitrification process were adopted from previous studies [14,33,35,41,42,43,44,45] The activity of HAO by No. 4 was 10~100 times higher than the previously reported activity levels [33,41,45]. The reason behind this higher activity is not clear yet, but the cells attached to foam significantly enhanced the denitrification ratio with higher HAO activity. This is primarily due to the fact No. 4 (aerobic bacteria) more functionally exhibited their aerobic denitrification ability on the cell-air interface of foam surface.
Although A. faecalis is known to denitrify hydroxylamine [33,35,45], denitrification from nitrite has not been reported before in past studies. Figure 6 shows nitrite reductase (NIR) activity measured under anaerobic condition using methyl viologen [36,46]. Activity of NIR was observed both in foam and foam-free culture (No. 4 culture broth). The NIR activity in the culture without foam-recycle was about 25% of that in the culture with foam-recycle. However, NIR activity (nmol-nitrite/μL-foam · min) was significantly lower, compared to HAO activities (Table 2) and NIR activity produced by other microorganisms under anaerobic conditions [47]. The likelihood that NIR activity is associated with denitrification under aerobic conditions is negligible, and therefore the dominant denitrification pathway of No. 4 is most likely via hydroxylamine under HAO.
3.4. Effect of Initial Ammonium Load and Airflow Rate
The effect of initial ammonium concentration on ammonium removal was investigated at 250, 1000, and 3000 ppm under different airflow rates. The nitrogen mass balances for these experiments are shown in Table 3. Enhanced denitrification ratio and ammonium removal rate at the higher ammonium concentrations (1000 and 3000 ppm) were observed at higher airflow rates (0.5 and 0.8 L/min) compared with that at lower airflow rate of 0.2 L/min.
Denitrification ratio at 3000 ppm initial ammonium level was more than 60% and the ammonium removal rate was the highest. These results indicated that the airflow rate is critical for denitrification ability of No. 4 especially for high-strength ammonium wastewater. Higher ammonium removal rate was achieved at elevated ammonium concentration levels and air supply. Ammonia stripping was also significant at higher ammonium concentrations (i.e., ammonia stripping increased with increase in ammonium concentration). The highest ammonia (170 ppm) which was about 15% of initial ammonium concentration was stripped out at pH 8, while only 79 and 41 ppm of NH3-N were stripped out at pH 6 and 7, respectively. Ammonium concentration had a higher influence on ammonia stripping than airflow rate as is evident based on data presented in Table 3. Approximately 10~30% ammonia was stripped when initial of NH4+-N was 3000 ppm at the three airflow rates, suggesting that nitrogen loss via air stripping should be a concern during treatment of high-strength ammonium wastewater.
3.5. Effect of pH
The nitrogen mass balance and ammonium removal rate after the 96 h of aerated batch process at different initial pH-conditions are shown in Table 4. Ammonium removal rates at pH 7 and 6 were 25 mg-N/L·h, and 20 mg-N/L·h, respectively, which were higher than that at pH 8. The denitrification ratio at pH 6 was higher than that at either pH 7. However, similar denitrification ratios at pH 6 and 7 suggested that the initial pH in aerobic cultivation with No. 4 should be maintained in the range of 6 to 7. Ammonia stripping was significantly dependent on pH-conditions (Figure 7). Extremely low ammonia stripping occurred below pH of 8.5, but this situation changed rapidly in higher pH connditions and ammonia stripping became significant above pH 9. This phenomenon has been well documented in past research and this was atributed to the increase (or release) in the concentration of free ammonia with increase in the pH [11,48]. The reported pH values of high-strength ammonium wastewaters range from 7.5 to 8.5 [49,50,51,52,53]. Given that 7.5 to 8.5 is natural pH range of high-strength wastewater, it is quite apparent that pH control at around 7 would be the optimal compromise for No. 4 to exhibit the maximum capability of denitrification and also to minimize ammonia stripping.
The total nitrogen mass balance in the ammonium removal and the removal pathway by No. 4 is shown in Figure 8. This figure was prepared based on the data presented in Figure 2 and some results from previous research [16]. No. 4 removed 51% of ammonium by means of heterotrophic-nitrification and 45% by assimilation into cells. At least 48% of removed ammonium (51%) was converted to denitrified products by HAO activity which was significantly high in the produced foam. Except for a part of assimilation, most of the ammonium (87%) denitrified via hydroxyamine.
4. Conclusions
Factors that affect ammonium removal rate and denitrification ratio of No. 4 include the following foam-recycle, initial pH, airflow rate, and initial ammonium concentration were investigated in this study. Ammonium was first oxidized to hydroxylamine, nitrite, and nitrate under aerobic condition by No. 4, providing these intermediate substrates for subsequent denitrification. Hydroxylamine was utilized without citrate consumption. Less than 10% of consumed hydroxylamine was converted to nitrite, implying that most of hydroxylamine was converted to denitrified products (such as N2 and N2O).
Measurements of hydroxylamine oxidoreducatse (HAO) activity confirmed the existence of HAO activity both in the foam and the culture broth. In autotrophic nitrifiers, HAO was observed to convert hydroxylamine to NO2−. However, in A. faecalis which conducts heterotrophic nitrification and aerobic denitrification, HAO is believed to work also as an enzyme to convert hydroxylamine to denitrified product via HNO. In No. 4 biochemical processes, HAO converted more than 90% of hydroxylamine to denitrified products and 10% to nitrite. The activity of HAO in the foam was significantly higher than that in the culture broth.
A higher ammonium removal rate was achieved at elevated ammonium levels and air supply. Ammonia stripping, however, was significant at higher ammonium concentration (i.e., ammonia stripping increased with increase in ammonium concentration).
No. 4 removed 51% of ammonium by means of heterotrophic-nitrification and 45% by assimilation into cells. At least 48% of removed ammonium was converted to denitrified products. Evidently, under controlled conditions, No. 4 can be used in the treatment of wastewaters with high concentrations of ammonium, such as a piggery and dairy wastewater. Future research to advance the current work should be conducted with these kinds of wastewaters.
Author Contributions
Conceptualization, methodology, formal analysis, investigation, writing review and editing, H.-S.J.; data curation and writing-original draft preparation, K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the research project of “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ017075)” Rural Development Administration of Korea and the support of Particulate Matter Management Specialized Graduate Program through the Korea Environmental Industry & Technology Institute (KEITI) funded by the Ministry of Environment (MOE).
Data Availability Statement
Data from this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McCatry, P.L. What is the Best Biological Process for Nitrogen Removal: When and Why? Environ. Sci. Technol. 2018, 52, 3835–3841. [Google Scholar] [CrossRef]
- Hooper, A.B.; Vannelli, T.; Bergmann, D.J.; Arociero, D.M. Enzymology of the oxidation of ammonia to nitrite by bacteria. Antonie Van Leeuwenhoek 1997, 71, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Van Loosdrecht, M.C.M.; Jetten, M.S.M. Microbiological conversion in nitrogen removal. Water Sci. Technol. 1998, 38, 1–7. [Google Scholar] [CrossRef]
- Itokawa, H.; Hanaki, K.; Matsuo, T. Nitrous oxide production in high-loading biological nitrogen removal process under low COD/N ratio condition. Water Res. 2001, 35, 657–664. [Google Scholar] [CrossRef]
- Wrage, N.; Velthof, G.L.; van Beusichem, M.L.; Oenema, O. Role of nitrifer denitrification in the production of nitrous oxide. Soil Biol. Biochem. 2001, 33, 1723–1732. [Google Scholar] [CrossRef]
- Carrera, J.; Baeza, J.A.; Vicent, T.; Lafuente, J. Biological nitrogen removal of high-strength ammonium industrial wastewater with two-sludge system. Water Res. 2003, 37, 4211–4221. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Logemann, S.; Muyzer, G.; Robertson, L.A.; de Vries, S.; van Loosdrecht, M.C.M.; Kuenen, J.G. Novel principles in the microbial conversion of nitrogen compounds. Antonie Van Leeuwenhoek 1997, 71, 75–93. [Google Scholar] [CrossRef]
- Gupta, A.B.; Gupta, S.K. Simultaneous carbon and nitrogen removal in mixed culture aerobic RBC biofilm. Water Res. 1999, 33, 556–561. [Google Scholar] [CrossRef]
- Gupta, A.B.; Gupta, S.K. Simultaneous carbon and nitrogen removal from high strength domestic wastewater in an aerobic RBC biofilm. Water Res. 2001, 35, 1714–1722. [Google Scholar] [CrossRef]
- Schmidt, I.; Sliekers, O.; Schmid, M.; Bock, E.; Fuerst, J.; Kuenen, J.G.; Jetten, M.S.M.; Strous, M. New concepts of microbial treatment processes for the nitrogen removal in wastewater. FEMS Microbiol. Rev. 2003, 772, 481–492. [Google Scholar] [CrossRef]
- Khin, T.; Annachhatre, A.P. Novel microbial nitrogen removal processes. Biotechnol. Adv. 2004, 22, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, L.; Pan, D.; Lin, H.; Ren, Y.; Zhang, J.; Zhou, B.; Lin, J.; Lin, J. Heterotrophic nitrification and related functional gene expression characteristics of Alcaligenes faecalis SDU20 with the potential use in swine wastewater treatment. Bioprocess Biosyst. Eng. 2021, 44, 2035–2050. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, J.; Zhang, S.; Liu, F.; Peng, J.; Peng, Y.; Wu, J. Nitrogen removal characteristics of a novel heterotrophic nitrification and aerobic denitrification bacteria, Alcaligenes faecalis WT14. J. Environ. Manag. 2021, 282, 111961. [Google Scholar] [CrossRef] [PubMed]
- Jetten, M.S.M.; de Bruijn, P.; Kuenen, G. Hydroxylamine metabolism in Pseudomonas PB16: Involvement of a novel hydroxylamine oxidoreductase. Antonie Van Leeuwenhoek 1997, 71, 69–74. [Google Scholar] [CrossRef]
- Wang, Y.; Singh, R.P.; Geng, C.; Fu, D. Carbon to nitrogen ratio influence on the performance of bioretention for wastewater treatment. Environ. Sci. Pollut. Res. 2020, 27, 17652–17660. [Google Scholar] [CrossRef]
- Joo, H.S.; Hirai, M.; Shoda, M. Characteristics of ammonium removal by hetetrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef]
- Joo, H.S.; Hirai, M.; Shoda, M. Piggery wastewater treatment using Alcaligenes faecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification. Water Res. 2006, 40, 3029–3036. [Google Scholar] [CrossRef]
- Yang, S.F.; Tay, J.H.; Liu, Y. Inhibition of free ammonia to the formation of aerobic granules. Biochem. Eng. J. 2004, 17, 41–48. [Google Scholar] [CrossRef]
- Liu, Y.; Ngo, H.H.; Guo, W.; Peng, L.; Wnag, D.; Ni, B. The roles of free ammonia (FA) in biological wastewater treatment processes: A review. Environ. Int. 2019, 123, 10–19. [Google Scholar] [CrossRef]
- Liu, Y.; Capdeville, B. Response pattern of nitrifying biofilm reactor to shock loading. Biotechnol. Lett. 1994, 16, 655–660. [Google Scholar] [CrossRef]
- Kim, N.J.; Sugano, Y.; Hirai, M.; Shoda, M. Removal of a high load of ammonia gas by a marine bacterium, Vibrio alginolyticus. J. Biosci. Bioeng. 2000, 90, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Uemoto, H.; Ando, A.; Saiki, H. Effect of oxygen concentration on nitrogen removal by Nitrosomonas europaea and Paracoccus denitrificans immobilized within tubular polymeric gel. J. Biosci. Bioeng. 2000, 6, 654–660. [Google Scholar] [CrossRef]
- Su, J.J.; Liu, B.Y.; Liu, C.Y. Comparison of aerobic denitrification under high oxygen atmosphere by Thiosphaera pantotropha ATCC 35512 and Pseudomonas stutzeri SU2 newly isolated from the activated sludge of a piggery wastewater treatment system. J. Appl. Microbiol. 2001, 90, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.; Obbard, J.P. Ammonia removal from prawn aquaculture water using immobilized nitrifying bacteria. Appl. Microbiol. Biotechnol. 2001, 57, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaka, E.; Nomura, N.; Nakajima-Kambe, T.; Okada, N.; Nakahara, T. A simple screening procedure for heterotrophic nitrifying bacteria with oxygen-tolerant denitrification activity. J. Biosci. Bioeng. 2003, 95, 409–411. [Google Scholar] [CrossRef]
- Obaja, D.; Mace, S.; Sans, C.C.; Alvarez, J.M. Nitrification, denitrification and biological phosphorus removal in piggery wastewater using a sequencing batch reactor. Bioresour. Technol. 2003, 87, 103–111. [Google Scholar] [CrossRef]
- Du, X.; Wang, J.; Jegatheesan, V.; Shi, G. Dissolved Oxygen Control in Activated Sludge Process Using a Neural Network-Based Adaptive PID Algorithm. Appl. Sci. 2018, 8, 261. [Google Scholar] [CrossRef]
- Padhi, S.K.; Tripathy, S.; Mohanty, S.; Maiti, N.K. Aerobic and heterotrophic nitrogen removal by Enterobacter cloacae CF-S27 with efficient utilization of hydroxylamine. Bioresour. Technol. 2017, 232, 285–296. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, E.; Huang, S. Heterotrophic nitrogen removal in Bacillus sp. K5: Involvement of a novel hydroxylamine oxidase. Water Sci. Technol. 2017, 76, 3461–3467. [Google Scholar] [CrossRef]
- Liu, Y.; Ni, G.-M.; Wu, M.-R.; Li, S.-S.; Miao, L.-L.; Liu, Z.-P. Photobacterium sp. NNA4, an efficient hydroxylamine-transforming heterotrophic nitrifier/aerobic denitrifier. J. Biosci. Bioeng. 2019, 128, 64–71. [Google Scholar] [CrossRef]
- Huang, F.; Pan, L.; He, Z.; Zhang, M.; Zhang, M. Identification, interactions, nitrogen removal pathways and performances of culturable heterotrophic nitrification-aerobic denitrification bacteria from mariculture water by using cell culture and metagenomics. Sci. Total Environ. 2020, 732, 13268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, A.; Yao, Q.; Wu, Q.; Zhu, H. Nitrogen removal characteristics of a versatile heterotrophic nitrifying-aerobic denitrifying bacterium, Pseudomonas bauzanensis DN13-1, isolated from deep-sea sediment. Bioresour. Technol. 2020, 305, 122626. [Google Scholar] [CrossRef] [PubMed]
- Otte, S.; Schalk, J.; Kuenen, J.G.; Jetten, M.S.M. Hydroxylamine oxidation and subsequent nitrous oxide production by the heterotrophic ammonia oxidizer Alcaligenes faecalis. Appl. Microbiol. Biotechnol. 1999, 51, 255–261. [Google Scholar] [CrossRef]
- Zhao, B.; An, Q.; Liang, H.Y.; Guo, J.S. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresour. Technol. 2012, 116, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, H.; Liu, Y.-X.; Ren, R.-P.; Lv, Y.-K. Effect of temperature, salinity, heavy metals, ammonium concentration, pH and dissolved oxygen on ammonium removal by an aerobic nitrifier. RCS Adv. 2015, 5, 79988–79996. [Google Scholar] [CrossRef]
- Suruga, K.; Murakami, K.; Taniyama, Y.; Hama, T.; Chida, H.; Satoh, T.; Yamada, S.; Hakamata, W.; Kawachi, R.; Isogai, Y.; et al. A novel microperoxidase activity: Methyl viologen-linked nitrite reducing activity of microperoxidase. Biochem. Biophys. Res. Commun. 2004, 315, 815–822. [Google Scholar] [CrossRef]
- Da Silva, S.; Cosnier, S.; Almeida, M.G.; Moura, J.J.G. An efficient poly(pyrrole-viologen)-nitrite reductase biosensor for the mediated detection of nitrite. Electrochem. Commun. 2004, 6, 404–408. [Google Scholar] [CrossRef]
- The Indophenol Blue Absorptiometric Method (JIS) k-0102, Japanese Industrial Standard; Japanese Standard Association: Tokyo, Japan, 1991; Volume 42, pp. 134–145. Available online: https://www.jsa.or.jp/en/ (accessed on 23 July 2023).
- Frear, D.S.; Burrell, R.C. Spectrophotometric methods for determining hydroxylamine reductase activity in higher plants. Anal. Chem. 1955, 27, 1664–1665. [Google Scholar] [CrossRef]
- Wako, R. Removal Characteristics of Ammonium by Heterotrophic Nitrifier Alcaligenes faecalis No. 4; Tokyo Institute of Technology: Yokohama, Japan, 2002. [Google Scholar]
- Hooper, A.B.; Balny, C. Reaction of oxygen with hydroxylamine oxidoreductase of Nitrosomonas fast kinetics. FEBS Lett. 1982, 144, 229–303. [Google Scholar] [CrossRef]
- Sasaki, S.; Karube, I.; Hirota, N.; Arikawa, Y.; Nishiyama, M.; Kukimoto, M.; Horinouchi, S.; Beppu, T. Application of nitrite reductase from Alcaligenes faecalis S-6 for nitrite measurement. Biosens. Bioelectron. 1998, 13, 1–5. [Google Scholar] [CrossRef]
- Song, B.; Ward, B.B. Nitrite Reductase genes in halobenzoate degrading denitrifying bacteria. FEMS Microbiol. Ecol. 2003, 43, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhao, X.; Cheng, Y.; Jiang, M.; Li, X.; Xue, G. Iron Robustly Stimulates Simultaneous Nitrification and Denitrification Under Aerobic Conditions. Environ. Sci. Technol. 2018, 52, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Lee, K.; Shoda, M.; Mwangi, N.P. Application of Alcaligenes faecalis No. 4 for treatment of high-strength ammonium wastewater. Environ. Eng. Manag. J. 2020, 19, 589–598. Available online: http://www.eemj.icpm.tuiasi.ro/ (accessed on 21 June 2023).
- Bueno, E.; Sit, B.; Waldor, M.K.; Cava, F. Anaerobic nitrate reduction divergently governs population expansion of the enteropathogen Vibrio cholerae. Nat. Microbiol. 2018, 3, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Polcyn, W.; Lucinski, R. Aerobic and anaerobic nitrate and nitrite reduction in free-living cells of Bradyrhizobium sp. (Lupinus). FEMS Microbiol. Lett. 2003, 226, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Mohammed-Nour, A.; Al-Sewailem, M.; El-Naggar, A.H. The Influence of Alkalization and Temperature on Ammonia Recovery from Cow Manure and the Chemical Properties of the Effluents. Sustainability 2019, 1, 2441. [Google Scholar] [CrossRef]
- Beline, F.; Martinez, J.; Marol, C.; Guiraud, G. Application of the 15N technique to determine the contributions of nitrification and denitrification to the flux of nitrous oxide from aerated pig slurry. Water Res. 2001, 35, 2774–2778. [Google Scholar] [CrossRef]
- Pollice, A.; Tandoi, V.; Lestingi, C. Influence of aeration and sludge retention time on ammonium oxidation to nitrite and nitrate. Water Res. 2002, 36, 2541–2546. [Google Scholar] [CrossRef]
- Uygur, A.; Kargi, F. Biological nutrient removal from pre-treated landfill leachate in a sequencing batch reactor. J. Environ. Manag. 2004, 71, 9–14. [Google Scholar] [CrossRef]
- Obaja, D.; Mace, S.; Alvarez, J.M. Biological nutrient removal by a sequencing batch reactor (SBR) using an internal organic carbon source in digested piggery wastewater. Bioresour. Technol. 2005, 96, 7–14. [Google Scholar] [CrossRef]
- Neerackal, G.M.; Ndegwa, P.M.; Joo, H.S.J.; Harrison, H. Manure-pH management for ammonia emission from dairy barns and liquid manure storage. Appl. Eng. Agric. 2017, 33, 235–242. [Google Scholar] [CrossRef]
Figure 1.
A schematic diagram of an aerated batch reactor used in this study. (1: reactor (500 mL conical flask), 2: stirrer, 3: magnetic bar, 4: sampling syringe, 5: temperature controller, 6: flow meter, 7: air filter, 8: air compressor, 9: foam holder, 10: NOx analyzer, 11: ammonia trap bottle, 12: water bath, 13: peristaltic pump (p), 14: gas effluent).
Figure 1.
A schematic diagram of an aerated batch reactor used in this study. (1: reactor (500 mL conical flask), 2: stirrer, 3: magnetic bar, 4: sampling syringe, 5: temperature controller, 6: flow meter, 7: air filter, 8: air compressor, 9: foam holder, 10: NOx analyzer, 11: ammonia trap bottle, 12: water bath, 13: peristaltic pump (p), 14: gas effluent).
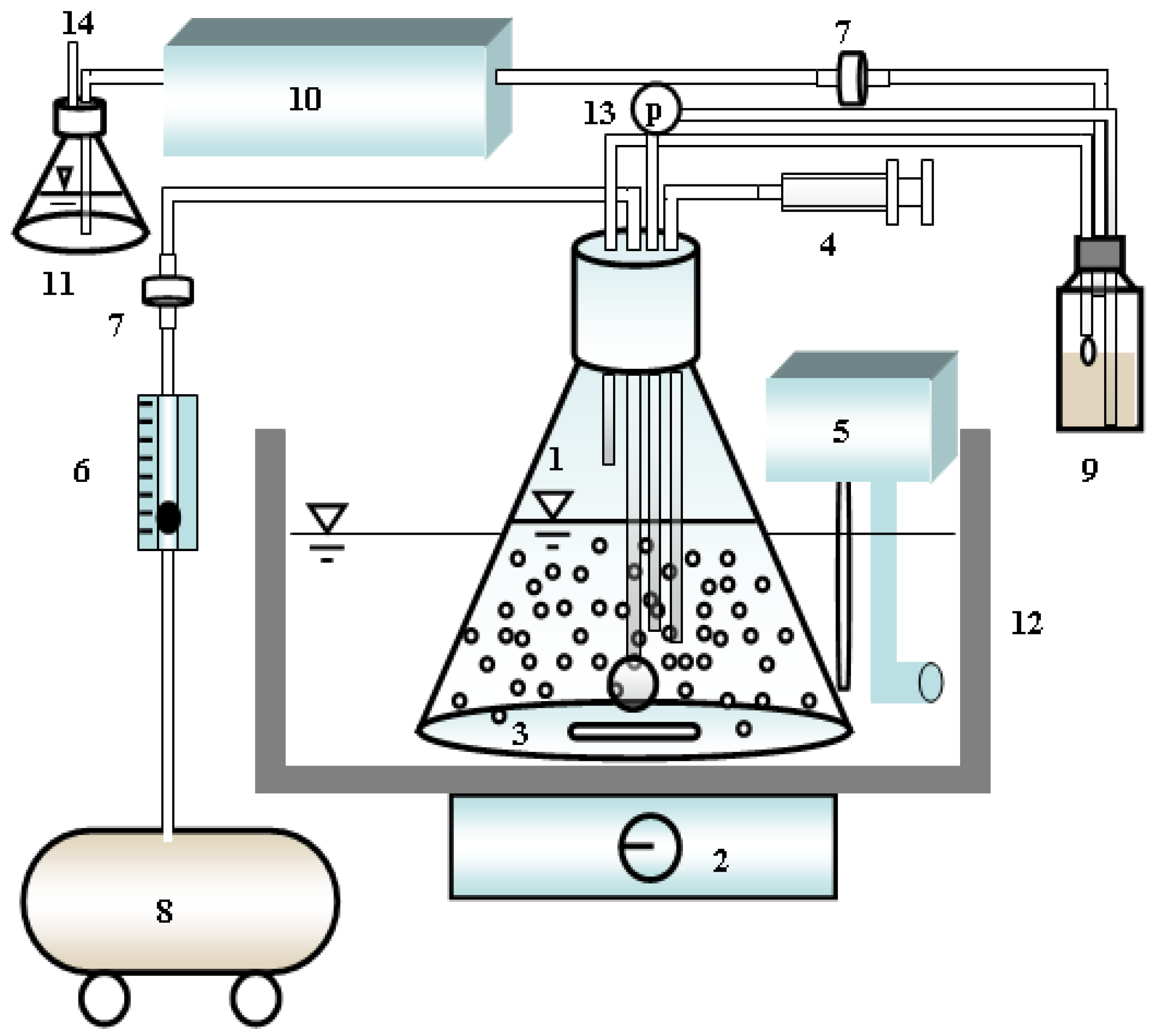
Figure 2.
Changes of nitrogen species and substrates, the pH in the culture medium, citrate concentration, and viable cell numbers in the aerated batch experiments with foam recycling (mean values of two replications). (A): pH (◆), viable cell number (○) and citrate-C (▲), (B): NH4+-N (◇), intracellular nitrogen (○), NH2OH-N (▲), NO2−-N (◆), stripped NH3-N (■), and NO3−-N (●).
Figure 2.
Changes of nitrogen species and substrates, the pH in the culture medium, citrate concentration, and viable cell numbers in the aerated batch experiments with foam recycling (mean values of two replications). (A): pH (◆), viable cell number (○) and citrate-C (▲), (B): NH4+-N (◇), intracellular nitrogen (○), NH2OH-N (▲), NO2−-N (◆), stripped NH3-N (■), and NO3−-N (●).
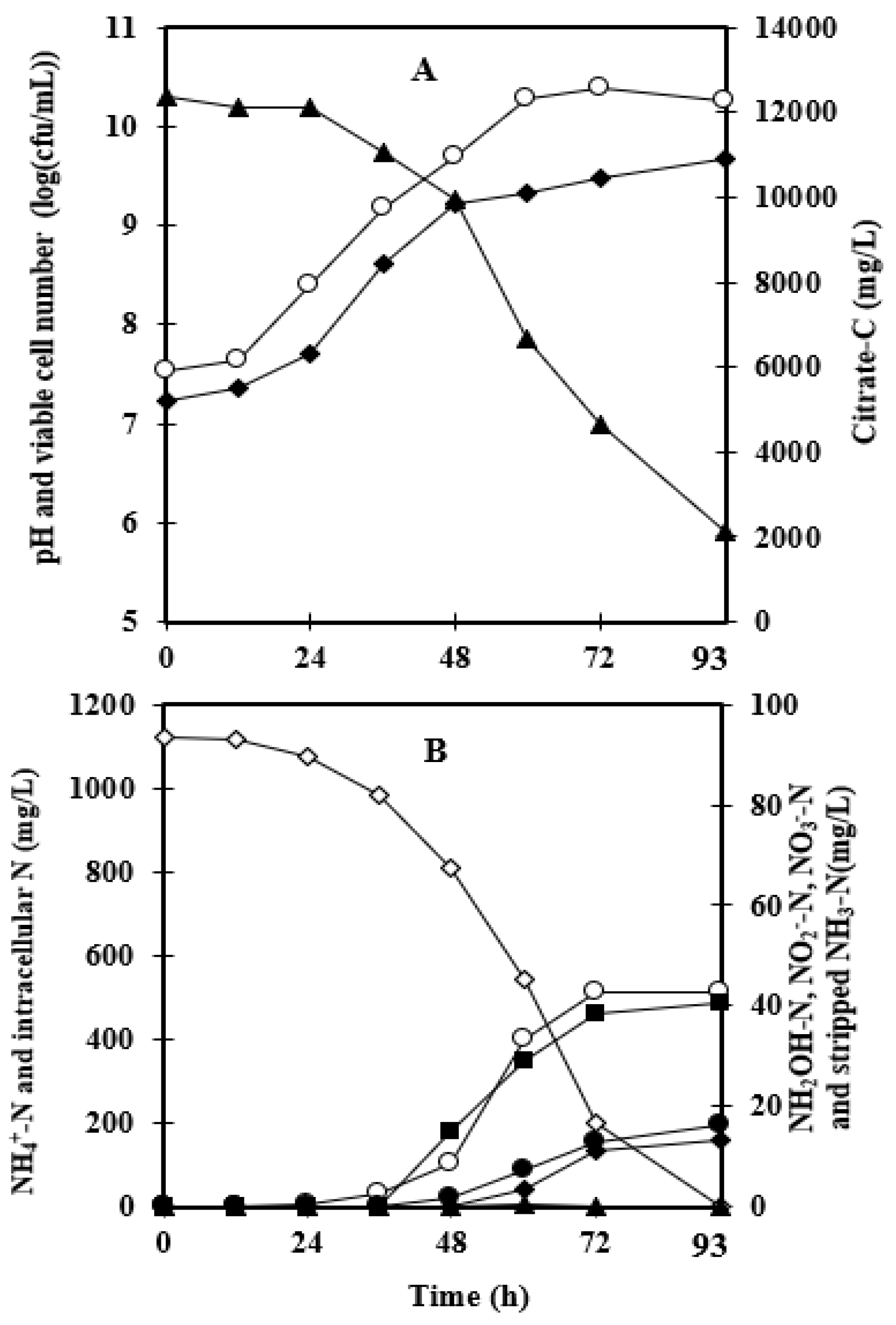
Figure 3.
Changes of nitrogen species and substrates, the pH in the culture medium, citrate concentration, and viable cell numbers in the aerated batch experiments without foam recycling (mean values of two replications). (A): pH (◆), viable cell number (○) and citrate-C (▲), (B): NH4+-N (◇), intracellular nitrogen (○), NH2OH-N (▲), NO2−-N (◆), stripped NH3-N (■) and NO3−-N (●).
Figure 3.
Changes of nitrogen species and substrates, the pH in the culture medium, citrate concentration, and viable cell numbers in the aerated batch experiments without foam recycling (mean values of two replications). (A): pH (◆), viable cell number (○) and citrate-C (▲), (B): NH4+-N (◇), intracellular nitrogen (○), NH2OH-N (▲), NO2−-N (◆), stripped NH3-N (■) and NO3−-N (●).
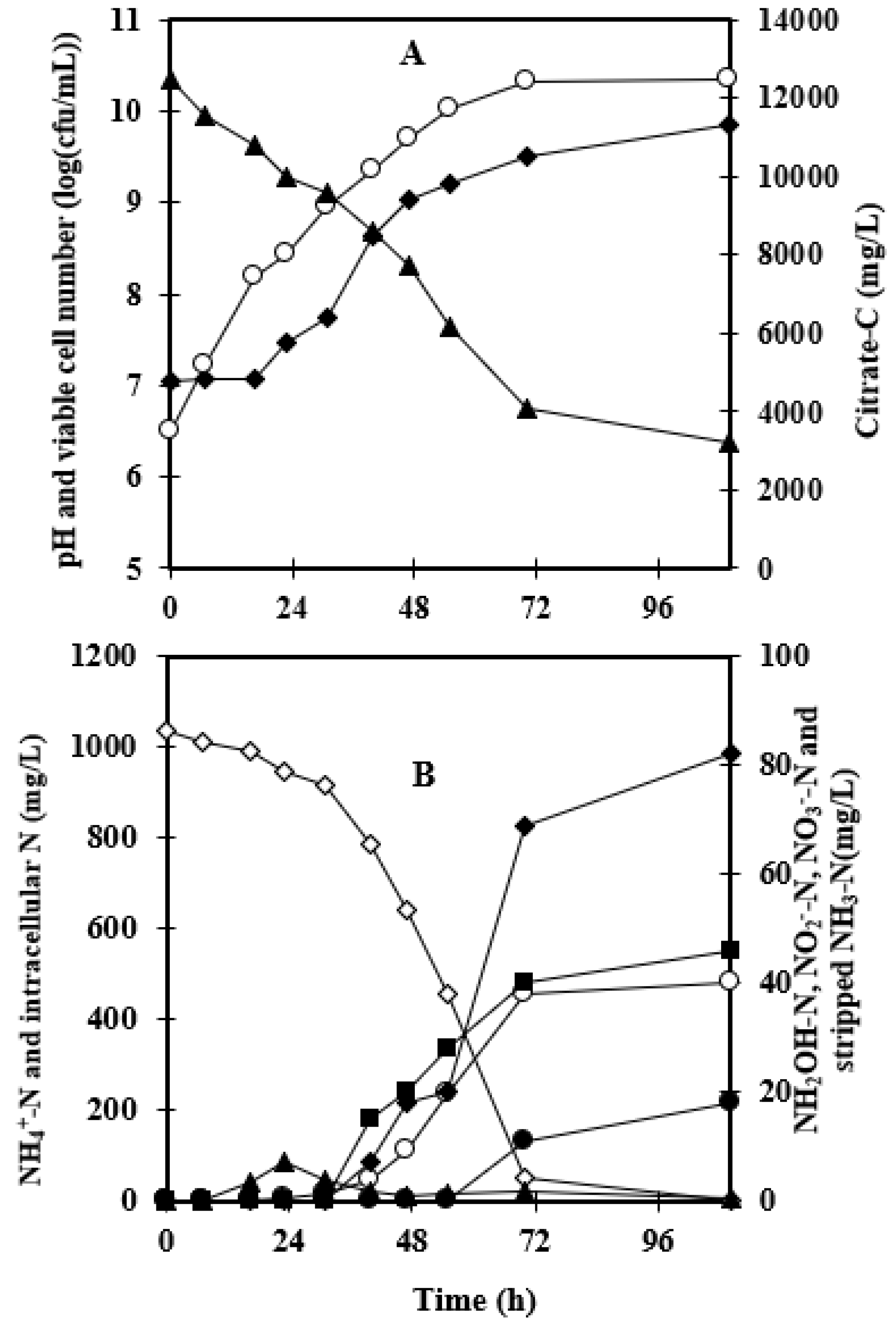
Figure 4.
Removal of nitrogen spieces (with hydroxylamine (A) and nitrite (B) instead of ammonium) and carbon consumption in batch reactors with produced foam. Hydroxylamine (◆), nitrite (▲), nitrate (○), and citrate (□).
Figure 4.
Removal of nitrogen spieces (with hydroxylamine (A) and nitrite (B) instead of ammonium) and carbon consumption in batch reactors with produced foam. Hydroxylamine (◆), nitrite (▲), nitrate (○), and citrate (□).
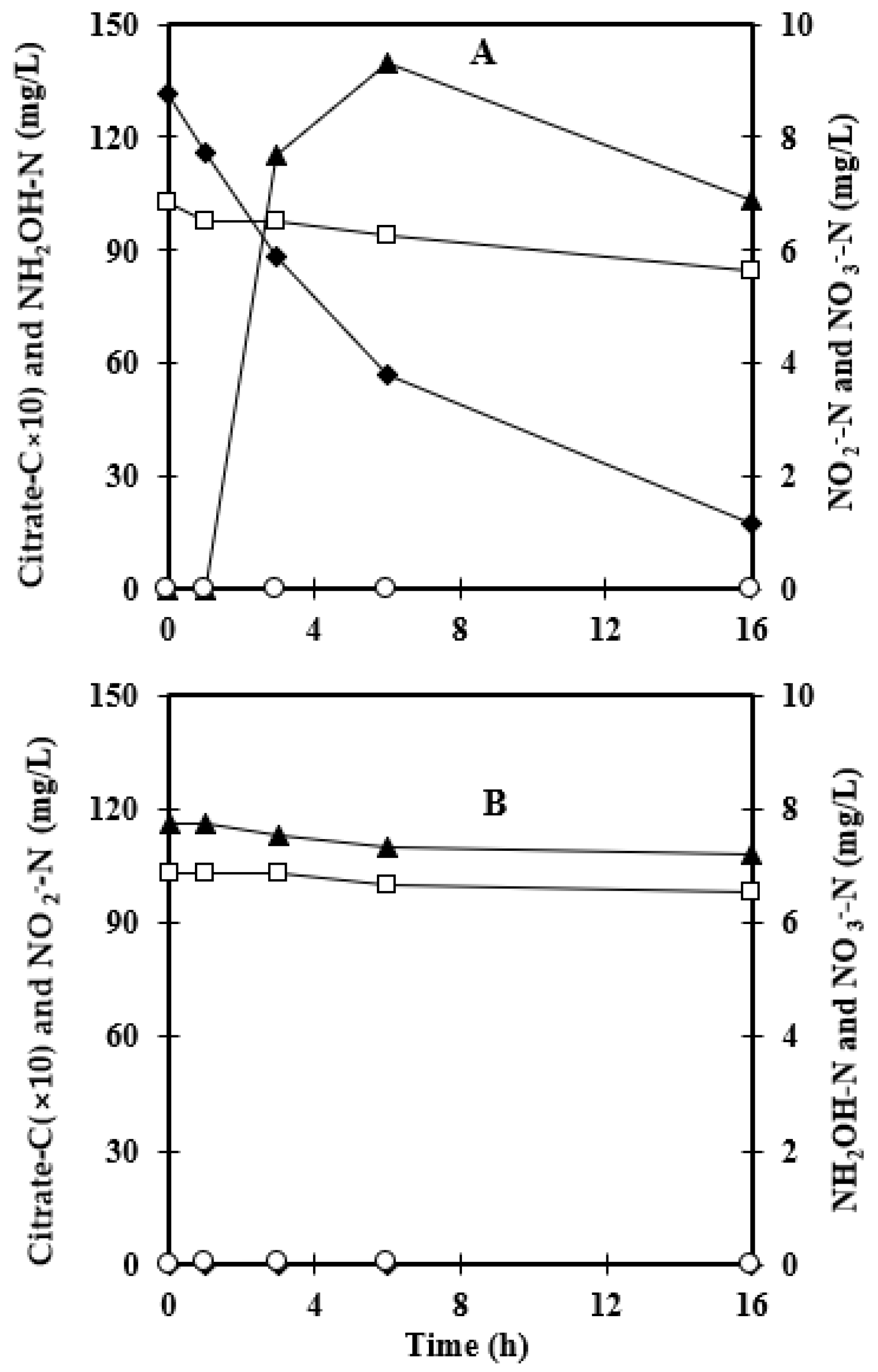
Figure 5.
Removal of nitrogen spieces (with hydroxylamine (A) and nitrite (B) instead of ammonium) and carbon consumption in batch reactors with No. 4 culture. Hydroxylamine (◆), nitrite (▲), nitrate (○), and citrate (□).
Figure 5.
Removal of nitrogen spieces (with hydroxylamine (A) and nitrite (B) instead of ammonium) and carbon consumption in batch reactors with No. 4 culture. Hydroxylamine (◆), nitrite (▲), nitrate (○), and citrate (□).
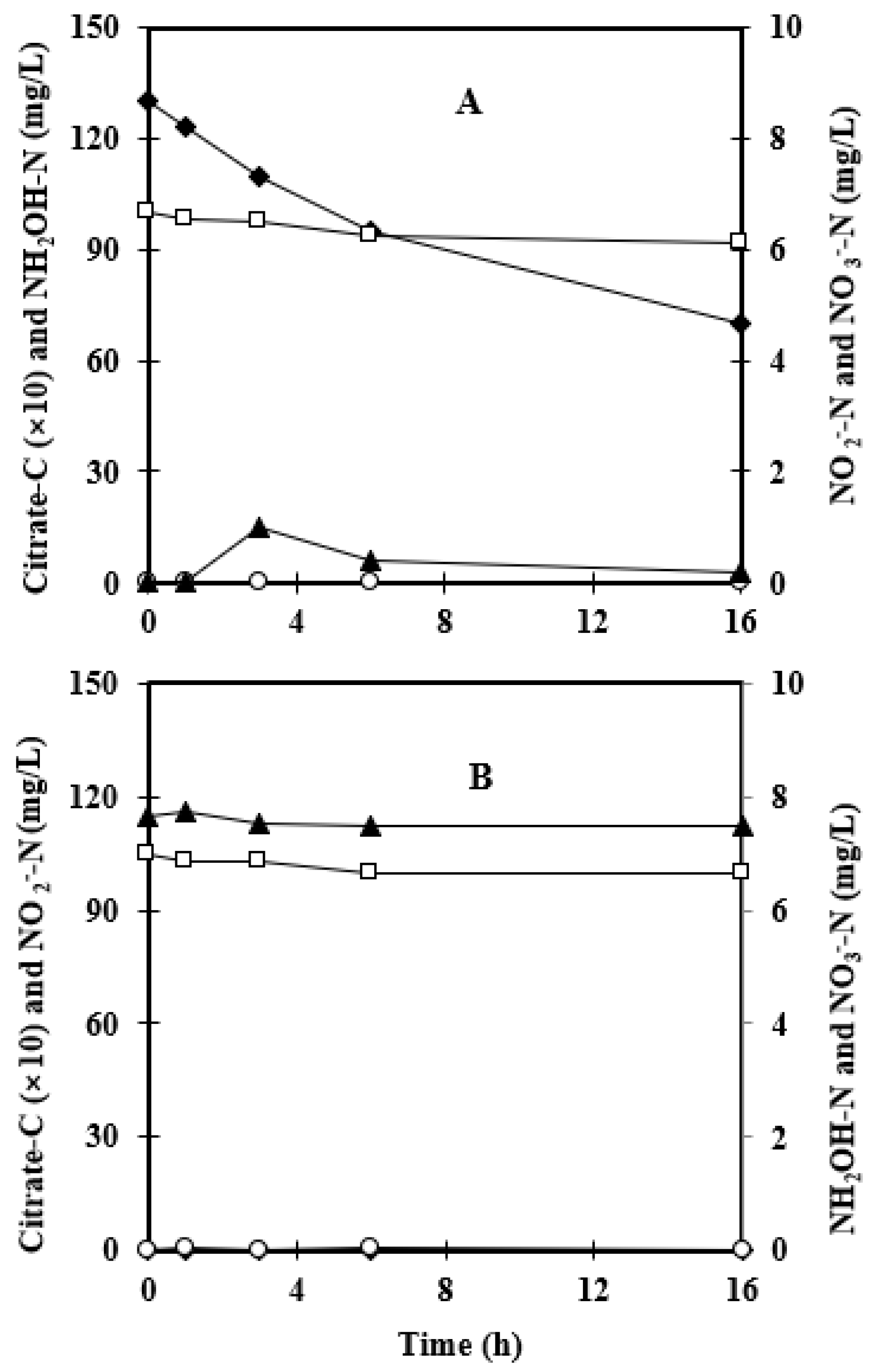
Figure 6.
Spectra of nitrite reducing activity (NIR activity). Control (no addition of enzyme), F (separated foam from aerated batch cultivation by No. 4), Cb (No. 4 culture broth).
Figure 6.
Spectra of nitrite reducing activity (NIR activity). Control (no addition of enzyme), F (separated foam from aerated batch cultivation by No. 4), Cb (No. 4 culture broth).

Figure 7.
Ammonia stripping with pH change during the aerated batch experiments (simulated based on data presented in Table 4).
Figure 7.
Ammonia stripping with pH change during the aerated batch experiments (simulated based on data presented in Table 4).

Figure 8.
Nitrogen balance and removal pathway in the heterotrophic nitrification-aerobic denitrification by A. faecalis No. 4.
Figure 8.
Nitrogen balance and removal pathway in the heterotrophic nitrification-aerobic denitrification by A. faecalis No. 4.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nitrogen balance in aerated batch experiments of A. faecalis No. 4 with and without foam recycling (Initial pH: 7, the time elasped: 110 h).
Table 1.
Nitrogen balance in aerated batch experiments of A. faecalis No. 4 with and without foam recycling (Initial pH: 7, the time elasped: 110 h).
| Without Foam Recycling | Foam Recycling | |
|---|---|---|
| Initial NH4+-N (mg/L) | 1037 ± 22 | 1122 ± 5.5 |
| Removed N (mg/L) | 1037 ± 22 | 1122 ± 5.5 |
| Intracellular N (mg/L) | 481 ± 9.8 | 509 ± 13 |
| Nitrification products (mg-N/L) | 101 ± 8.7 | 30 ± 4.3 |
| Stripped N (mg/L) | 46 ± 3.6 | 41 ± 4.3 |
| Denitrification ratio (%) * | 39 ± 1.1 | 48 ± 0.4 |
| Maximum ammonium removal rate (mg-N/L·h) | 26 ± 0.9 | 25 ± 1.2 |
*: Denitrification ratio is percentage of the removed ammonium based on initial ammonium concentration.
Table 2.
Hydroxylamine oxidoreductase activities (HAO) in separated foam and No. 4 culture media.
| Foam | Culture of No. 4 | |
|---|---|---|
| Cell number (cfu/mL) | 2.1 ± 2.1 × 1010 | 4.0 ± 1.2 × 109 |
| Activity (U/mL) * | 34.3 ± 3.2 | 1.25 ± 0.1 |
| Specific Activity (U/cfu) | 1.5 × 10−9 | 3.3 × 10−10 |
* One unit (U) is the amount of the enzyme, which reduce 1 μM-K3Fe(C/N)6/min.
Table 3.
Nitrogen balance in different airflow rate and initial ammonium concentrations.
| Airflow Rate (L/min) | Initial Ammonium Level (mg-N/L) | |||
|---|---|---|---|---|
| 3000 | 1000 | 250 | ||
| Initial N (mg/L) Removed N (mg/L) (%) Intracellular N (mg/L) (%) * Nitrification N (mg/L) (%) * Stripped N (mg/L) (%) * Denitrification ratio (%) * Maximum ammonium removal rate (mg-N/L·h) | 0.2 | 2965 | 1167 | 217 |
| 990 (33) | 617 (53) | 217 (100) | ||
| 378 (38) | 318 (52) | 110 (51) | ||
| 19 (2) | 14 (2) | 13 (6) | ||
| 289 (29) | 40 (6) | 2.5 (1) | ||
| 31 | 40 | 42 | ||
| 13 | 11 | 11 | ||
| Initial N (mg/L) Removed N (mg/L) (%) Intracellular N (mg/L) (%) * Nitrification N (mg/L (%) * Stripped N (mg/L) (%) * Denitrification ratio (%) * Maximum ammonium removal rate (mg-N/L·h) | 0.5 | 2975 | 1190 | 218 |
| 2697 (91) | 1063 (89) | 218 (100) | ||
| 546 (20) | 397 (37) | 124 (57) | ||
| 39 (2) | 29 (3) | 21 (10) | ||
| 333 (12) | 59 (6) | 5 (2) | ||
| 66 | 54 | 31 | ||
| 28 | 19 | 13 | ||
| Initial N (mg/L) Removed N (mg/L) (%) Intracellular N (mg/L) (%) * Nitrification N (mg/L) (%) * Stripped N (mg/L) (%) * Denitrification ratio (%) * Maximum ammonium removal rate (mg-N/L·h) | 0.8 | 2967 | 1177 | 225 |
| 2967 (100) | 1104 (94) | 225 (100) | ||
| 668 (23) | 404 (37) | 128 (57) | ||
| 56 (2) | 38 (3) | 24 (11) | ||
| 354 (12) | 72 (6) | 4 (2) | ||
| 64 | 54 | 31 | ||
| 39 | 21 | 15 | ||
* Percentages based on removed ammonium. The cultivation times in 3000 ppm, 1000 ppm, and 250 ppm of initial ammonium concentrations (mg-N/L) were 137 h, 80 h and 34 h, respectively.
Table 4.
Nitrogen balance at different initial pH-conditions.
| pH 6 | pH 7 | pH 8 | |
|---|---|---|---|
| Initial NH4+-N (mg/L) | 1186 | 1122 | 1169 |
| Removed N (mg/L) | 1165 | 1122 | 1159 |
| Intracellular N (mg/L) | 424 | 509 | 521 |
| Nitrification N (mg/L) | 25 | 30 | 21 |
| Stripped N (mg/L) | 79 | 41 | 170 |
| Denitrification ratio (%) * | 55 | 48 | 39 |
| Maximum ammonium removal rate (mg-N/L) | 20 | 25 | 15 |
* Denitrification ratio is percentage against removed ammonium and there were no changes at pH 5.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, K.; Joo, H.-S. The Effect of Foam-Recycle on Ammonium Removal by Aerobic Denitrification Using Alcaligenes faecalis No. 4. Environments 2023, 10, 184. https://doi.org/10.3390/environments10100184
AMA Style
Lee K, Joo H-S. The Effect of Foam-Recycle on Ammonium Removal by Aerobic Denitrification Using Alcaligenes faecalis No. 4. Environments. 2023; 10(10):184. https://doi.org/10.3390/environments10100184
Chicago/Turabian StyleLee, Kwanyong, and Hung-Soo Joo. 2023. "The Effect of Foam-Recycle on Ammonium Removal by Aerobic Denitrification Using Alcaligenes faecalis No. 4" Environments 10, no. 10: 184. https://doi.org/10.3390/environments10100184
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.