
Inoculation with Biofilm of Bacillus subtilis Is a Safe and Sustainable Alternative to Promote Tomato (Solanum lycopersicum) Growth
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biofilm and Planktonic State Characterization and Inoculant Preparation
2.1.1. Vegetative Cells and Spores in Biofilm and Planktonic State
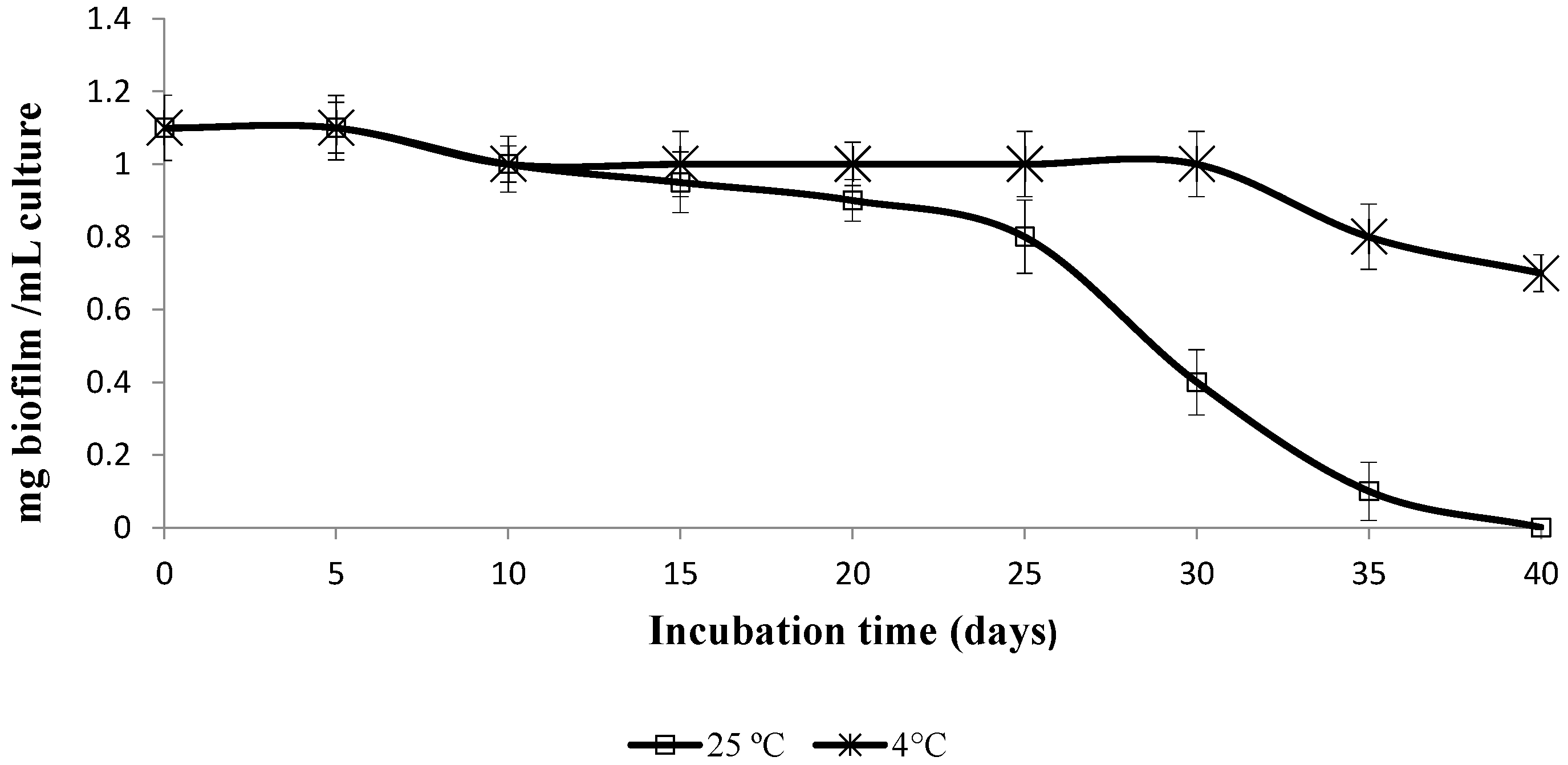
2.1.2. Biofilm Stability
2.1.3. Inoculant Preparation
2.2. Experimental Site and Design
2.2.1. Seed Germination Assays
2.2.2. Greenhouse Assays
2.3. Agronomic and Production Variables
2.4. Chlorophyll Estimations
2.5. Bacterial Counts in Root Extracts of Solanum lycopersicum var. Río Grande
2.6. Statistical Analysis
3. Results
3.1. Content of Vegetative Cells and Spores in the Planktonic State and in Biofilm
3.2. Biofilm Stability
3.2.1. Effect of Carbon Source
3.2.2. Effect of Temperature
3.3. Effects of Inoculation Form on Germination and Growth of Two Varieties of S. lycopersicum
3.3.1. Effects on Germination Percent of Platense and Río Grande Varieties
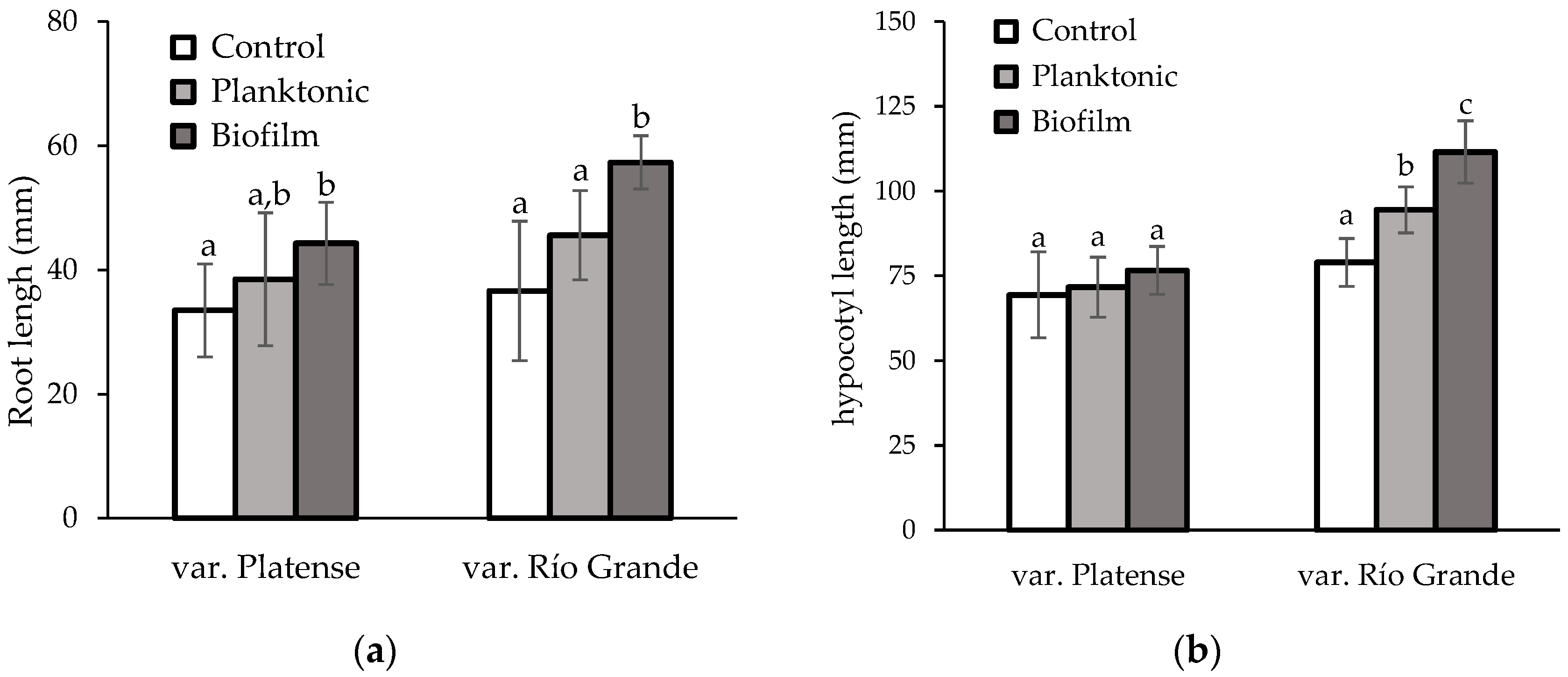
3.3.2. Effects on Seedlings of Platense and Río Grande Varieties after Germination
3.3.3. Effects on Biomass of Platense and Río Grande Varieties at the End of Vegetative Growth Stage
3.4. Effects on Production and Quality of Río Grande Variety at Harvest Time
3.4.1. Effects on Plant Biomass and Plant Height
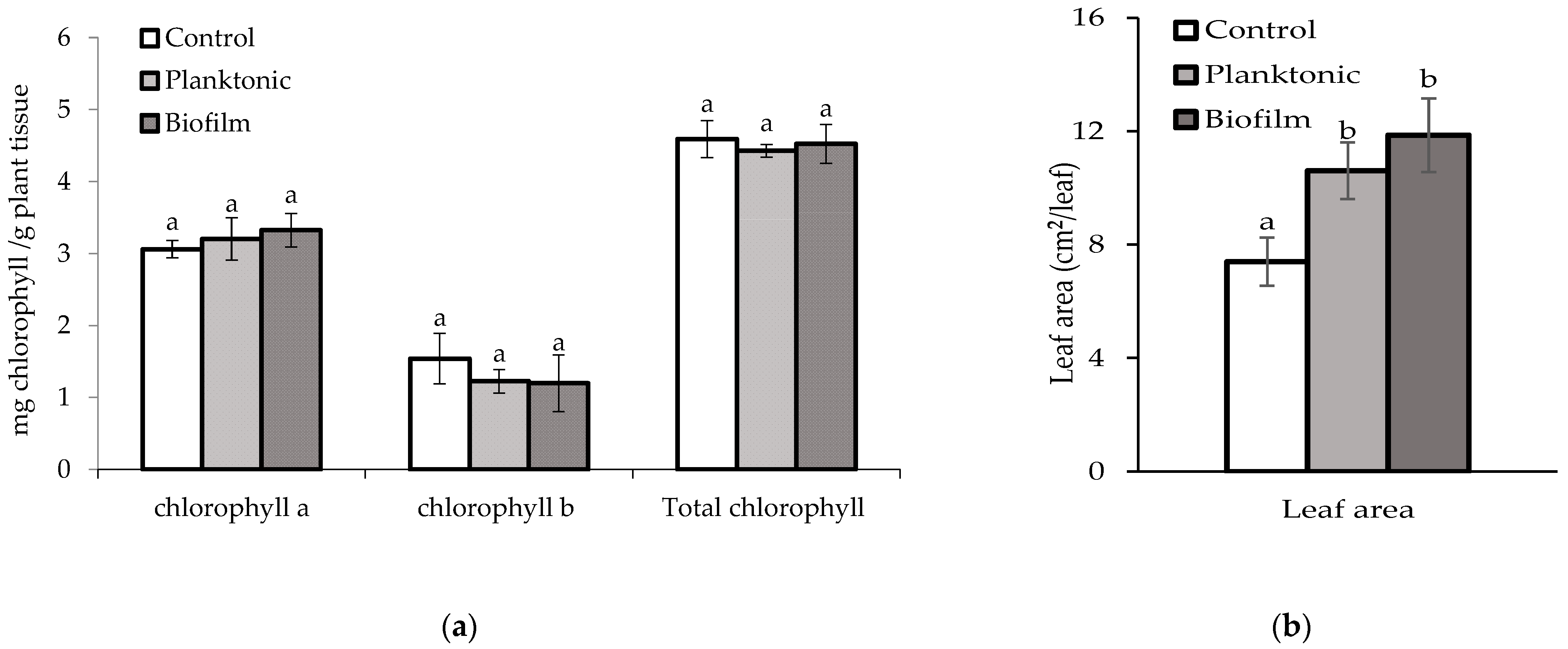
3.4.2. Quantification of Chlorophyll and Leaf Area
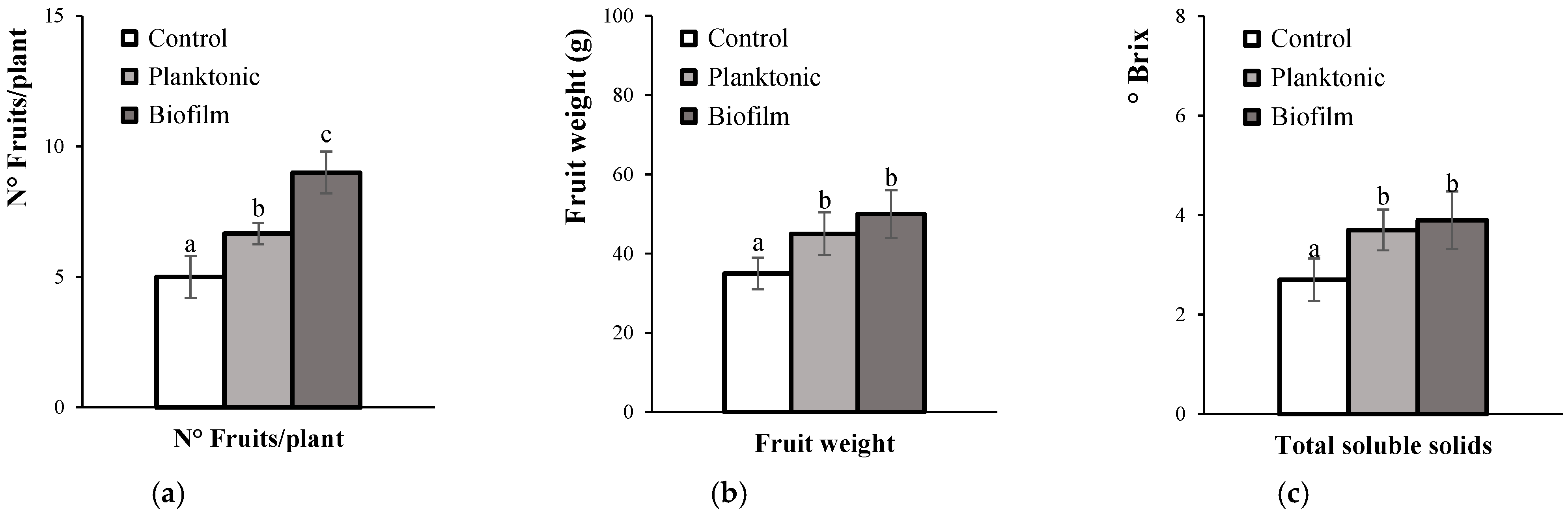
3.4.3. Fruit Productivity and Quality
3.5. Bacterian Endophytism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Corato, U. Soil Microbiota Manipulation and Its Role in Suppressing Soil-Borne Plant Pathogens in Organic Farming Systems under the Light of Microbiome-Assisted Strategies. Chem. Biol. Technol. Agric. 2020, 7, 17. [Google Scholar] [CrossRef]
- Cao, M.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Ma, Y. Optimistic Contributions of Plant Growth-Promoting Bacteria for Sustainable Agriculture and Climate Stress Alleviation. Environ. Res. 2023, 217, 114924. [Google Scholar] [CrossRef]
- Ahmed, H.M.; Gad, S.B.; El-Sherif, A.G.; El-Hadidy, E.M. Efficacy of Five Biopesticides for the Management of Root-Knot Nematode, Meloidogyne Incognita Infecting Pepper (Capsicum anuum L.) Plants. Egypt. J. Agronematol. 2022, 21, 23–33. [Google Scholar] [CrossRef]
- FAO; IFAD; WFP. The State of Food Insecurity in the World 2013; FAO: Rome, Italy, 2013. [Google Scholar]
- FAO; GEF. Partners in Action Feeding People, Protecting the Planet; FAO: Rome, Italy, 2018. [Google Scholar]
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A Meta-Analysis of Projected Global Food Demand and Population at Risk of Hunger for the Period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Massa, F.; Defez, R.; Bianco, C. Exploitation of Plant Growth Promoting Bacteria for Sustainable Agriculture: Hierarchical Approach to Link Laboratory and Field Experiments. Microorganisms 2022, 10, 865. [Google Scholar] [CrossRef]
- Sammauria, R.; Kumawat, S.; Kumawat, P.; Singh, J.; Jatwa, T.K. Microbial Inoculants: Potential Tool for Sustainability of Agricultural Production Systems. Arch. Microbiol. 2020, 202, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; Enshasy, H. El Plant Growth Promoting Rhizobacteria (Pgpr) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Aydi Ben Abdallah, R.; Jabnoun-Khiareddine, H.; Regaieg, H.; Daami-Remadi, M. Effect of Indigenous Bio-Inoculants and Commercial Biological Inputs on Soil Microbial Population, Soil Health Dynamics and Pepper (Capsicum anuum L.) Production. Arch. Agron. Soil Sci. 2023, 69, 3484–3501. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef]
- Prasad, M.; Srinivasan, R.; Chaudhary, M.; Choudhary, M.; Jat, L.K. Plant Growth Promoting Rhizobacteria (PGPR) for Sustainable Agriculture: Perspectives and Challenges. In PGPR Amelioration in Sustainable Agriculture; Woodhead Publishing: Sawston, UK, 2019; pp. 129–157. [Google Scholar]
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight Into the Role of PGPR in Sustainable Agriculture and Environment. Front. Sustain. Food Syst. 2021, 5, 667150. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Gomez-Ramirez, L.F.; Uribe-Velez, D. Phosphorus Solubilizing and Mineralizing Bacillus spp. Contribute to Rice Growth Promotion Using Soil Amended with Rice Straw. Curr. Microbiol. 2021, 78, 932–943. [Google Scholar] [CrossRef]
- Pérez Vargas, J.; Vigueras Carmona, S.E.; Zamudio Moreno, E.; Rivera Casado, N.A.; Calva Calva, G. Bioremediation of Soils from Oil Spill Impacted Sites Using Bioaugmentation with Biosurfactants Producing, Native, Free-Living Nitrogen Fixing Bacteria. Rev. Int. Contam. Ambient. 2017, 33, 105–114. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial Plant Biostimulants: A Sustainable Way towards Improving Growth, Productivity, and Health of Crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Das, P.P.; Singh, K.R.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.A.; Kumar, A.; Singh, J. Plant-Soil-Microbes: A Tripartite Interaction for Nutrient Acquisition and Better Plant Growth for Sustainable Agricultural Practices. Environ. Res. 2022, 214, 113821. [Google Scholar] [CrossRef]
- Ortiz, A.; Sansinenea, E. The Role of Beneficial Microorganisms in Soil Quality and Plant Health. Sustainability 2022, 14, 5358. [Google Scholar] [CrossRef]
- Sarti, G.C.; Galelli, M.E.; Arreghini, S.; Cristóbal-Miguez, J.A.E.; Curá, J.A.; Paz-González, A. Inoculation with Biofilm of Bacillus subtilis Promotes the Growth of Lactuca Sativa. Sustainability 2023, 15, 15406. [Google Scholar] [CrossRef]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in Agriculture: A Sustainable Approach to Increasing Climate Change Resilience. Front. Sustain. Food Syst. 2021, 5, 667546. [Google Scholar] [CrossRef]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef]
- Hobley, L.; Harkins, C.; MacPhee, C.E.; Stanley-Wall, N.R. Giving Structure to the Biofilm Matrix: An Overview of Individual Strategies and Emerging Common Themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef]
- Penha, R.O.; Vandenberghe, L.P.S.; Faulds, C.; Soccol, V.T.; Soccol, C.R. Bacillus Lipopeptides as Powerful Pest Control Agents for a More Sustainable and Healthy Agriculture: Recent Studies and Innovations. Planta 2020, 251, 70. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Angelini, L.L.; Chai, Y. Bacillus subtilis Cell Differentiation, Biofilm Formation and Environmental Prevalence. Microorganisms 2022, 10, 1108. [Google Scholar] [CrossRef]
- Galelli, M.E.; Sarti, G.C.; Miyazaki, S.S. Lactuca sativa Biofertilization Using Biofilm from Bacillus with PGPR Activity. J. Appl. Hortic. 2015, 17, 186–191. [Google Scholar] [CrossRef]
- Sarti, G.; Curá, A. Optimización de Las Condiciones de Cultivo Para El Desarrollo de Una Biopelícula Bacteriana y Su Aplicación Como Biofertilizante En Solanum lycopersicum L. Var. Río Grande. Rev. Prot. Veg. 2019, 34, 1–19. [Google Scholar]
- FAO. World Food and Agriculture—Statistical Yearbook 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- Tripodi, P.; Kumar, S. The Capsicum Crop: An Introduction. In Compendium of Plant Genomes; Ramchiary, N., Kole, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 1–8. ISBN 9783319972176. [Google Scholar]
- Rosa-Martínez, E.; García-Martínez, M.D.; Adalid-Martínez, A.M.; Pereira-Dias, L.; Casanova, C.; Soler, E.; Figàs, M.R.; Raigón, M.D.; Plazas, M.; Soler, S.; et al. Fruit Composition Profile of Pepper, Tomato and Eggplant Varieties Grown under Uniform Conditions. Food Res. Int. 2021, 147, 110531. [Google Scholar] [CrossRef]
- Imran, M.; Ghorat, F.; Ul-haq, I.; Ur-rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef]
- Paladino, I.; Sokolowski, A.C.; Prack Mec Cormick, B.; Enrique, W.J.; Rodríguez, H.; Navas, M. Soil Quality Problems Associated with Horticulture in the Southern Urban and Peri-Urban Area of Buenos Aires, Argentina. In Urban Horticulture—Necessity of the Future; Chapters: Toronto, ON, Canada, 2020; pp. 1–21. ISBN 0000957720. [Google Scholar]
- Giuffré, L.; Ratto, S.; Marbán, L.; Schonwald, J.; Romaniuk, R. Heavy Metals Risk in Urban Agriculture. Cienc. Suelo 2005, 23, 101–106. [Google Scholar]
- Scarpatti, N. Producción de Tomate en Argentina: Evolución del Cultivo hasta la Temporada 2021/22 Febrero; Informe Técnico de la Secretaría de Agricultura, Ganadería y Pesca, Ministerio de Economía: Buenos Aires, Argentina, 2023; pp. 1–19. [Google Scholar]
- Luna Martínez, L.; Peniche, R.A.M.; Iturriaga, M.H.; Medrano, S.M.A.; Pacheco Aguilar, J.R. Characterization of Rhizobacteria Isolated from Tomato and Their Effect on Tomato and Bell Pepper Growth. Rev. Fitotec. Mex. 2013, 36, 63–69. [Google Scholar]
- Carrillo-Castañeda, G.; Juárez Muñoz, J.; Ruiz Landa, D.; Muller García, R. Aumento Del Rendimiento de Tomate (Lycopersicon esculentum Mill) Cuando La Raiz Se Desarrolla Colonizada Por Microorganismos. Biotecnol. Appl. 2000, 17, 171–176. [Google Scholar]
- Carillo-Sosa, Y.; Terry-Alfonso, E.; Ruiz-Padrón, J. Effect of a Microbial Inoculum on the Growth of Tomato Plants (Solanum lycopersicum L.). Cultiv. Trop. 2020, 41, e01. [Google Scholar]
- Balderas-Ruíz, K.A.; Gómez-Guerrero, C.I.; Trujillo-Roldán, M.A.; Valdez-Cruz, N.A.; Aranda-Ocampo, S.; Juárez, A.M.; Leyva, E.; Galindo, E.; Serrano-Carreón, L. Bacillus velezensis 83 Increases Productivity and Quality of Tomato (Solanum lycopersicum L.): Pre and Postharvest Assessment. Curr. Res. Microb. Sci. 2021, 2, 100076. [Google Scholar] [CrossRef]
- Philipp, G. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994. [Google Scholar]
- Frioni, L. Ecología Microbiana Del Suelo; UDELAR, Ed.; Universidad de la República: Montevideo, Uruguay, 1990. [Google Scholar]
- Alfonso Martí, L. Efecto de La Salinidad y de La Temperatura En La Germinación de Semillas de Limonium mansanetianum. Ph.D. Thesis, Universidad Politécnica de Valencia, Valencia, Spain, 2010. [Google Scholar]
- Araya, E.; Gómez, L.; Hidalgo, N.; Valverde, R. Efecto de La Luz y Del Ácido Gibérelico Sobre La Germinación in Vitro de Jaul (Alnus acuminata). Agron. Costarric. 2000, 24, 75–80. [Google Scholar]
- Sarti, G.C. New Perspectives for Urban Horticulture: Evaluation of a Microbial Biofilm as Plant Growth Promoter; University of A Coruña: Coruña, Spain, 2021. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis HHS Public Access. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction Coefficients of Chlorophyll a and b in N,N—Dimethylformamide and 80% Acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef]
- Dobereiner, J.; Urquiaga, S.; Boddey, R.M. Alternatives for Nitrogen Nutrition of Crops in Tropical Agriculture. Fertil. Res. 1995, 42, 339–346. [Google Scholar] [CrossRef]
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. The Potential of Bacilli Rhizobacteria for Sustainable Crop Production and Environmental Sustainability. Microbiol. Res. 2019, 219, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Zahir, Z.A.; Iqbal, M.; Mehmood, A.; Javed, A.; Hussain, A.; Bushra; Ahmad, M. Efficacy of Rhizobacterial Exopolysaccharides in Improving Plant Growth, Physiology, and Soil Properties. Environ. Monit. Assess. 2021, 193, 515. [Google Scholar] [CrossRef]
- Kaniz, F.; Zheng, W.; Bais, H.; Jin, Y. Plant Growth-Promoting Rhizobacteria Mediate Soil Hydro-Physical Properties: An Investigation with Bacillus subtilis and Its Mutants. Vadose Zone J. 2023, 22, e20274. [Google Scholar] [CrossRef]
- Ajijah, N.; Fiodor, A.; Pandey, A.K.; Rana, A. Plant Growth-Promoting Bacteria (PGPB) with Biofilm-Forming Ability: A Multifaceted Agent for Sustainable Agriculture. Diversity 2023, 15, 112. [Google Scholar] [CrossRef]
- Grimbergen, A.J.; Siebring, J.; Solopova, A.; Kuipers, O.P. Microbial Bet-Hedging: The Power of Being Different. Curr. Opin. Microbiol. 2015, 25, 67–72. [Google Scholar] [CrossRef]
- Kovács, Á.T.; Stanley-Wall, N.R. Biofilm Dispersal for Spore Release in Bacillus subtilis. J. Bacteriol. 2021, 203, e00192-21. [Google Scholar] [CrossRef]
- Branda, S.S.; González-Pastor, J.E.; Ben-Yehuda, S.; Losick, R.; Kolter, R. Fruiting Body Formation by Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2001, 98, 11621–11626. [Google Scholar] [CrossRef]
- Lemon Kolter, R.; Earl, A.M.; Vlamakis, H.C.; Aguilar, C.; Kolter, R. Biofilm Development with Emphasis on Bacillus subtilis. In Bacterial Biofilms; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2008; Volume 322, pp. 1–16. [Google Scholar]
- Nishikawa, M.; Kobayashi, K. Calcium Prevents Biofilm Dispersion in Bacillus subtilis. J. Bacteriol. 2021, 203, 10–1128. [Google Scholar] [CrossRef]
- Wang, Z.R.; Sheng, J.P.; Tian, X.L.; Wu, T.T.; Liu, W.Z.; Shen, L. Optimization of the Production of Exopolysaccharides by Bacillus thuringiensis 27 in Sand Biological Soil Crusts and Its Bioflocculant Activity. Afr. J. Microbiol. Res. 2011, 5, 2359–2366. [Google Scholar]
- Pham, P.L.; Dupont, I.; Roy, D.; Lapointe, G.; Cerning, J. Production of Exopolysaccharide by Lactobacillus rhamnosus R and Analysis of Its Enzymatic Degradation during Prolonged Fermentation. Appl. Environ. Microbiol. 2000, 66, 2302–2310. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Arellano, C.; Zúñiga, D. Efecto de Diferentes Bacterias Aisladas de Rizósfera de Caesalpina Spinosa En La Germinación de Diferentes Especies Vegetales Culivados. Zonas Árid. 2008, 12, 137–153. [Google Scholar]
- Dakora, F.D.; Phillips, D.A. Root Exudates as Mediators of Mineral Acquisition in Low-Nutrient Environments. Plant Soil 2002, 245, 35–47. [Google Scholar] [CrossRef]
- González, F.H.; Fuentes, M.N. Mecanismo de Acción de Cinco Microorganismos Promotores de Crecimiento Vegetal. Rev. Cienc. Agríc. 2017, 34, 17–31. [Google Scholar] [CrossRef]
- ISTA. Seed Health Testing. In International Rules for Seed Testing 2024; ISTA: Wallisellen, Switzerland, 2024; p. 345. ISBN 9783906549699. [Google Scholar]
- Barreto, D.; Valero, N.; Muñoz, A.; Peralta, A. Efecto de Microorganismos Rizosféricos Sobre Germinación y Crecimiento Temprano de Anacardium Excelsum. Zonas Árid. 2007, 11, 240–250. [Google Scholar]
- Yildirim, E.; Karlidag, H.; Turan, M.; Dursun, A.; Goktepe, F. Growth, Nutrient Uptake, and Yield Promotion of Broccoli by Plant Growth Promoting Rhizobacteria With. Hort Sci. 2011, 46, 932–936. [Google Scholar] [CrossRef]
- Sánchez López, D.B.; Gómez-Vargas, R.M.; Garrido Rubiano, M.F.; Bonilla Buitrago, R.R. Inoculación Con Bacterias Promotoras de Crecimiento Vegetal En Tomate Bajo Condiciones de Invernadero. Rev. Mex. Cienc. Agríc. 2018, 3, 1401–1415. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Yildirim, E.; Güneş, A.; Karagöz, K.; Kotan, R.; Dursun, A. Plant Growth-Promoting Rhizobacteria Improved Growth, Nutrient, and Hormone Content of Cabbage (Brassica oleracea) Seedlings. Turk. J. Agric. For. 2014, 38, 327–333. [Google Scholar] [CrossRef]
- Widnyana, K.I. PGPR (Plant Growth Promoting Rizobacteria) Benefits in Spurring Germination, Growth and Increase the Yield of Tomato Plants. In Recent Advances in Tomato Breeding and Production; IntechOpen: London, UK, 2018; pp. 17–25. [Google Scholar]
- Chakraborty, A.P. Carrier Based Bioformulations of PGPR-Characteristics, Shelf Life and Application in Improving Health Status of Crop Plants—A Mini Review. Int. J. Res. Rev. 2020, 7, 88–98. [Google Scholar]
- Qasem, J.R. Weed Seed Dormancy: The Ecophysiology and Survival Strategies. In Seed Dormancy and Germination; IntechOpen: London, UK, 2019; p. 13. [Google Scholar]
- Costales, D.; Martínez, L.; Núñez, M. Efecto Del Tratamiento De Semillas Con Una Mezcla De Oligogalacturónidos Sobre El Crecimiento De Plántulas De Tomate (Lycopersicon esculentum Mill). Cultiv. Trop. 2007, 28, 85–91. [Google Scholar]
- Monge-Pérez, J.E.; Loría-Coto, M. Determinación de Criterios de Selección Para El Rendimiento de Tomate (Solanum lycopersicum L.) Cultivado Bajo Invernadero. Av. Investig. Agropecu. 2021, 25, 7–19. [Google Scholar]
- Mendoza Tafolla, R.; Juarez Lopez, P.; Ontiveros Capurata, R.; Aliaga, I.; Guillén, D.; Villegas, O.; Chávez, A. Estimación de la concentración de clorofila, nitrógeno y biomasa en arúgula (Eruca sativa Mill.) mediante mediciones portátiles no destructivas Rodrigo. Bioagro 2022, 34, 151–162. [Google Scholar] [CrossRef]
- Jacobs, M.J.; Bugbee, W.M.; Gabrielson, D.A. Enumeration, Location, and Characterization of Endophytic Bacteria within Sugar Beet Roots. Can. J. Bot. 1985, 63, 1262–1265. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial Endophytes: Recent Developments and Applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef]
- Cochard, B.; Giroud, B.; Crovadore, J.; Chablais, R.; Arminjon, L.; Lefort, F. Endophytic PGPR from Tomato Roots: Isolation, In Vitro Characterization and In Vivo Evaluation of Treated Tomatoes (Solanum lycopersicum L.). Microorganisms 2022, 10, 765. [Google Scholar] [CrossRef]
- Ricci, E.; Schwinghamer, T.; Fan, D.; Smith, D.L.; Gravel, V. Growth Promotion of Greenhouse Tomatoes with Pseudomonas sp. and Bacillus sp. Biofilms and Planktonic Cells. Appl. Soil Ecol. 2019, 138, 61–68. [Google Scholar] [CrossRef]
- Domínguez-González, K.G.; Robledo-Medrano, J.J.; Valdez-Alarcón, J.J.; Hernández-Cristobal, O.; Martínez-Flores, H.E.; Cerna-Cortés, J.F.; Garnica-Romo, M.G.; Cortés-Martínez, R. Streptomyces Spp. Biofilmed Solid Inoculant Improves Microbial Survival and Plant-Growth Efficiency of Triticum Aestivum. Appl. Sci. 2022, 12, 11425. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination (%) 5 Days | Germination (%) 10 Days | |||||
|---|---|---|---|---|---|---|
| Tomato varieties | Control | Planktonic | Biofilm | Control | Planktonic | Biofilm |
| Platense | 74.2 ± 1.3 b | 82.4 ± 0.8 a | 27.0 ± 0.9 c | 80.3 ± 2.1 b | 87.0 ± 0.9 a | 90.1 ± 0.9 a |
| Río Grande | 88.0 ± 1.1 a | 90.0 ± 1.5 a | 30.0 ± 1.1 b | 90.4 ± 1.4 b | 95.2 ± 1.3 a | 96.0 ± 0.8 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarti, G.C.; Galelli, M.E.; Cristóbal-Miguez, J.A.E.; Cárdenas-Aguiar, E.; Chudil, H.D.; García, A.R.; Paz-González, A. Inoculation with Biofilm of Bacillus subtilis Is a Safe and Sustainable Alternative to Promote Tomato (Solanum lycopersicum) Growth. Environments 2024, 11, 54. https://doi.org/10.3390/environments11030054
Sarti GC, Galelli ME, Cristóbal-Miguez JAE, Cárdenas-Aguiar E, Chudil HD, García AR, Paz-González A. Inoculation with Biofilm of Bacillus subtilis Is a Safe and Sustainable Alternative to Promote Tomato (Solanum lycopersicum) Growth. Environments. 2024; 11(3):54. https://doi.org/10.3390/environments11030054
Chicago/Turabian StyleSarti, Gabriela Cristina, Mirta Esther Galelli, Josefina Ana Eva Cristóbal-Miguez, Eliana Cárdenas-Aguiar, Hugo Daniel Chudil, Ana Rosa García, and Antonio Paz-González. 2024. "Inoculation with Biofilm of Bacillus subtilis Is a Safe and Sustainable Alternative to Promote Tomato (Solanum lycopersicum) Growth" Environments 11, no. 3: 54. https://doi.org/10.3390/environments11030054