
Evaluation of the Effects of Environmental Factors on Seasonal Variations in Fish Diversity on a Coastal Island in Western Japan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Environmental DNA (eDNA) Analysis and Species Analysis
2.4. Bioinformatic Analysis
2.5. Statistical Analysis
2.5.1. Nonparametric Analyses
2.5.2. Multivariate Analysis
3. Results
3.1. Seasonal Variations in Water Temperature, Salinity, and Nutrients
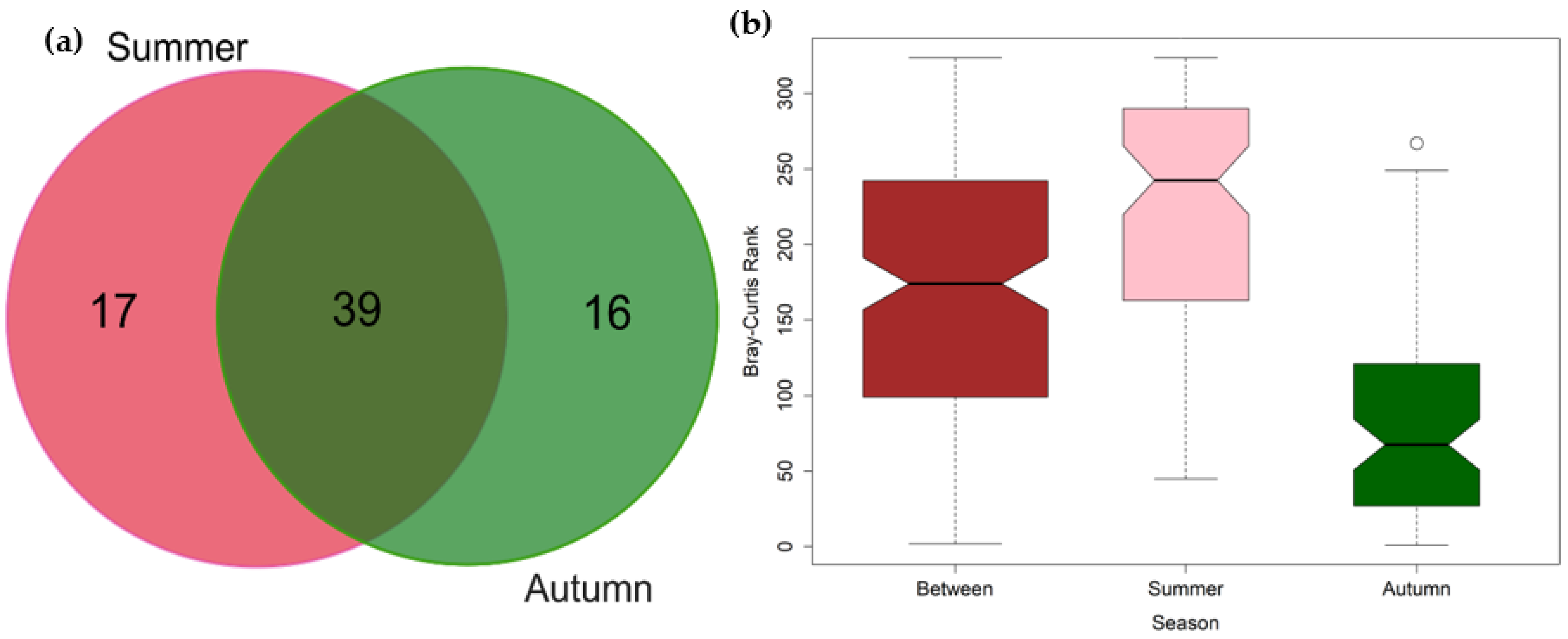
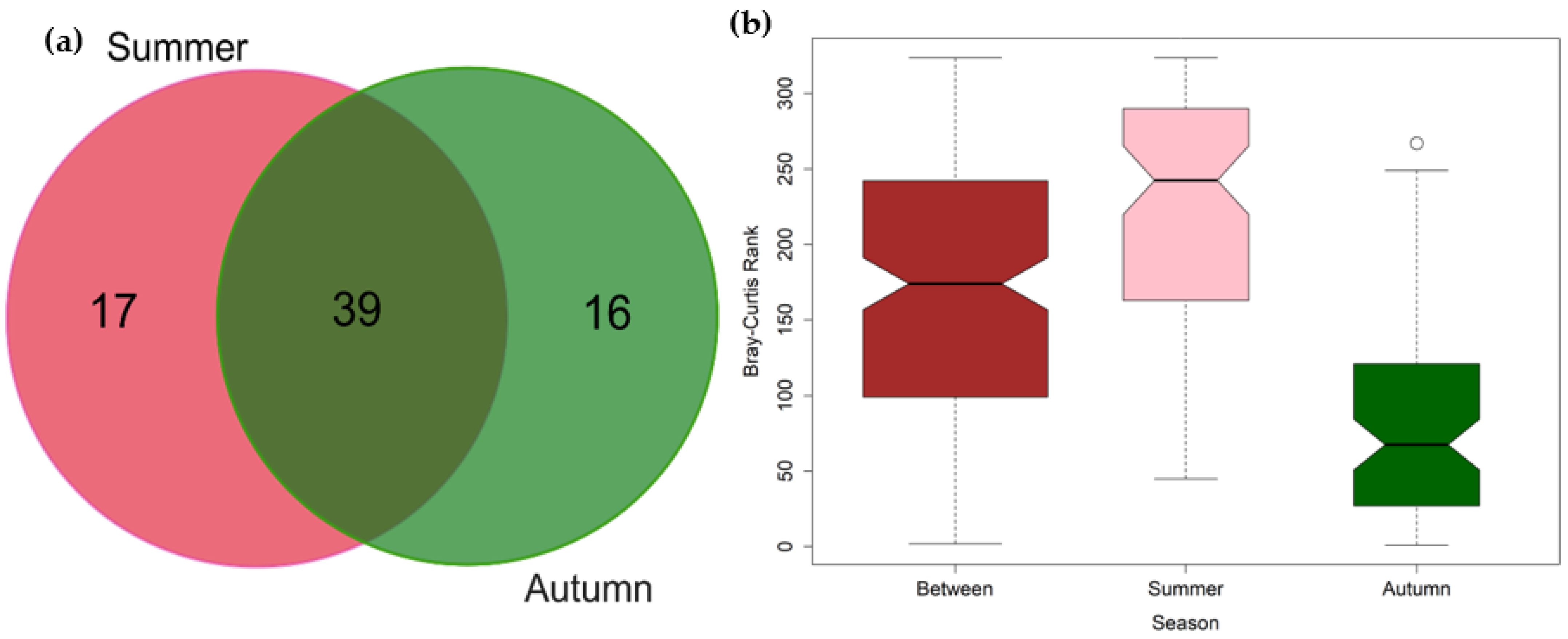
3.2. Seasonal Variations in the Composition of Fish Communities
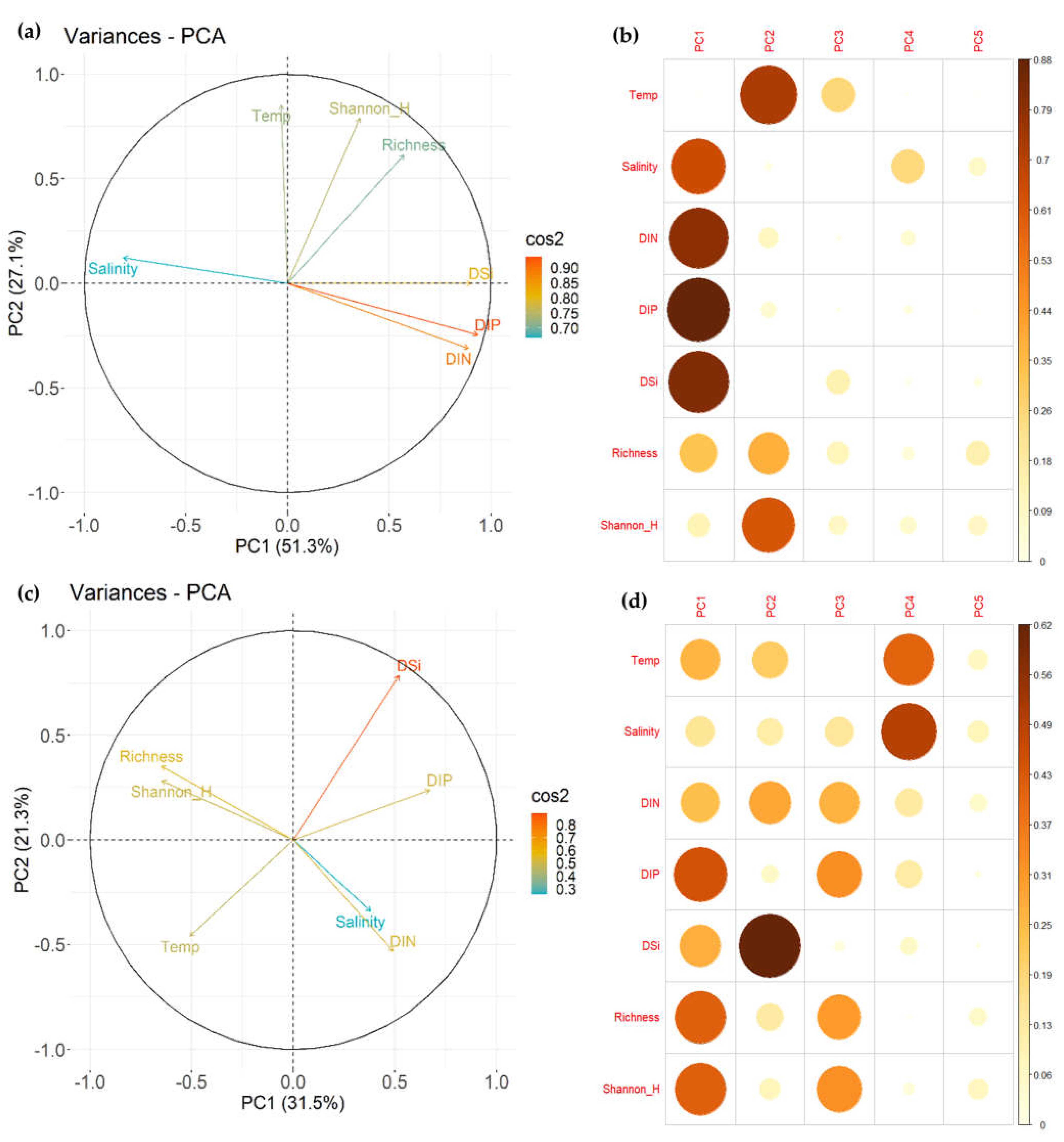
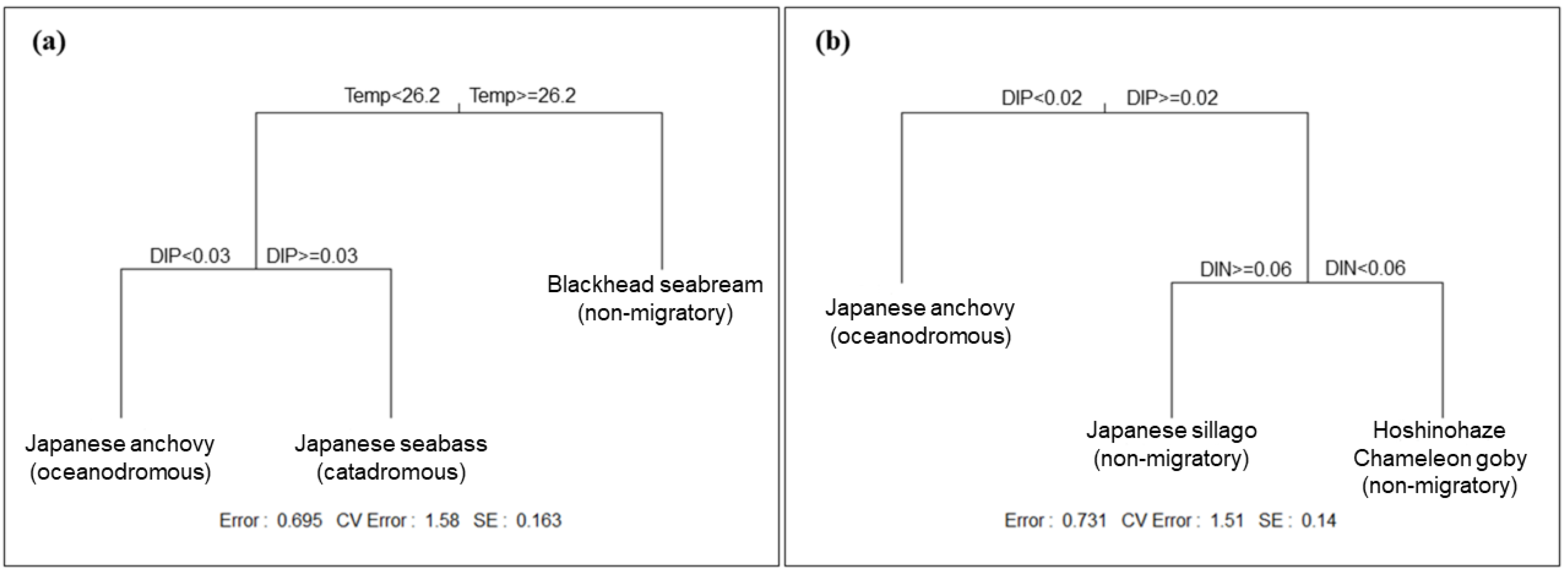
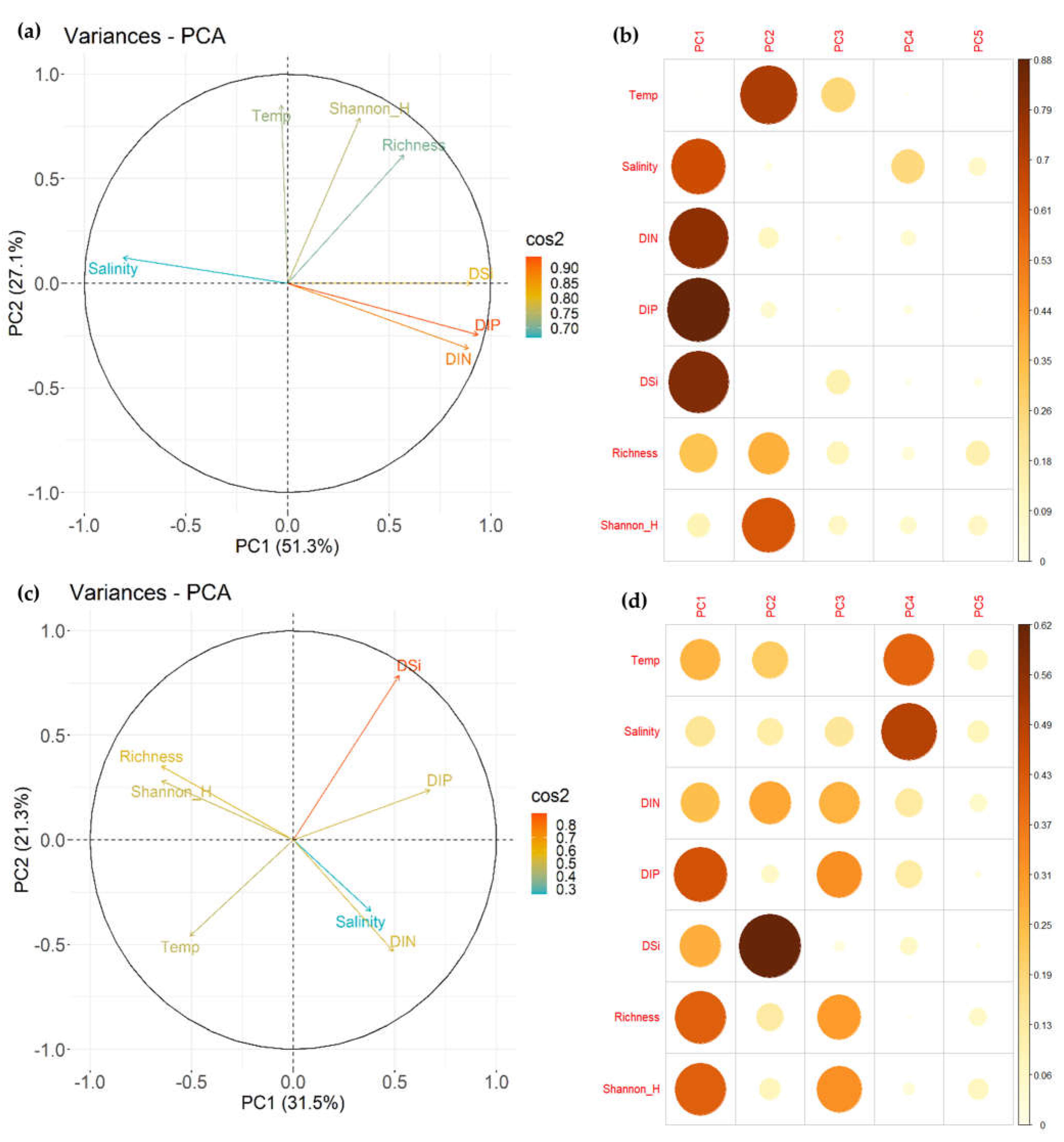
3.3. Influence of Environmental Factors on Fish Communities
3.4. Comparison of eDNA-Based Species with Previous Reports
4. Discussion
4.1. Controlling Factors of the Seasonal Fluctuations in Fish Community Compositions
4.2. Comprehensive Fish Species Detection through This Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedlander, A.M.; Ballesteros, E.; Caselle, J.E.; Gaymer, C.F.; Palma, A.T.; Petit, I.; Varas, E.; Muñoz Wilson, A.; Sala, E. Marine Biodiversity in Juan Fernández and Desventuradas Islands, Chile: Global Endemism Hotspots. PLoS ONE 2016, 11, e0145059. [Google Scholar] [CrossRef]
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- FAO. Coastal Fisheries Initiative. Available online: https://www.fao.org/in-action/coastal-fisheries-initiative/overview/context/en/ (accessed on 3 January 2024).
- Henriques, S.; Guilhaumon, F.; Villéger, S.; Amoroso, S.; França, S.; Pasquaud, S.; Cabral, H.N.; Vasconcelos, R.P. Biogeographical Region and Environmental Conditions Drive Functional Traits of Estuarine Fish Assemblages Worldwide. Fish Fish. 2017, 18, 752–771. [Google Scholar] [CrossRef]
- Endo, H.; Matsuura, K. Geography, Currents, and Fish Diversity of Japan. In Fish Diversity of Japan; Springer Nature Singapore: Singapore, 2022; pp. 7–18. [Google Scholar]
- Kume, M.; Lavergne, E.; Ahn, H.; Terashima, Y.; Kadowaki, K.; Ye, F.; Kameyama, S.; Kai, Y.; Henmi, Y.; Yamashita, Y.; et al. Factors Structuring Estuarine and Coastal Fish Communities across Japan Using Environmental DNA Metabarcoding. Ecol. Indic. 2021, 121, 107216. [Google Scholar] [CrossRef]
- Kim, E.-B.; Sagong, H.; Lee, J.-H.; Kim, G.; Kwon, D.-H.; Kim, Y.; Kim, H.-W. Environmental DNA Metabarcoding Analysis of Fish Assemblages and Phytoplankton Communities in a Furrowed Seabed Area Caused by Aggregate Mining. Front. Mar. Sci. 2022, 9, 788380. [Google Scholar] [CrossRef]
- Li, C.; Long, H.; Yang, S.; Zhang, Y.; Tang, F.; Jin, W.; Wang, G.; Chang, W.; Pi, Y.; Gao, L.; et al. EDNA Assessment of Pelagic Fish Diversity, Distribution, and Abundance in the Central Pacific Ocean. Reg. Stud. Mar. Sci. 2022, 56, 102661. [Google Scholar] [CrossRef]
- Pintueles-Tamayo, J.F.; Nieto-Navarro, J.T.; Marquez-Cuétara, A.; Domínguez-Ojeda, D.; Zetina-Rejón, M.J. Fish Community Diversity in a Coastal Zone at Southeastern Gulf of California, Mexico. Environ. Sci. Pollut. Res. 2022, 29, 61747–61766. [Google Scholar] [CrossRef]
- Lee, Y.-D.; Lee, G.-M.; Gwak, W.-S. Assessment of Fish Diversity in the Coastal Waters off Nodaedo Island, Tongyeong, Korea, Using an Underwater Visual Census and Environmental DNA Metabarcoding. Mar. Biol. 2024, 171, 23. [Google Scholar] [CrossRef]
- Itsukushima, R. Broad-Scale Environmental Factors Determining Fish Species Composition of River Estuaries in the Japanese Archipelago. Estuaries Coasts 2023, 46, 1829–1843. [Google Scholar] [CrossRef]
- Riofrío-Lazo, M.; Zetina-Rejón, M.J.; Vaca-Pita, L.; Murillo-Posada, J.C.; Páez-Rosas, D. Fish Diversity Patterns along Coastal Habitats of the Southeastern Galapagos Archipelago and Their Relationship with Environmental Variables. Sci. Rep. 2022, 12, 3604. [Google Scholar] [CrossRef]
- Luo, Z.; Yang, C.; Wang, L.; Liu, Y.; Shan, B.; Liu, M.; Chen, C.; Guo, T.; Sun, D. Relationships between Fish Community Structure and Environmental Factors in the Nearshore Waters of Hainan Island, South China. Diversity 2023, 15, 901. [Google Scholar] [CrossRef]
- Lee, Y.D.; Lee, G.M.; Park, J.Y.; Gwak, W.S. Comparing Environmental DNA Metabarcoding and Underwater Visual Census to Monitor Korean Coastal Fish Community. Ocean Sci. J. 2022, 57, 592–606. [Google Scholar] [CrossRef]
- Nguyen, B.N.; Shen, E.W.; Seemann, J.; Correa, A.M.S.; O’Donnell, J.L.; Altieri, A.H.; Knowlton, N.; Crandall, K.A.; Egan, S.P.; McMillan, W.O.; et al. Environmental DNA Survey Captures Patterns of Fish and Invertebrate Diversity across a Tropical Seascape. Sci. Rep. 2020, 10, 6729. [Google Scholar] [CrossRef]
- Gillespie, R.G. Oceanic Islands: Models of Diversity. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2001; pp. 590–599. [Google Scholar]
- Pippard, H.; Ralph, G.M.; Harvey, M.S.; Carpenter, K.E.; Buchanan, J.R.; Greenfield, D.W.; Harwell, H.D.; Larson, H.K.; Lawrence, A.; Linardich, C.; et al. The Conservation Status of Marine Biodiversity of the Pacific Islands of Oceania; IUCN International Union for Conservation of Nature: Gland, Switzerland, 2017. [Google Scholar]
- Ceríaco, L.M.P.; Santos, B.S.; de Lima, R.F.; Bell, R.C.; Norder, S.J.; Melo, M. Physical Geography of the Gulf of Guinea Oceanic Islands. In Biodiversity of the Gulf of Guinea Oceanic Islands; Springer International Publishing: Cham, Switzerland, 2022; pp. 13–36. [Google Scholar]
- Cheng, X.; Wang, Z.; Zhang, S.; Zhao, X.; Lin, J.; Huang, H.; Chen, Y.; Zou, Q. Analysis of Short-Term and Local Scale Variations in Fish Community Structure in Dachen Island Waters. Front. Mar. Sci. 2023, 10, 1199524. [Google Scholar] [CrossRef]
- Baptista, J.; Martinho, F.; Martins, R.; Carneiro, M.; Azevedo, M.; Vieira, A.R.; Gomes, P.; Pardal, M.A. Water Temperature Gradient Shapes the Structure and Composition of Nearshore Marine Fish Communities in Southern Europe. J. Sea Res. 2019, 154, 101807. [Google Scholar] [CrossRef]
- Getz, E.; Eckert, C. Effects of Salinity on Species Richness and Community Composition in a Hypersaline Estuary. Estuaries Coasts 2023, 46, 2175–2189. [Google Scholar] [CrossRef]
- Jiang, P.; Zhang, S.; Xu, S.; Xiong, P.; Cao, Y.; Chen, Z.; Li, M. Comparison of Environmental DNA Metabarcoding and Bottom Trawling for Detecting Seasonal Fish Communities and Habitat Preference in a Highly Disturbed Estuary. Ecol. Indic. 2023, 146, 109754. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, P.; Wang, L.; Liu, L.; Li, M.; Zou, K. A Comparison of Seasonal Composition and Structure of Fish Community between Environmental DNA Technology and Gillnetting in the Pearl River Estuary, China. Ecol. Indic. 2023, 147, 109915. [Google Scholar] [CrossRef]
- Nishijima, W.; Umehara, A.; Yamamoto, K.; Asaoka, S.; Fujii, N.; Otani, S.; Wang, F.; Okuda, T.; Nakai, S. Temporal Distribution of Primary and Secondary Production Estimated from Water Quality Data in the Seto Inland Sea, Japan. Ecol. Indic. 2021, 124, 107405. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chiu, M.Y.; Shih, Y.M.; Chen, I.S.; Lee, M.A.; Shao, K.T. Species Composition and Assemblages of Ichthyoplankton during Summer in the East China Sea. Cont. Shelf Res. 2016, 126, 64–78. [Google Scholar] [CrossRef]
- Shiomoto, A.; Inoue, K. Seasonal Variations of Size-Fractionated Chlorophyll a and Primary Production in the Coastal Area of Hokkaido in the Okhotsk Sea. SN Appl. Sci. 2020, 2, 1880. [Google Scholar] [CrossRef]
- Szabó, P. Ideal Free Distribution of Metabolic Activity: Implications of Seasonal Metabolic-Activity Patterns on Competitive Coexistence. Theor. Popul. Biol. 2016, 111, 1–8. [Google Scholar] [CrossRef]
- Bloomfield, E.J.; Guzzo, M.M.; Middel, T.A.; Ridgway, M.S.; McMeans, B.C. Seasonality Can Affect Ecological Interactions between Fishes of Different Thermal Guilds. Front. Ecol. Evol. 2022, 10, 986459. [Google Scholar] [CrossRef]
- Bergström, L.; Bergström, U.; Olsson, J.; Carstensen, J. Coastal Fish Indicators Response to Natural and Anthropogenic Drivers–Variability at Temporal and Different Spatial Scales. Estuar. Coast. Shelf Sci. 2016, 183, 62–72. [Google Scholar] [CrossRef]
- Long, X.; Wan, R.; Li, Z.; Ren, Y.; Song, P.; Tian, Y.; Xu, B.; Xue, Y. Spatio-Temporal Distribution of Konosirus Punctatus Spawning and Nursing Ground in the South Yellow Sea. Acta Oceanol. Sin. 2021, 40, 133–144. [Google Scholar] [CrossRef]
- Shin, D.; Park, T.H.; Kim, D.; Lee, C.I.; Lee, S.J.; Lee, J.H.; Kang, S.; Park, H.J. Spatial and Temporal Variations in Trophic Structure of Fish Assemblages in the Yellow Sea Revealed by C and N Stable Isotopes. Mar. Pollut. Bull. 2023, 196, 115678. [Google Scholar] [CrossRef]
- Fujita, T.; Yamamoto, M.; Kono, N.; Tomiyama, T.; Sugimatsu, K.; Yoneda, M. Temporal Variations in Hatch Date and Early Survival of Japanese Anchovy (Engraulis japonicus) in Response to Environmental Factors in the Central Seto Inland Sea, Japan. Fish. Oceanogr. 2021, 30, 527–541. [Google Scholar] [CrossRef]
- Da Silva, V.E.L.; Dolbeth, M.; Fabré, N.N. Assessing Tropical Coastal Dynamics across Habitats and Seasons through Different Dimensions of Fish Diversity. Mar. Environ. Res. 2021, 171, 105458. [Google Scholar] [CrossRef]
- Edgar, G.J.; Stuart-Smith, R.D. Systematic Global Assessment of Reef Fish Communities by the Reef Life Survey Program. Sci. Data 2014, 1, 140007. [Google Scholar] [CrossRef]
- Whitmarsh, S.K.; Fairweather, P.G.; Huveneers, C. What Is Big BRUVver up to? Methods and Uses of Baited Underwater Video. Rev. Fish Biol. Fish. 2017, 27, 53–73. [Google Scholar] [CrossRef]
- Ortega, A.; Geraldi, N.R.; Duarte, C.M. Environmental DNA Identifies Marine Macrophyte Contributions to Blue Carbon Sediments. Limnol. Oceanogr. 2020, 65, 3139–3149. [Google Scholar] [CrossRef]
- Wei, N.; Nakajima, F.; Tobino, T. Variation of Environmental DNA in Sediment at Different Temporal Scales in Nearshore Area of Tokyo Bay. J. Water Environ. Technol. 2019, 17, 153–162. [Google Scholar] [CrossRef]
- Selleslagh, J.; Amara, R. Environmental Factors Structuring Fish Composition and Assemblages in a Small Macrotidal Estuary (Eastern English Channel). Estuar. Coast. Shelf Sci. 2008, 79, 507–517. [Google Scholar] [CrossRef]
- Minamoto, T.; Fukuda, M.; Katsuhara, K.R.; Fujiwara, A.; Hidaka, S.; Yamamoto, S.; Takahashi, K.; Masuda, R. Environmental DNA Reflects Spatial and Temporal Jellyfish Distribution. PLoS ONE 2017, 12, e0173073. [Google Scholar] [CrossRef]
- Sakata, M.K.; Maki, N.; Sugiyama, H.; Minamoto, T. Identifying a Breeding Habitat of a Critically Endangered Fish, Acheilognathus Typus, in a Natural River in Japan. Sci. Nat. 2017, 104, 100. [Google Scholar] [CrossRef]
- Fujii, K.; Doi, H.; Matsuoka, S.; Nagano, M.; Sato, H.; Yamanaka, H. Environmental DNA Metabarcoding for Fish Community Analysis in Backwater Lakes: A Comparison of Capture Methods. PLoS ONE 2019, 14, e0210357. [Google Scholar] [CrossRef]
- Oka, S.; Doi, H.; Miyamoto, K.; Hanahara, N.; Sado, T.; Miya, M. Environmental DNA Metabarcoding for Biodiversity Monitoring of a Highly Diverse Tropical Fish Community in a Coral Reef Lagoon: Estimation of Species Richness and Detection of Habitat Segregation. Environ. DNA 2021, 3, 55–69. [Google Scholar] [CrossRef]
- DiBattista, J.D.; Fowler, A.M.; Riley, I.J.; Reader, S.; Hay, A.; Parkinson, K.; Hobbs, J.-P.A. The Use of Environmental DNA to Monitor Impacted Coastal Estuaries. Mar. Pollut. Bull. 2022, 181, 113860. [Google Scholar] [CrossRef]
- Cheang, C.C.; Lee, B.Y.; Ip, B.H.Y.; Yiu, W.H.; Tsang, L.M.; Ang, P.O. Fish and Crustacean Biodiversity in an Outer Maritime Estuary of the Pearl River Delta Revealed by Environmental DNA. Mar. Pollut. Bull. 2020, 161, 111707. [Google Scholar] [CrossRef]
- Bleijswijk, J.D.L.; Engelmann, J.C.; Klunder, L.; Witte, H.J.; Witte, J.I.; Veer, H.W. Analysis of a Coastal North Sea Fish Community: Comparison of Aquatic Environmental DNA Concentrations to Fish Catches. Environ. DNA 2020, 2, 429–445. [Google Scholar] [CrossRef]
- Sigsgaard, E.E.; Carl, H.; Møller, P.R.; Thomsen, P.F. Monitoring the Near-Extinct European Weather Loach in Denmark Based on Environmental DNA from Water Samples. Biol. Conserv. 2015, 183, 46–52. [Google Scholar] [CrossRef]
- Chucholl, F.; Fiolka, F.; Segelbacher, G.; Epp, L.S. EDNA Detection of Native and Invasive Crayfish Species Allows for Year-Round Monitoring and Large-Scale Screening of Lotic Systems. Front. Environ. Sci. 2021, 9, 639380. [Google Scholar] [CrossRef]
- Fernandez, S.; Miller, D.L.; Holman, L.E.; Gittenberger, A.; Ardura, A.; Rius, M.; Mirimin, L. Environmental DNA Sampling Protocols for the Surveillance of Marine Non-Indigenous Species in Irish Coastal Waters. Mar. Pollut. Bull. 2021, 172, 112893. [Google Scholar] [CrossRef]
- Thalinger, B.; Wolf, E.; Traugott, M.; Wanzenböck, J. Monitoring Spawning Migrations of Potamodromous Fish Species via EDNA. Sci. Rep. 2019, 9, 15388. [Google Scholar] [CrossRef]
- Boussarie, G.; Bakker, J.; Wangensteen, O.S.; Mariani, S.; Bonnin, L.; Juhel, J.-B.; Kiszka, J.J.; Kulbicki, M.; Manel, S.; Robbins, W.D.; et al. Environmental DNA Illuminates the Dark Diversity of Sharks. Sci. Adv. 2018, 4, eaap9661. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.P.; Port, J.A.; Yamahara, K.M.; Crowder, L.B. Using Environmental DNA to Census Marine Fishes in a Large Mesocosm. PLoS ONE 2014, 9, e86175. [Google Scholar] [CrossRef] [PubMed]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a Set of Universal PCR Primers for Metabarcoding Environmental DNA from Fishes: Detection of More than 230 Subtropical Marine Species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Masuda, R.; Sato, Y.; Sado, T.; Araki, H.; Kondoh, M.; Minamoto, T.; Miya, M. Environmental DNA Metabarcoding Reveals Local Fish Communities in a Species-Rich Coastal Sea. Sci. Rep. 2017, 7, 40368. [Google Scholar] [CrossRef] [PubMed]
- McElroy, M.E.; Dressler, T.L.; Titcomb, G.C.; Wilson, E.A.; Deiner, K.; Dudley, T.L.; Eliason, E.J.; Evans, N.T.; Gaines, S.D.; Lafferty, K.D.; et al. Calibrating Environmental DNA Metabarcoding to Conventional Surveys for Measuring Fish Species Richness. Front. Ecol. Evol. 2020, 8, 276. [Google Scholar] [CrossRef]
- Okaichi, T. Environmental Management of the Semi Enclosed Sea and Red Tide Problems in the Seto Inland Sea. Fish. Sci. 2002, 68, 483–488. [Google Scholar] [CrossRef] [PubMed]
- JMA. Coastal Sea Surface Temperature Information Bingonada/Hibinada. Available online: https://www.data.jma.go.jp/kaiyou/data/db/kaikyo/series/engan/engan509.html (accessed on 2 March 2024).
- International EMECS Center. Environmental Conservation of the Seto Inland Sea; International EMECS Center, Ed.; Kobe International EMECS Center: Kobe, Japan, 2008. [Google Scholar]
- Zhu, A.; Saito, M.; Onodera, S.; Shimizu, Y.; Jin, G.; Ohta, T.; Chen, J. Evaluation of the Spatial Distribution of Submarine Groundwater Discharge in a Small Island Scale Using the 222Rn Tracer Method and Comparative Modeling. Mar. Chem. 2019, 209, 25–35. [Google Scholar] [CrossRef]
- Kamimura, Y.; Shoji, J. Seasonal Changes in the Fish Assemblage in a Mixed Vegetation Area of Seagrass and Macroalgae in the Central Seto Inland Sea. Aquac. Sci. 2009, 57, 233–241. [Google Scholar] [CrossRef]
- Mohri, K.; Kamimura, Y.; Mizuno, K.; Kinoshita, H.; Toshito, S.; Shoji, J. Seasonal Changes in the Fish Assemblage in a Seagrass Bed in the Central Seto Inland Sea. Aquac. Sci. 2013, 61, 215–220. [Google Scholar] [CrossRef]
- Onodera, S.; Saito, M.; Hayashi, M.; Sawano, M. Nutrient Dynamics with Groundwater-Seawater Interactions in a Beach Slope of a Steep Island, Western Japan. IAHS Publ. 2007, 312, 150. [Google Scholar]
- Saito, M.; Onodera, S.; Jin, G.; Shimizu, Y.; Taniguchi, M. Nitrogen Dynamics in a Highly Urbanized Coastal Area of Western Japan: Impact of Sewage-Derived Loads. Prog. Earth Planet. Sci. 2018, 5, 17. [Google Scholar] [CrossRef]
- Minamoto, T.; Miya, M.; Sado, T.; Seino, S.; Doi, H.; Kondoh, M.; Nakamura, K.; Takahara, T.; Yamamoto, S.; Yamanaka, H.; et al. An Illustrated Manual for Environmental DNA Research: Water Sampling Guidelines and Experimental Protocols. Environ. DNA 2021, 3, 8–13. [Google Scholar] [CrossRef]
- Sato, Y.; Miya, M.; Fukunaga, T.; Sado, T.; Iwasaki, W. MitoFish and MiFish Pipeline: A Mitochondrial Genome Database of Fish with an Analysis Pipeline for Environmental DNA Metabarcoding. Mol. Biol. Evol. 2018, 35, 1553–1555. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, W.; Fukunaga, T.; Isagozawa, R.; Yamada, K.; Maeda, Y.; Satoh, T.P.; Sado, T.; Mabuchi, K.; Takeshima, H.; Miya, M.; et al. Mitofish and Mitoannotator: A Mitochondrial Genome Database of Fish with an Accurate and Automatic Annotation Pipeline. Mol. Biol. Evol. 2013, 30, 2531–2540. [Google Scholar] [CrossRef]
- Zhu, T.; Sato, Y.; Sado, T.; Miya, M.; Iwasaki, W. MitoFish, MitoAnnotator, and MiFish Pipeline: Updates in 10 Years. Mol. Biol. Evol. 2023, 40, msad035. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Laporte, M.; Berger, C.S.; García-Machado, E.; Côté, G.; Morissette, O.; Bernatchez, L. Cage Transplant Experiment Shows Weak Transport Effect on Relative Abundance of Fish Community Composition as Revealed by EDNA Metabarcoding. Ecol. Indic. 2022, 137, 108785. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A Tribute to Claude Shannon (1916–2001) and a Plea for More Rigorous Use of Species Richness, Species Diversity and the ‘Shannon-Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-Parametric Multivariate Analyses of Changes in Community Structure. Austral. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal Component Analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Glenn, D. Multivariate Regression Trees: A New Technique for Modeling Species–Environment Relationships. Ecology 2002, 83, 1105–1117. [Google Scholar] [CrossRef]
- Makowski, D.; Ben-Shachar, M.; Patil, I.; Lüdecke, D. Methods and Algorithms for Correlation Analysis in R. J. Open Source Softw. 2020, 5, 2306. [Google Scholar] [CrossRef]
- Huang, H. LinkET: Everything Is Linkable. In R Package Version 0.0, Shenzhen, China, 2021; Volume 2. Available online: https://github.com/Hy4m/linkET (accessed on 15 February 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Zhang, Z.; Xu, S.; Capinha, C.; Weterings, R.; Gao, T. Using Species Distribution Model to Predict the Impact of Climate Change on the Potential Distribution of Japanese Whiting Sillago Japonica. Ecol. Indic. 2019, 104, 333–340. [Google Scholar] [CrossRef]
- Yoneda, M.; Fujita, T.; Yamamoto, M.; Tadokoro, K.; Okazaki, Y.; Nakamura, M.; Takahashi, M.; Kono, N.; Matsubara, T.; Abo, K.; et al. Bottom-up Processes Drive Reproductive Success of Japanese Anchovy in an Oligotrophic Sea: A Case Study in the Central Seto Inland Sea, Japan. Prog. Oceanogr. 2022, 206, 102860. [Google Scholar] [CrossRef]
- Sasano, S.; Murakami, H.; Suzuki, K.W.; Minamoto, T.; Yamashita, Y.; Masuda, R. Seasonal Changes in the Distribution of Black Sea Bream Acanthopagrus Schlegelii Estimated by Environmental DNA. Fish. Sci. 2022, 88, 91–107. [Google Scholar] [CrossRef]
- Kawai, K.; Fujita, H.; Sanchez, G.; Furusawa, S.; Umino, T. Estimating the Spawning Season of Black Sea Bream Acanthopagrus Schlegelii in Hiroshima Bay, Japan, from Temporal Variation in Egg Density. Fish. Sci. 2020, 86, 645–653. [Google Scholar] [CrossRef]
- Kong, L.; Kawasaki, M.; Kuroda, K.; Kohno, H.; Fujita, K. Spawning Characteristics of the Konoshiro Gizzard Shad in Tokyo and Sagami Bays, Central Japan. Fish. Sci. 2004, 70, 116–122. [Google Scholar] [CrossRef]
- Gwak, W.-S.; Zhang, Q.; Roy, A. Low Genetic Diversity and Shallow Population Structure of the Japanese Halfbeak Hyporhamphus Sajori Revealed from Mitochondrial DNA in the Northeast Asia. Korean J. Ichthyol. 2019, 31, 187–194. [Google Scholar] [CrossRef]
- Oya, F.; Tsuji, T.; Fujiwara, S.-I. Relative Growth and Feeding Habits of Halfbeak, Hyporhamphus Sajori, Larvae and Juveniles in Toyama Bay of the Japan Sea. Aquac. Sci. 2002, 50, 47–54. [Google Scholar]
- Niu, M.; Jin, X.; Li, X.; Wang, J. Effects of Spatio-Temporal and Environmental Factors on Distribution and Abundance of Wintering Anchovy Engraulis japonicus in Central and Southern Yellow Sea. Chin. J. Oceanol. Limnol. 2014, 32, 565–575. [Google Scholar] [CrossRef]
- Sakamoto, W.; Tanaka, Y. Water Temperature Patterns and Distributions of Fish Eggs and Larvae in the Vicinity of Shallow Sea Front. NIPPON SUISAN GAKKAISHI 1986, 52, 767–776. [Google Scholar] [CrossRef]
- Park, J.M.; Huh, S.H.; Baeck, G.W. Temporal Variations of Fish Assemblage in the Surf Zone of the Nakdong River Estuary, Southeastern Korea. Anim. Cells Syst. 2015, 19, 350–358. [Google Scholar] [CrossRef]
- Zhu, Y.; Minami, K.; Iwahara, Y.; Oda, K.; Hidaka, K.; Hoson, O.; Morishita, K.; Hirota, M.; Tsuru, S.; Shirakawa, H.; et al. Seasonal Variation in Fish School Spatial Distribution and Abundance under the Kuroshio Regular Pattern and the Large Meander in Suzu Coastal Waters. PLoS ONE 2021, 16, e0260629. [Google Scholar] [CrossRef] [PubMed]
- Onomichi City Hall. Onomichi City Fisheries Promotion Vision; Onomichi City Hall: Onomichi City, Japan, 2019. [Google Scholar]
- Oehler, T.; Eiche, E.; Putra, D.; Adyasari, D.; Hennig, H.; Mallast, U.; Moosdorf, N. Seasonal Variability of Land-Ocean Groundwater Nutrient Fluxes from a Tropical Karstic Region (Southern Java, Indonesia). J. Hydrol. 2018, 565, 662–671. [Google Scholar] [CrossRef]
- Sugimoto, R.; Honda, H.; Kobayashi, S.; Takao, Y.; Tahara, D.; Tominaga, O.; Taniguchi, M. Seasonal Changes in Submarine Groundwater Discharge and Associated Nutrient Transport into a Tideless Semi-Enclosed Embayment (Obama Bay, Japan). Estuaries Coasts 2016, 39, 13–26. [Google Scholar] [CrossRef]
- Tada, K.; Monaka, K.; Morishita, M.; Hashimoto, T. Standing Stocks and Production Rates of Phytoplankton and Abundance of Bacteria in the Seto Inland Sea, Japan. J. Oceanogr. 1998, 54, 285–295. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Hirade, N.; Higashizono, K.; Tada, K.; Kishimoto, K.; Oyama, K.; Ichimi, K. Light and Nutrient Limitation on Phytoplankton Production in the Strait of an Enclosed Coastal Sea (Bisan Strait, Eastern Seto Inland Sea, Japan). J. Sea Res. 2015, 103, 75–83. [Google Scholar] [CrossRef]
- Cloern, J. Our Evolving Conceptual Model of the Coastal Eutrophication Problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef]
- Cermeño, P.; Marañón, E.; Pérez, V.; Serret, P.; Fernández, E.; Castro, C.G. Phytoplankton Size Structure and Primary Production in a Highly Dynamic Coastal Ecosystem (Ría de Vigo, NW-Spain): Seasonal and Short-Time Scale Variability. Estuar. Coast. Shelf Sci. 2006, 67, 251–266. [Google Scholar] [CrossRef]
- Lavanya, D.; Ramalingaiah, D.; Suguna, T.; Kumar Reddy, D.R.; Madhavi, K. Food and Feeding Ecology of Mugil Cephalus from Krishnapatnam and Mypadu Coasts of Nellore District, Andhra Pradesh. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2616–2630. [Google Scholar] [CrossRef]
- Nixon, S.W.; Oviatt, C.A.; Frithsen, J.; Sullivan, B. Nutrients And The Productivity Of Estuarine And Coastal Marine Ecosystems. J. Limnol. Soc. S. Afr. 1986, 12, 43–71. [Google Scholar] [CrossRef]
- Czeglédi, I.; Sály, P.; Specziár, A.; Preiszner, B.; Szalóky, Z.; Maroda, Á.; Pont, D.; Meulenbroek, P.; Valentini, A.; Erős, T. Congruency between Two Traditional and EDNA-Based Sampling Methods in Characterising Taxonomic and Trait-Based Structure of Fish Communities and Community-Environment Relationships in Lentic Environment. Ecol. Indic. 2021, 129, 107952. [Google Scholar] [CrossRef]
- Snigirov, S.; Goncharov, O.; Sylantyev, S. The Fish Community in Zmiinyi Island Waters: Structure and Determinants. Mar. Biodivers. 2012, 42, 225–239. [Google Scholar] [CrossRef]
- Sugimoto, K.; Nakano, Y.; Okuda, T.; Nakai, S.; Nishijima, W.; Okada, M. Coexistence between Zostera Marina and Zostera Japonica in Seagrass Beds of the Seto Inland Sea, Japan. J. Ecol. Environ. 2017, 41, 523. [Google Scholar] [CrossRef]
- Zamora-López, A.; Guerrero-Gómez, A.; Torralva, M.; Zamora-Marín, J.M.; Guillén-Beltrán, A.; Oliva-Paterna, F.J. Shallow Waters as Critical Habitats for Fish Assemblages under Eutrophication-Mediated Events in a Coastal Lagoon. Estuar. Coast Shelf Sci. 2023, 291, 108447. [Google Scholar] [CrossRef]
- Afzali, S.F.; Bourdages, H.; Laporte, M.; Mérot, C.; Normandeau, E.; Audet, C.; Bernatchez, L. Comparing Environmental Metabarcoding and Trawling Survey of Demersal Fish Communities in the Gulf of St. Lawrence, Canada. Env. DNA 2021, 3, 22–42. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Summer | Autumn | |||
|---|---|---|---|---|---|
| Variable | Mean | SD | Mean | SD | |
| Temp (°C) | 26.5 | ±0.67 | 20.7 | ±0.55 | *** |
| Salinity (psu) | 30.5 | ±0.19 | 33.5 | ±0.10 | *** |
| DIN (mg/L) | 0.02 | ±0.02 | 0.06 | ±0.01 | *** |
| DIP (mg/L) | 0.04 | ±0.02 | 0.02 | ±0.00 | ** |
| DSi (mg/L) | 0.57 | ±0.12 | 0.2 | ±0.02 | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nhat, N.H.; Saito, M.; Hamada, M.; Onodera, S.-i. Evaluation of the Effects of Environmental Factors on Seasonal Variations in Fish Diversity on a Coastal Island in Western Japan. Environments 2024, 11, 60. https://doi.org/10.3390/environments11030060
Nhat NH, Saito M, Hamada M, Onodera S-i. Evaluation of the Effects of Environmental Factors on Seasonal Variations in Fish Diversity on a Coastal Island in Western Japan. Environments. 2024; 11(3):60. https://doi.org/10.3390/environments11030060
Chicago/Turabian StyleNhat, Nguyen Hong, Mitsuyo Saito, Mayuko Hamada, and Shin-ichi Onodera. 2024. "Evaluation of the Effects of Environmental Factors on Seasonal Variations in Fish Diversity on a Coastal Island in Western Japan" Environments 11, no. 3: 60. https://doi.org/10.3390/environments11030060