
Impairments of Spatial Memory and N-methyl-d-aspartate Receptors and Their Postsynaptic Signaling Molecules in the Hippocampus of Developing Rats Induced by As, Pb, and Mn Mixture Exposure


Abstract
:
1. Background
2. Materials and Methods
2.1. Animals and Treatment
2.2. Neurobehavioral Studies
2.2.1. Spatial Learning and Memory Performance by Morris Water Maze test
2.2.2. Continuous Alternation and Spatial Memory in Y-Maze
2.2.3. Memory in a Passive Avoidance System
2.2.4. Learning in an Active Avoidance System
2.3. Neurochemical Studies
2.3.1. mRNA Expression of Hippocampal NR2A, NR2B, and NR1 Genes
2.3.2. Immunodetection of Proteins by Western Blotting
2.3.3. Estimation of As, Pb, and Mn Levels in the Hippocampus
2.3.4. Protein Estimation
2.3.5. Statistical Analysis
3. Results
3.1. Neurobehavioral Studies
3.1.1. The Effect on Spatial Memory and Learning: Morris Water Maze Analysis
3.1.2. Effect on Continuous Alternation and Spatial Memory in the Y-Maze
3.1.3. Effect on Memory in the Passive Avoidance System
3.1.4. Effect on Learning Ability in the Active Avoidance System
3.2. Neurochemical Studies
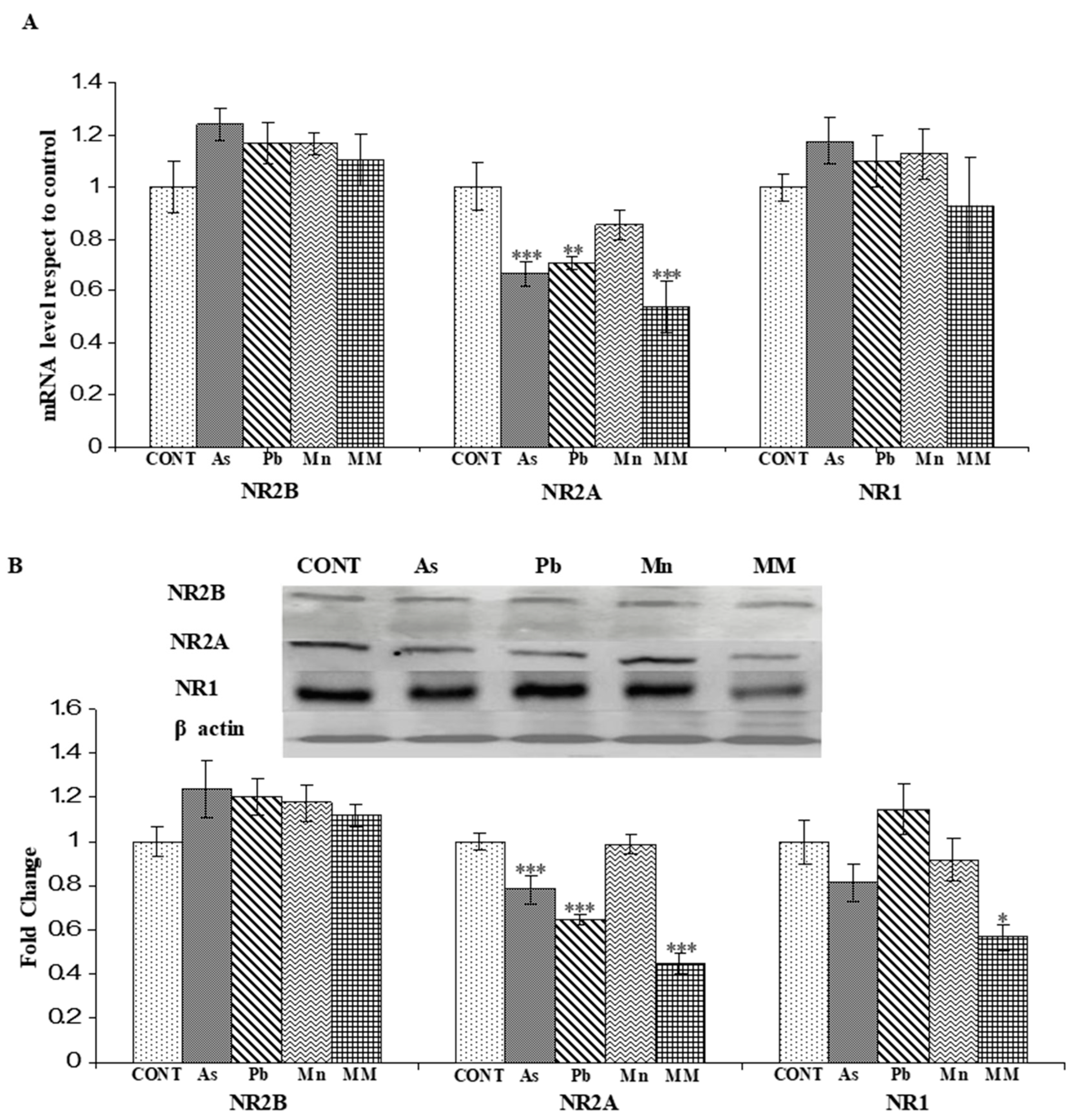
3.2.1. Effect on the mRNA Expression of NR2A, NR2B, and NR1 Genes
3.2.2. Effect on the Protein Expression of NNMDARs Subunits (NR2A, NR2B, and NR1) and Their Postsynaptic Signaling Proteins (PSD-95, SynGAP, pCAMKII, and pCREB)
3.2.3. As, Pb, and Mn Levels in the Hippocampus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karri, V.; Schuhmacher, M.; Kumar, V. Heavy metals (Pb, Cd, As and MeHg) as risk factors for cognitive dysfunction: A general review of metal mixture mechanism in brain. Environ. Toxicol. Pharmacol. 2016, 48, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Bakulski, K.M.; Seo, Y.A.; Hickman, R.C.; Brandt, D.; Vadari, H.S.; Hu, H.; Park, S.K. Heavy Metals Exposure and Alzheimer’s Disease and Related Dementias. J. Alzheimer’s Dis. 2020, 76, 1215–1242. [Google Scholar] [CrossRef] [PubMed]
- Farace, C.; Fiorito, G.; Pisano, A.; Etzi, F.; Sabalic, A.; Fenu, G.; Asara, Y.; Solinas, G.; Madeddu, R. Human tissue lead (Pb) levels and amyotrophic lateral sclerosis: A systematic review and meta-analysis of case-control studies. Neurol. Sci. 2022, 43, 5851–5859. [Google Scholar] [CrossRef]
- Ozturk, M.; Metin, M.; Altay, V.; Bhat, R.A.; Ejaz, M.; Gul, A.; Unal, B.T.; Hasanuzzaman, M.; Nibir, L.; Nahar, K.; et al. Arsenic and Human Health: Genotoxicity, Epigenomic Effects, and Cancer Signaling. Biol. Trace Elem. Res. 2022, 200, 988–1001. [Google Scholar] [CrossRef]
- Daka, E.R.; Hawkins, S.J. Interactive effects of copper, cadmium and lead on zinc accumulation in the gastropod mollusc Littorina saxatilis. Water Air Soil Pollut. 2006, 171, 19–28. [Google Scholar] [CrossRef]
- Borgmann, U.; Norwood, W.P.; Dixon, D.G. Modelling Bioaccumulation and Toxicity of Metal Mixtures. Hum. Ecol. Risk Assess. Int. J. 2008, 14, 266–289. [Google Scholar] [CrossRef]
- Lopes de Andrade, V.; Marreilha Dos Santos, A.P.; Aschner, M. Neurotoxicity of Metal Mixtures. Adv. Neurotoxicol. 2021, 5, 329–364. [Google Scholar]
- Sanders, A.P.; Claus Henn, B.; Wright, R.O. Perinatal and Childhood Exposure to Cadmium, Manganese, and Metal Mixtures and Effects on Cognition and Behavior: A Review of Recent Literature. Curr. Environ. Health Rep. 2015, 2, 284–294. [Google Scholar] [CrossRef]
- Zheng, T.; Zhang, J.; Sommer, K.; Bassig, B.A.; Zhang, X.; Braun, J.; Xu, S.; Boyle, P.; Zhang, B.; Shi, K.; et al. Effects of Environmental Exposures on Fetal and Childhood Growth Trajectories. Ann. Glob. Health 2016, 82, 41–99. [Google Scholar] [CrossRef]
- Luo, Y.; McCullough, L.E.; Tzeng, J.Y.; Darrah, T.; Vengosh, A.; Maguire, R.L.; Maity, A.; Samuel-Hodge, C.; Murphy, S.K.; Mendez, M.A.; et al. Maternal blood cadmium, lead and arsenic levels, nutrient combinations, and offspring birthweight. BMC Public Health 2017, 17, 354. [Google Scholar] [CrossRef] [PubMed]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Gu, T.; Jia, X.; Shi, H.; Gong, X.; Ma, J.; Gan, Z.; Yu, Z.; Li, Z.; Wei, Y. An Evaluation of Exposure to 18 Toxic and/or Essential Trace Elements Exposure in Maternal and Cord Plasma during Pregnancy at Advanced Maternal Age. Int. J. Environ. Res. Public Health 2022, 19, 14485. [Google Scholar] [CrossRef] [PubMed]
- Barbee, N.C.; Ganio, K.; Swearer, S.E. Integrating multiple bioassays to detect and assess impacts of sublethal exposure to metal mixtures in an estuarine fish. Aquat. Toxicol. 2014, 152, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S.; Liu, C.; Li, Q.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef] [PubMed]
- Delgado, C.F.; Ullery, M.A.; Jordan, M.; Duclos, C.; Rajagopalan, S.; Scott, K. Lead Exposure and Developmental Disabilities in Preschool-Aged Children. J. Public Health Manag. Pract. 2018, 24, e10–e17. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Loira, B.; Cebrián, M.E.; López-Carrillo, L. Arsenic exposure in northern Mexican women. Salud Publica Mex. 2020, 62, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Appleton, A.A.; Jackson, B.P.; Karagas, M.; Marsit, C.J. Prenatal exposure to neurotoxic metals is associated with increased placental glucocorticoid receptor DNA methylation. Epigenetics 2017, 26, 607–615. [Google Scholar] [CrossRef]
- Ma, Y.; Hu, Q.; Yang, D.; Zhao, Y.; Bai, J.; Mubarik, S.; Yu, C. Combined exposure to multiple metals on serum uric acid in NHANES under three statistical models. Chemosphere 2022, 301, 134416. [Google Scholar] [CrossRef]
- Liu, L.; Wong, T.P.; Pozza, M.F.; Lingenhoehl, K.; Wang, Y.; Sheng, M.; Auberson, Y.P.; Wang, Y.T. Role of NMDA receptor subtypes in governing the direction of hippocampal synaptic plasticity. Science 2004, 304, 1021–1024. [Google Scholar] [CrossRef]
- Figueiredo, A.; Costa, I.M.; Fernandes, T.A.; Gonçalves, L.L.; Brito, J. Food Supplements for Weight Loss: Risk Assessment of Selected Impurities. Nutrients 2020, 12, 954. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.; Lozano, G.; Hardisson, A.; Rubio, C.; González, T. Content of toxic and essential metals in canned mussels commonly consumed in Tenerife, Canary Islands, Spain. J. Food Prot. 2004, 67, 1526–1532. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.P.; Mazzella, M.J.; Malin, A.J.; Hair, G.M.; Busgang, S.A.; Saland, J.M.; Curtin, P. Combined exposure to lead, cadmium, mercury, and arsenic and kidney health in adolescents age 12–19 in NHANES 2009–2014. Environ. Int. 2019, 131, 104993. [Google Scholar] [CrossRef] [PubMed]
- Levin-Schwartz, Y.; Cowell, W.; Leon Hsu, H.H.; Enlow, M.B.; Amarasiriwardena, C.; Andra, S.S.; Wright, R.J.; Wright, R.O. Metal mixtures are associated with increased anxiety during pregnancy. Environ. Res. 2022, 204, 112276. [Google Scholar] [CrossRef] [PubMed]
- Kortenkamp, A. Low dose mixture effects of endocrine disrupters and their implications for regulatory thresholds in chemical risk assessment. Curr. Opin. Pharmacol. 2014, 19, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Claus Henn, B.; Ettinger, A.S.; Schwartz, J.; Téllez-Rojo, M.M.; Lamadrid-Figueroa, H.; Hernández-Avila, M.; Schnaas, L.; Amarasiriwardena, C.; Bellinger, D.C.; Hu, H.; et al. Early postnatal blood manganese levels and children’s neurodevelopment. Epidemiology 2010, 21, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.G.; Bellinger, D.C.; Valeri, L.; Hasan, M.O.; Quamruzzaman, Q.; Golam, M.; Kile, M.L.; Christiani, D.C.; Wright, R.O.; Mazumdar, M. Neurodevelopmental outcomes among 2- to 3-year-old children in Bangladesh with elevated blood lead and exposure to arsenic and manganese in drinking water. Environ. Health 2016, 15, 44. [Google Scholar] [CrossRef]
- Tamayo Yortiz, M.; Téllez-Rojo, M.M.; Trejo-Valdivia, B.; Schnaas, L.; Osorio-Valencia, E.; Coull, B.; Bellinger, D.; Wright, R.J.; Wright, R.O. Maternal stress modifies the effect of exposure to lead during pregnancy and 24-month old children’s neurodevelopment. Environ. Int. 2017, 98, 191–197. [Google Scholar] [CrossRef]
- Michelsen-Correa, S.; Martin, C.F.; Kirk, A.B. Evaluation of Fetal Exposures to Metals and Metalloids through Meconium Analyses: A Review. Int. J. Environ. Res. Public Health 2021, 18, 1975. [Google Scholar] [CrossRef]
- Etzel, R.A. The special vulnerability of children. Int. J. Hyg. Environ. Health 2020, 227, 113516. [Google Scholar] [CrossRef]
- Gilani, S.R.; Zaidi, S.R.; Batool, M.; Bhatti, A.A.; Durrani, A.I.; Mahmood, Z. Report: Central nervous system (CNS) toxicity caused by metal poisoning: Brain as a target organ. Pak. J. Pharm. Sci. 2015, 28, 1417–1423. [Google Scholar] [PubMed]
- Amaya, E.; Gil, F.; Freire, C.; Olmedo, P.; Fernández-Rodríguez, M.; Fernández, M.F.; Olea, N. Placental concentrations of heavy metals in a mother-child cohort. Environ. Res. 2013, 120, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Molina-Villalba, I.; Lacasaña, M.; Rodríguez-Barranco, M.; Hernández, A.F.; Gonzalez-Alzaga, B.; Aguilar-Garduño, C.; Gil, F. Biomonitoring of arsenic, cadmium, lead, manganese and mercury in urine and hair of children living near mining and industrial areas. Chemosphere 2015, 124, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Cordier, S.; Lehébel, A.; Amar, E.; Anzivino-Viricel, L.; Hours, M.; Monfort, C.; Chevrier, C.; Chiron, M.; Robert-Gnansia, E. Maternal residence near municipal waste incinerators and the risk of urinary tract birth defects. Occup. Environ. Med. 2010, 67, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Jover-Mengual, T.; Wong, J.; Bennett, M.V.; Zukin, R.S. PSD-95 and PKC converge in regulating NMDA receptor trafficking and gating. Proc. Natl. Acad. Sci. USA 2006, 103, 19902–19907. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, H.K.; Takamiya, K.; Huganir, R.L. The role of synaptic GTPase-activating protein in neuronal development and synaptic plasticity. J. Neurosci. 2003, 23, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, G.A.; Liu, X.; Loiacono, N.J.; Kline, J.; Factor-Litvak, P.; van Geen, A.; Mey, J.L.; Levy, D.; Abramson, R.; Schwartz, A.; et al. A cross-sectional study of well water arsenic and child IQ in Maine schoolchildren. Environ. Health 2014, 13, 23. [Google Scholar] [CrossRef]
- Chandravanshi, L.P.; Yadav, R.S.; Shukla, R.K.; Singh, A.; Sultana, S.; Pant, A.B.; Parmar, D.; Khanna, V.K. Reversibility of changes in brain cholinergic receptors and acetylcholinesterase activity in rats following early life arsenic exposure. Int. J. Dev. Neurosci. 2014, 34, 60–75. [Google Scholar] [CrossRef]
- Anderson, D.W.; Mettil, W.; Schneider, J.S. Effects of low level lead exposure on associative learning and memory in the rat: Influences of sex and developmental timing of exposure. Toxicol. Lett. 2016, 246, 57–64. [Google Scholar] [CrossRef]
- Jing, J.; Zheng, G.; Liu, M.; Shen, X.; Zhao, F.; Wang, J.; Zhang, J.; Huang, G.; Dai, P.; Chen, Y.; et al. Changes in the synaptic structure of hippocampal neurons and impairment of spatial memory in a rat model caused by chronic arsenite exposure. Neurotoxicology 2012, 33, 1230–1238. [Google Scholar] [CrossRef]
- Wang, X.M.; Liu, W.J.; Zhang, R.; Zhou, Y.K. Effects of exposure to low-level lead on spatial learning and memory and the expression of mGluR1, NMDA receptor in different developmental stages of rats. Toxicol. Ind. Health 2013, 29, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Bardullas, U.; Limón-Pacheco, J.H.; Giordano, M.; Carrizales, L.; Mendoza-Trejo, M.S.; Rodríguez, V.M. Chronic low-level arsenic exposure causes gender-specific alterations in locomotor activity, dopaminergic systems, and thioredoxin expression in mice. Toxicol. Appl. Pharmacol. 2009, 239, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Flores-Montoya, M.G.; Sobin, C. Early chronic lead exposure reduces exploratory activity in young C57BL/6J mice. J. Appl. Toxicol. 2015, 35, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, A.B.; Cushing, C.A.; Antonini, J.M.; Finley, B.L.; Mowat, F.S. State-of-the-science review: Does manganese exposure during welding pose a neurological risk? J. Toxicol. Environ. Health B Crit. Rev. 2007, 10, 417–465. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Ouyang, L.; Li, Q.; Yang, S.; Liu, S.; Yu, H.; Jia, Q.; Rao, S.; Xie, J.; Du, G.; et al. Hippocampal LIMK1-mediated Structural Synaptic Plasticity in Neurobehavioral Deficits Induced by a Low-dose Heavy Metal Mixture. Mol. Neurobiol. 2023, 60, 6029–6042. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Hong, N.S.; McDonald, R.J.; Wojtowicz, J.M. A role for adult neurogenesis in spatial long-term memory. Neuroscience 2005, 130, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Papadia, S.; Hardingham, G.E. The dichotomy of NMDA receptor signaling. Neuroscientist 2007, 13, 572–579. [Google Scholar] [CrossRef]
- Paoletti, P.; Ascher, P.; Neyton, J. High-affinity zinc inhibition of NMDA NR1-NR2A receptors. J. Neurosci. 1997, 17, 5711–5725. [Google Scholar] [CrossRef]
- Chatterton, J.E.; Awobuluyi, M.; Premkumar, L.S.; Takahashi, H.; Talantova, M.; Shin, Y.; Cui, J.; Tu, S.; Sevarino, K.A.; Nakanishi, N.; et al. Excitatory glycine receptors containing the NR3 family of NMDA receptor subunits. Nature 2002, 415, 793–798. [Google Scholar] [CrossRef]
- Sheng, M.; Kim, M.J. Postsynaptic signaling and plasticity mechanisms. Science 2002, 298, 776–780. [Google Scholar] [CrossRef]
- Lüscher, C.; Malenka, R.C. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 2012, 4, a005710. [Google Scholar] [CrossRef] [PubMed]
- Rusakov, D.A.; Fine, A. Extracellular Ca2+ depletion contributes to fast activity-dependent modulation of synaptic transmission in the brain. Neuron 2003, 37, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C.M. Transcription factors in long-term memory and synaptic plasticity. Physiol. Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Benito, E.; Barco, A. CREB’s control of intrinsic and synaptic plasticity: Implications for CREB-dependent memory models. Trends Neurosci. 2010, 33, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Skeberdis, V.A.; Francesconi, A.; Bennett, M.V.; Zukin, R.S. Postsynaptic density protein-95 regulates NMDA channel gating and surface expression. J. Neurosci. 2004, 24, 10138–10148. [Google Scholar] [CrossRef] [PubMed]
- MacGillavry, H.D.; Kerr, J.M.; Blanpied, T.A. Lateral organization of the postsynaptic density. Mol. Cell. Neurosci. 2011, 48, 321–331. [Google Scholar] [CrossRef]
- Chen, X.; Nelson, C.D.; Li, X.; Winters, C.A.; Azzam, R.; Sousa, A.A.; Leapman, R.D.; Gainer, H.; Sheng, M.; Reese, T.S. PSD-95 is required to sustain the molecular organization of the postsynaptic density. J. Neurosci. 2011, 31, 6329–6338. [Google Scholar] [CrossRef]
- Kim, J.H.; Liao, D.; Lau, L.F.; Huganir, R.L. SynGAP: A synaptic RasGAP that associates with the PSD-95/SAP90 protein family. Neuron 1998, 20, 683–691. [Google Scholar] [CrossRef]
- Park, C.S.; Elgersma, Y.; Grant, S.G.; Morrison, J.H. alpha-Isoform of calcium-calmodulin-dependent protein kinase II and postsynaptic density protein 95 differentially regulate synaptic expression of NR2A- and NR2B-containing N-methyl-d-aspartate receptors in hippocampus. Neuroscience 2008, 151, 43–55. [Google Scholar] [CrossRef]
- Franks, K.M.; Sejnowski, T.J. Complexity of calcium signaling in synaptic spines. Bioessays 2002, 24, 1130–1144. [Google Scholar] [CrossRef]
- Kessels, H.W.; Malinow, R. Synaptic AMPA receptor plasticity and behavior. Neuron 2009, 61, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Krinke, G.J.; Vidotto, N.; Weber, E. Teased-fiber technique for peripheral myelinated nerves: Methodology and interpretation. Toxicol. Pathol. 2000, 28, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Glowinski, J.; Iversen, L.L. Regional studies of catecholamines in the rat brain. I. The disposition of [3H]norepinephrine, [3H]dopamine and [3H]dopa in various regions of the brain. J. Neurochem. 1966, 13, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Roghani, M.; Joghataie, M.T.; Jalali, M.R.; Baluchnejadmojarad, T. Time course of changes in passive avoidance and Y—Maze performance in male diabetic rats. Iran. Biomed. J. 2006, 10, 99–104. [Google Scholar]
- Wang, X.J.; Qi, L.; Cheng, Y.F.; Ji, X.F.; Chi, T.Y.; Liu, P.; Zou, L.B. PINK1 overexpression prevents forskolin-induced tau hyperphosphorylation and oxidative stress in a rat model of Alzheimer’s disease. Acta Pharmacol. Sin. 2022, 43, 1916–1927. [Google Scholar] [CrossRef]
- Yadav, R.S.; Sankhwar, M.L.; Shukla, R.K.; Chandra, R.; Pant, A.B.; Islam, F.; Khanna, V.K. Attenuation of arsenic neurotoxicity by curcumin in rats. Toxicol. Appl. Pharmacol. 2009, 240, 367–376. [Google Scholar] [CrossRef]
- Gascón, S.; Deogracias, R.; Sobrado, M.; Roda, J.M.; Renart, J.; Rodríguez-Peña, A.; Díaz-Guerra, M. Transcription of the NR1 subunit of the N-methyl-D-aspartate receptor is down-regulated by excitotoxic stimulation and cerebral ischemia. J. Biol. Chem. 2005, 280, 35018–35027. [Google Scholar] [CrossRef]
- Soria Fregozo, C.; Miranda Beltrán, M.L.; Flores Soto, M.E.; Pérez Vega, M.I.; Beas Zárate, C.; Huacuja Ruiz, L. Expression of NMDA receptor subunits in rat prefrontal cortex with CCL4-induced hepatic damage after a treatment with Rosmarinus officinalis L. Neurologia 2012, 27, 261–267. [Google Scholar] [CrossRef]
- del Rey, A.; Randolf, A.; Wildmann, J.; Besedovsky, H.O.; Jessop, D.S. Re-exposure to endotoxin induces differential cytokine gene expression in the rat hypothalamus and spleen. Brain Behav. Immun. 2009, 23, 776–783. [Google Scholar]
- Ansari, J.A.; Ahmad, M.K.; Verma, A.K.; Fatima, N.; Khan, H.J.; Khan, A.R.; Mahdi, A.A. Microwave assisted determination of 17 minerals and four toxic metals in traditionally used medicinal plant Zingiber officinale Roscoe by Inductively Coupled Plasma-Optical Emission Spectrometer. Int. J. Adv. Res. 2015, 3, 879–887. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, S.N.; Charão, M.F.; Moro, A.M.; Roehrs, M.; Paniz, C.; Baierle, M.; Brucker, N.; Gioda, A.; Barbosa, F., Jr.; Bohrer, D.; et al. Evaluation of toxic metals and essential elements in children with learning disabilities from a rural area of southern Brazil. Int. J. Environ. Res. Public Health 2014, 11, 10806–10823. [Google Scholar] [CrossRef] [PubMed]
- Kordas, K.; Ardoino, G.; Coffman, D.L.; Queirolo, E.I.; Ciccariello, D.; Mañay, N.; Ettinger, A.S. Patterns of exposure to multiple metals and associations with neurodevelopment of preschool children from Montevideo, Uruguay. J. Environ. Public Health 2015, 2015, 493471. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.R.; Wu, H.; Bellinger, D.C.; Smith, D.R.; Wolff, M.S.; Savitz, D.A. Exposure to metal mixtures and neuropsychological functioning in middle childhood. Neurotoxicology 2022, 93, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Agency for Toxic Substances and Disease Registry (ASTDR), U.S. Department of Health and Human Sciences. Supplementary Guidance for Conducting Health Risk Assessment of Chemical Mixtures; Agency for Toxic Substances and Disease Registry (ASTDR), U.S. Department of Health and Human Sciences: Atlanta, GA, USA, 2000.
- Weiss, B. Lead, manganese, and methylmercury as risk factors for neurobehavioral impairment in advanced age. Int. J. Alzheimer’s Dis. 2010, 2011, 607543. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Finley, E.J.; Chakraborty, S.; Fretham, S.J.; Aschner, M. Cellular transport and homeostasis of essential and nonessential metals. Metallomics 2012, 4, 593–605. [Google Scholar] [CrossRef]
- Yokel, R.A. Manganese flux across the blood-brain barrier. Neuromol. Med. 2009, 11, 297–310. [Google Scholar] [CrossRef]
- Dong, X.; Ding, A.; Hu, H.; Xu, F.; Liu, L.; Wu, M. Placental Barrier on Cadmium Transfer from Mother to Fetus in Related to Pregnancy Complications. Int. J. Womens Health 2023, 10, 179–190. [Google Scholar] [CrossRef]
- Zheng, W.; Aschner, M.; Ghersi-Egeac, J.F. Brain barrier systems: A new frontier in metal neurotoxicological research. Toxicol. Appl. Pharmacol. 2003, 192, 1–11. [Google Scholar] [CrossRef]
- Xi, S.; Jin, Y.; Lv, X.; Sun, G. Distribution and speciation of arsenic by transplacental and early life exposure to inorganic arsenic in offspring rats. Biol. Trace Elem. Res. 2010, 134, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Andrade, V.; Mateus, M.L.; Santos, D.; Aschner, M.; Batoreu, M.C.; Marreilha dos Santos, A.P. Arsenic and manganese alter lead deposition in the rat. Biol. Trace Elem. Res. 2014, 158, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.H.; Qiu, Z.Q.; Zhang, L.; Shu, W.Q. Arsenite exposure altered the expression of NMDA receptor and postsynaptic signaling proteins in rat hippocampus. Toxicol. Lett. 2012, 211, 39–44. [Google Scholar] [CrossRef]
- Baddeley, A. Working memory. Science 1992, 255, 556–559. [Google Scholar] [CrossRef]
- Saxe, M.D.; Battaglia, F.; Wang, J.W.; Malleret, G.; David, D.J.; Monckton, J.E.; Garcia, A.D.; Sofroniew, M.V.; Kandel, E.R.; Santarelli, L.; et al. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc. Natl. Acad. Sci. USA 2006, 103, 17501–17506. [Google Scholar] [CrossRef]
- Chang, W.; Chen, J.; Wei, Q.Y.; Chen, X.M. Effects of Brn-3a protein and RNA expression in rat brain following low-level lead exposure during development on spatial learning and memory. Toxicol. Lett. 2006, 164, 63–70. [Google Scholar] [CrossRef]
- Martínez, L.; Jiménez, V.; García-Sepúlveda, C.; Ceballos, F.; Delgado, J.M.; Niño-Moreno, P.; Doniz, L.; Saavedra-Alanís, V.; Castillo, C.G.; Santoyo, M.E.; et al. Impact of early developmentalarsenic exposure on promotor CpG-island methylation of genes involved in neuronal plasticity. Neurochem. Int. 2011, 58, 574–581. [Google Scholar] [CrossRef]
- Luo, J.; Qiu, Z.; Chen, J.; Zhang, L.; Liu, W.; Tan, Y.; Shu, W. Maternal and early life arsenite exposure impairs neurodevelopment and increases the expression of PSA-NCAM in hippocampus of rat offspring. Toxicology 2013, 311, 99–106. [Google Scholar] [CrossRef]
- Liu, J.; Lewis, G. Environmental toxicity and poor cognitive outcomes in children and adults. J. Environ. Health 2014, 76, 130–138. [Google Scholar]
- Newcomer, J.W.; Farber, N.B.; Olney, J.W. NMDA receptor function, memory, and brain aging. Dialogues Clin. Neurosci. 2000, 2, 219–232. [Google Scholar] [CrossRef]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef]
- Monyer, H.; Burnashev, N.; Laurie, D.J.; Sakmann, B.; Seeburg, P.H. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron 1994, 12, 529–540. [Google Scholar] [CrossRef]
- Pizzi, M.; Consolandi, O.; Memo, M.; Spano, P.F. Activation of multiple metabotropic glutamate receptor subtypes prevents NMDA-induced excitotoxicity in rat hippocampal slices. Eur. J. Neurosci. 1996, 8, 1516–1521. [Google Scholar] [CrossRef]
- Scimemi, A.; Tian, H.; Diamond, J.S. Neuronal transporters regulate glutamate clearance, NMDA receptor activation, and synaptic plasticity in the hippocampus. J. Neurosci. 2009, 29, 14581–14595. [Google Scholar] [CrossRef]
- Kennedy, M.B. Synaptic Signaling in Learning and Memory. Cold Spring Harb. Perspect. Biol. 2013, 8, a016824. [Google Scholar] [CrossRef]
- Josselyn, S.A.; Nguyen, P.V. CREB, synapses and memory disorders: Past progress and future challenges. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 481–497. [Google Scholar] [CrossRef]
- Kandel, E.R. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef]
- Tauskela, J.S.; Fang, H.; Hewitt, M.; Brunette, E.; Ahuja, T.; Thivierge, J.P.; Comas, T.; Mealing, G.A. Elevated synaptic activity preconditions neurons against an in vitro model of ischemia. J. Biol. Chem. 2008, 283, 34667–34676. [Google Scholar] [CrossRef]
- Terasaki, Y.; Sasaki, T.; Yagita, Y.; Okazaki, S.; Sugiyama, Y.; Oyama, N.; Omura-Matsuoka, E.; Sakoda, S.; Kitagawa, K. Activation of NR2A receptors induces ischemic tolerance through CREB signaling. J. Cereb. Blood Flow. Metab. 2010, 30, 1441–1449. [Google Scholar] [CrossRef]
- Gardoni, F.; Caputi, A.; Cimino, M.; Pastorino, L.; Cattabeni, F.; Di Luca, M. Calcium/calmodulin-dependent protein kinase II is associated with NR2A/B subunits of NMDA receptor in postsynaptic densities. J. Neurochem. 1998, 71, 1733–1741. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Shinkawa, T.; Taoka, M.; Kobayashi, K.; Isobe, T.; Yamauchi, T. Identification of protein substrates of Ca2+/calmodulin-dependent protein kinase II in the postsynaptic density by protein sequencing and mass spectrometry. Biochem. Biophys. Res. Commun. 2002, 290, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.Z.; Xu, Z.; Ren, S.Q.; Hu, B.; Yao, W.; Wang, S.H.; Liu, S.Y.; Lu, W. Protein kinase C promotes N-methyl-D-aspartate (NMDA) receptor trafficking by indirectly triggering calcium/calmodulin-dependent protein kinase II (CaMKII) autophosphorylation. J. Biol. Chem. 2011, 286, 25187–25200. [Google Scholar] [CrossRef] [PubMed]
- Komiyama, N.H.; Watabe, A.M.; Carlisle, H.J.; Porter, K.; Charlesworth, P.; Monti, J.; Strathdee, D.J.; O’Carroll, C.M.; Martin, S.J.; Morris, R.G.; et al. SynGAP regulates ERK/MAPK signaling, synaptic plasticity, and learning in the complex with postsynaptic density 95 and NMDA receptor. J. Neurosci. 2002, 22, 9721–9732. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Laroche, S. Mitogen-activated protein kinase/extracellular regulated kinase signalling and memory stabilization: A review. Genes. Brain Behav. 2006, 5 (Suppl. S2), 61–72. [Google Scholar] [CrossRef]
- Chen, H.J.; Rojas-Soto, M.; Oguni, A.; Kennedy, M.B. A synaptic Ras-GTPase activating protein (p135 SynGAP) inhibited by CaM kinase II. Neuron 1998, 20, 895–904. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NR1 | FP:5′-TCCACCAAGAGCCCTTCGTG-3′ | RP; 5′-AGTTCAACAATCCGAAAAGCTGA-3′ |
| NR2A | FP:5′-GACTGGGACTACAGCCTG-3′ | RP; 5′-CTTCTCTGCCTGCCCATAGC-3′ |
| NR2B | FP:5′-GCCGGCAGCATTCCTACGACAC-3′ | RP; 5′-CCGGGGTTGTTGTGGTGGTGTC-3′ |
| GAPDH | FP:5′-ACGGGAAACCCATCACCAT-3′ | RP; 5′-CCAGCATCACCCCATTTGA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandravanshi, L.P.; Agrawal, P.; Darwish, H.W.; Trigun, S.K. Impairments of Spatial Memory and N-methyl-d-aspartate Receptors and Their Postsynaptic Signaling Molecules in the Hippocampus of Developing Rats Induced by As, Pb, and Mn Mixture Exposure. Brain Sci. 2023, 13, 1715. https://doi.org/10.3390/brainsci13121715
Chandravanshi LP, Agrawal P, Darwish HW, Trigun SK. Impairments of Spatial Memory and N-methyl-d-aspartate Receptors and Their Postsynaptic Signaling Molecules in the Hippocampus of Developing Rats Induced by As, Pb, and Mn Mixture Exposure. Brain Sciences. 2023; 13(12):1715. https://doi.org/10.3390/brainsci13121715
Chicago/Turabian StyleChandravanshi, Lalit P., Prashant Agrawal, Hany W. Darwish, and Surendra Kumar Trigun. 2023. "Impairments of Spatial Memory and N-methyl-d-aspartate Receptors and Their Postsynaptic Signaling Molecules in the Hippocampus of Developing Rats Induced by As, Pb, and Mn Mixture Exposure" Brain Sciences 13, no. 12: 1715. https://doi.org/10.3390/brainsci13121715