
What Does It Take to Play the Piano? Cognito-Motor Functions Underlying Motor Learning in Older Adults

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Cognitive Contributions of Motor Sequence Learning
1.2. Playing Music as a Special Type of Motor Sequence Learning
1.3. Transfer of Music Training beyond the Musical Domain
1.4. Aim of the Study
2. Materials and Methods
2.1. Participants
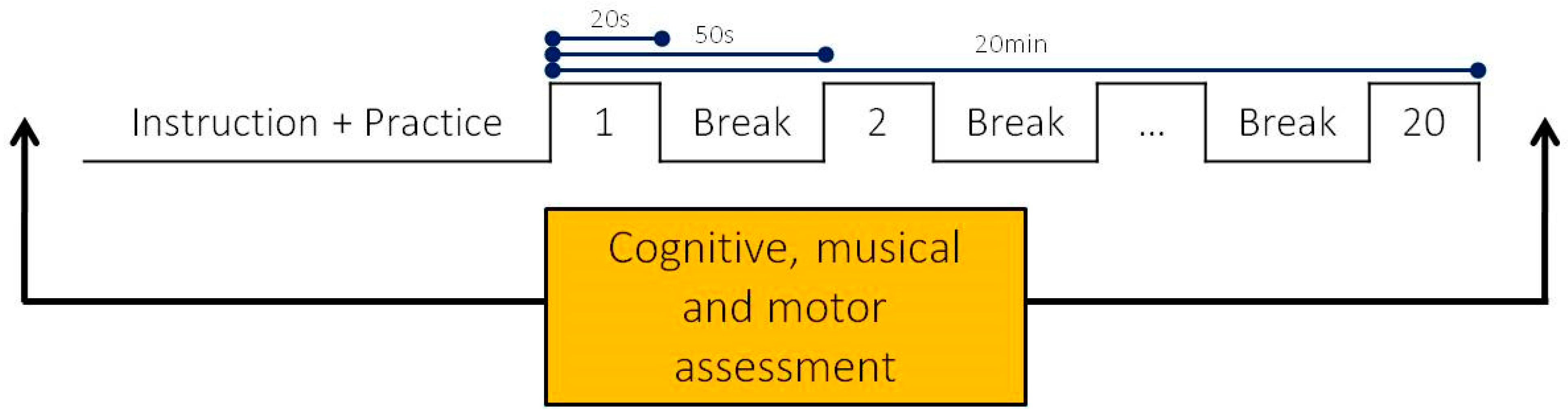
2.2. Procedure
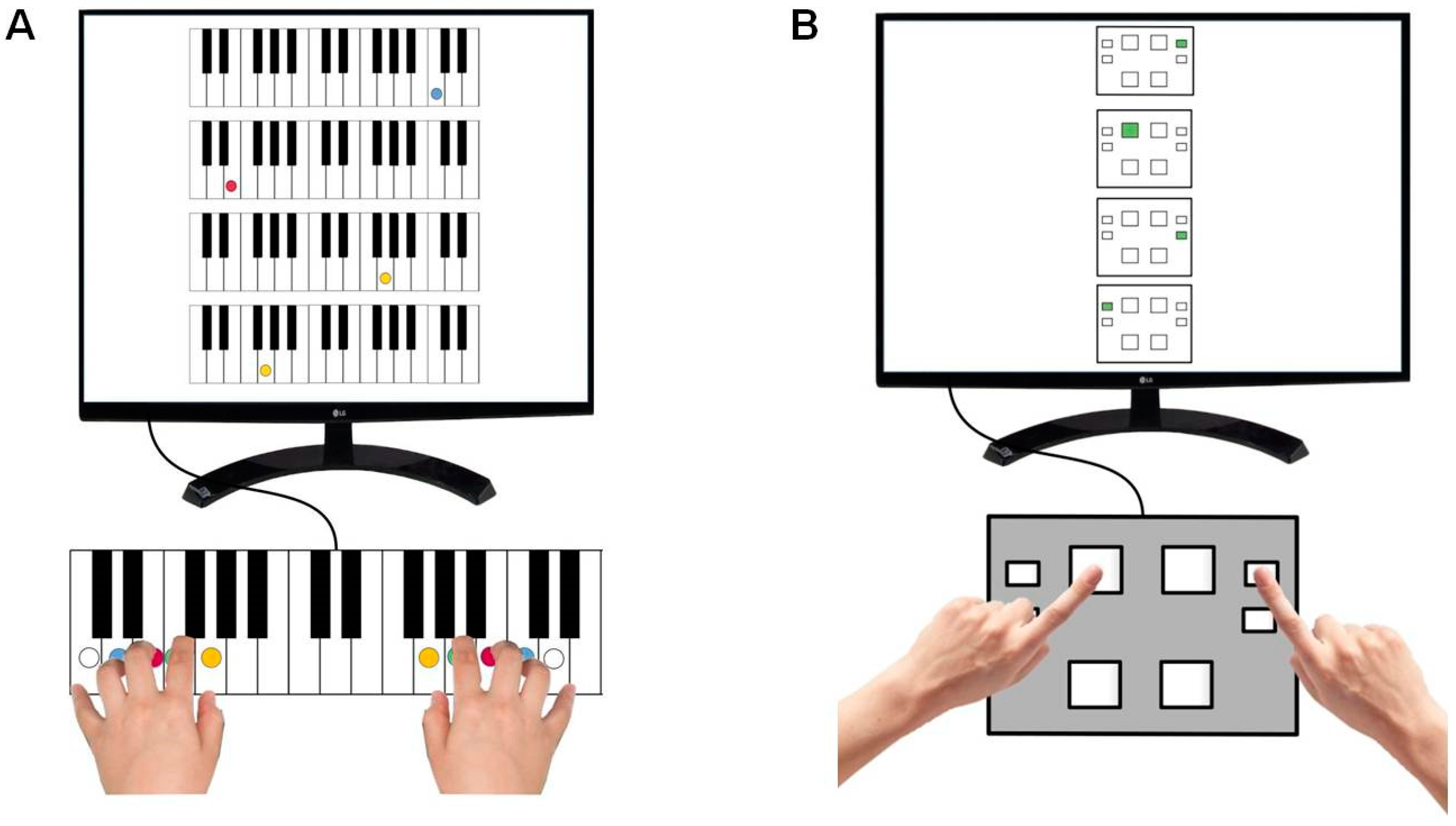
2.3. Motor Sequence Task
2.4. Gaze Behavior
2.5. Test-Battery
- Clicking speed
- Purdue Pegboard
- Coding
- Symbol Search
- Number Connection Test
- Reading letters
- Trail-Making-Test
- Digit Span
- Corsi Blocks
- Gold-MSI Questionnaire
- Piano experience questionnaire
2.6. Statistics
3. Results
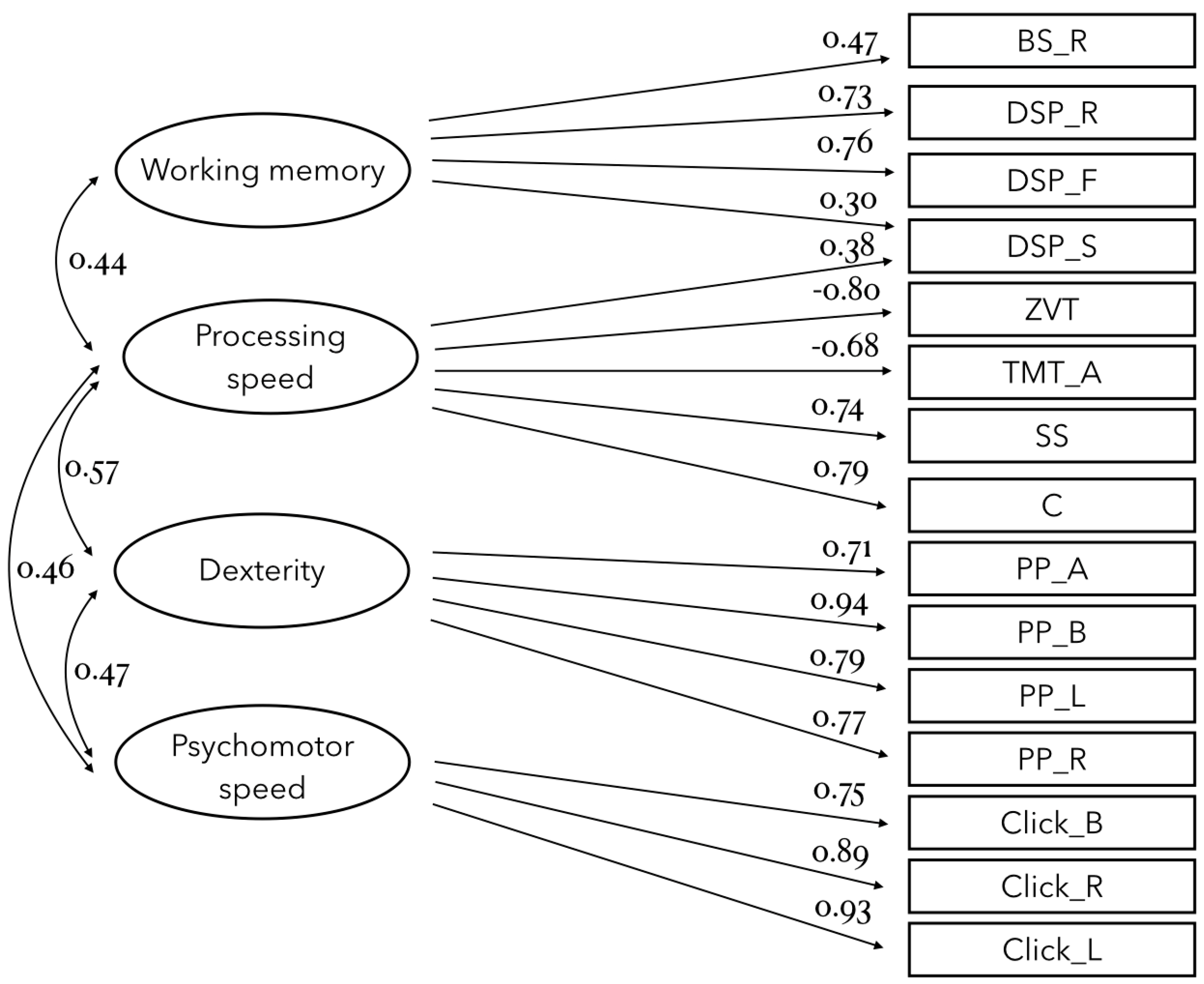
3.1. Latent Factor Analysis
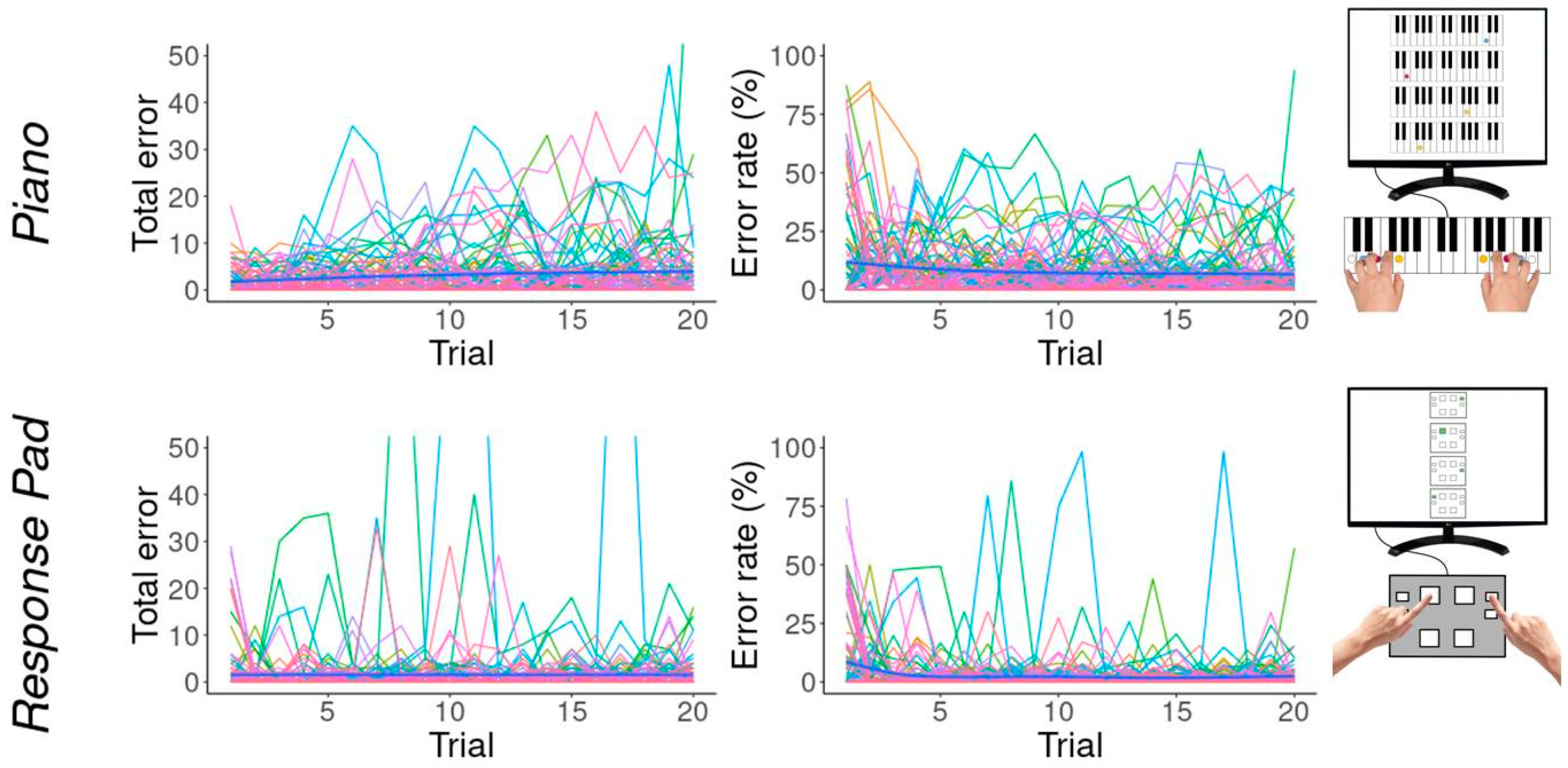
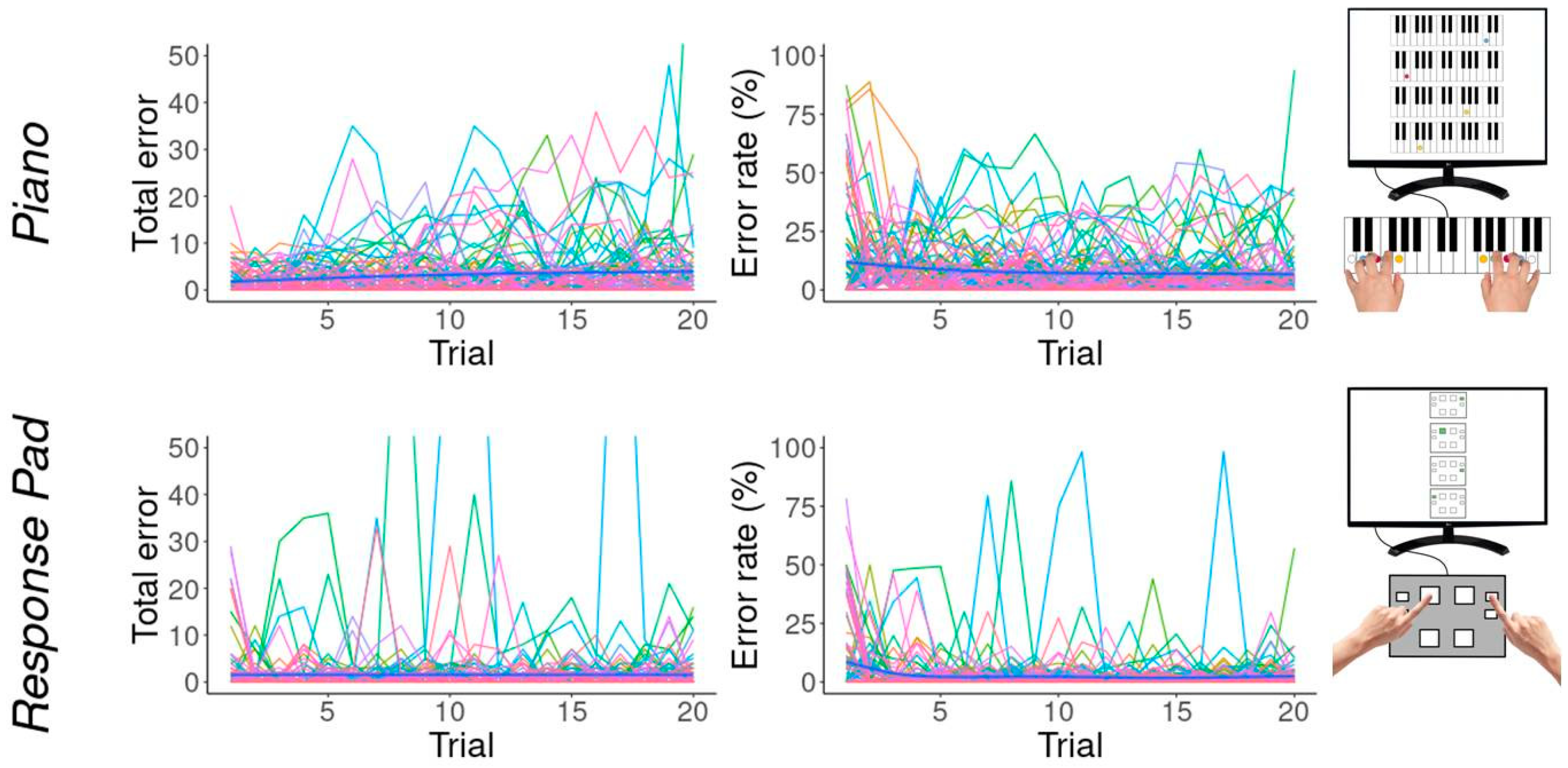
3.2. Errors
3.3. Gaze Behavior
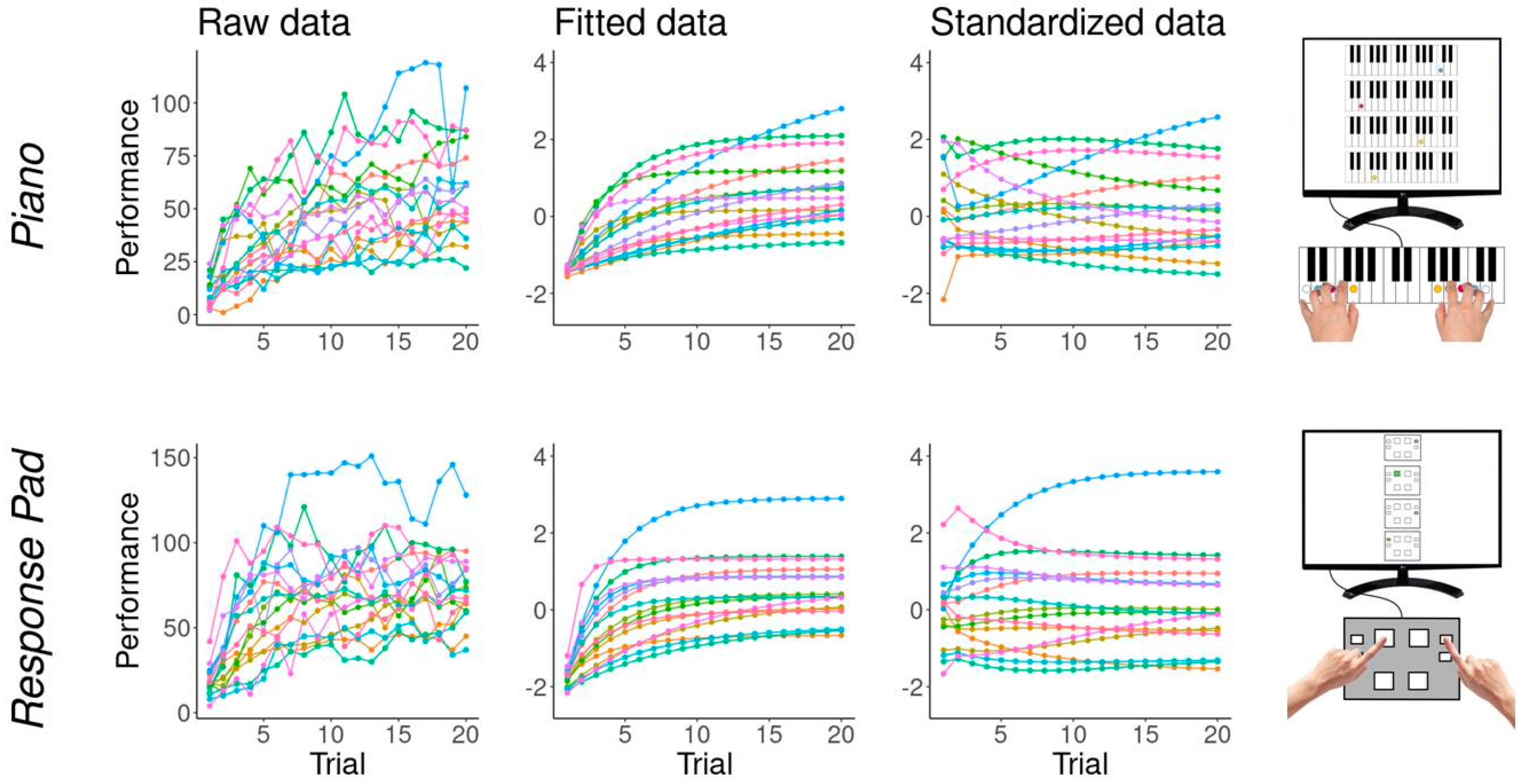
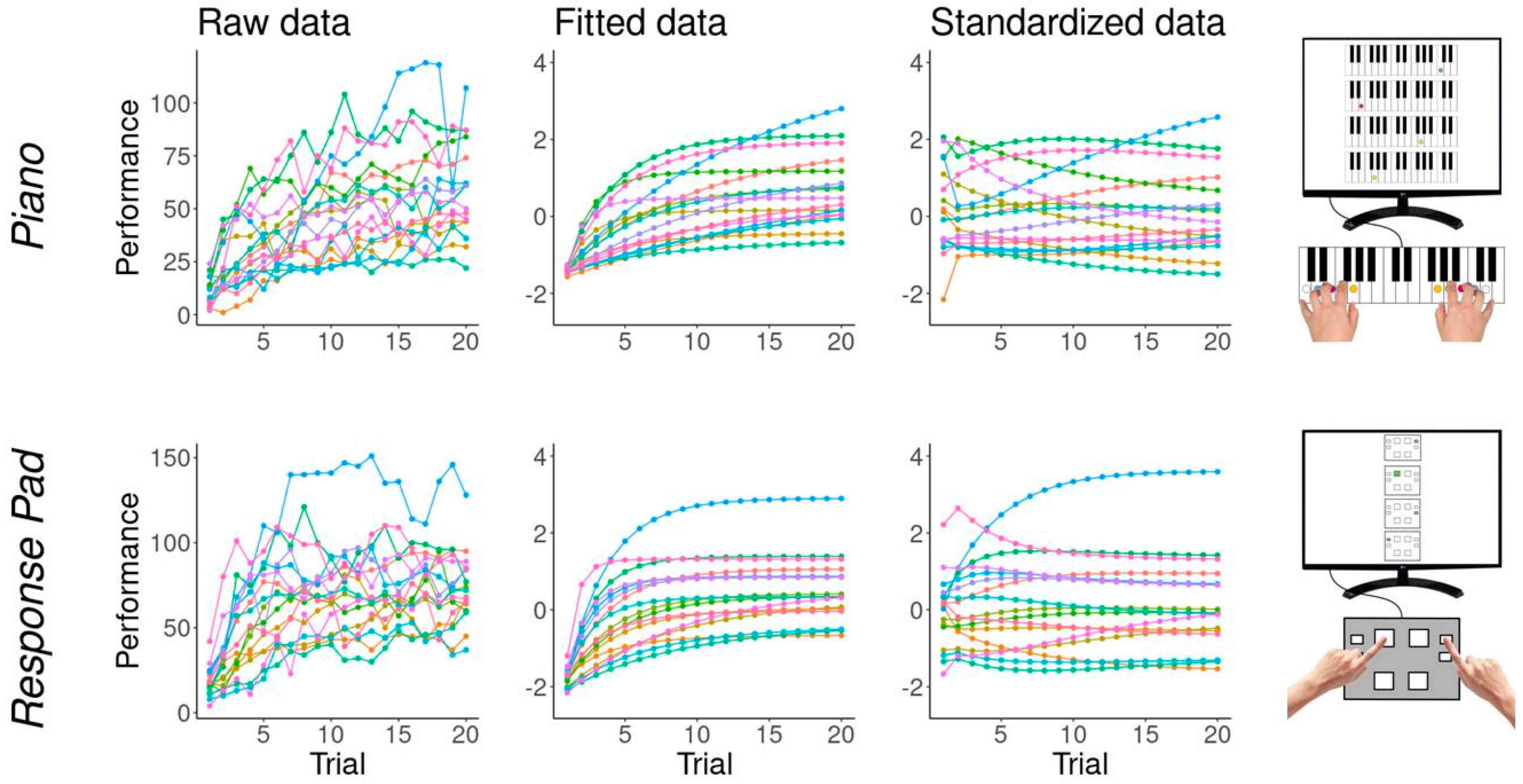
3.4. Motor Sequence Performance
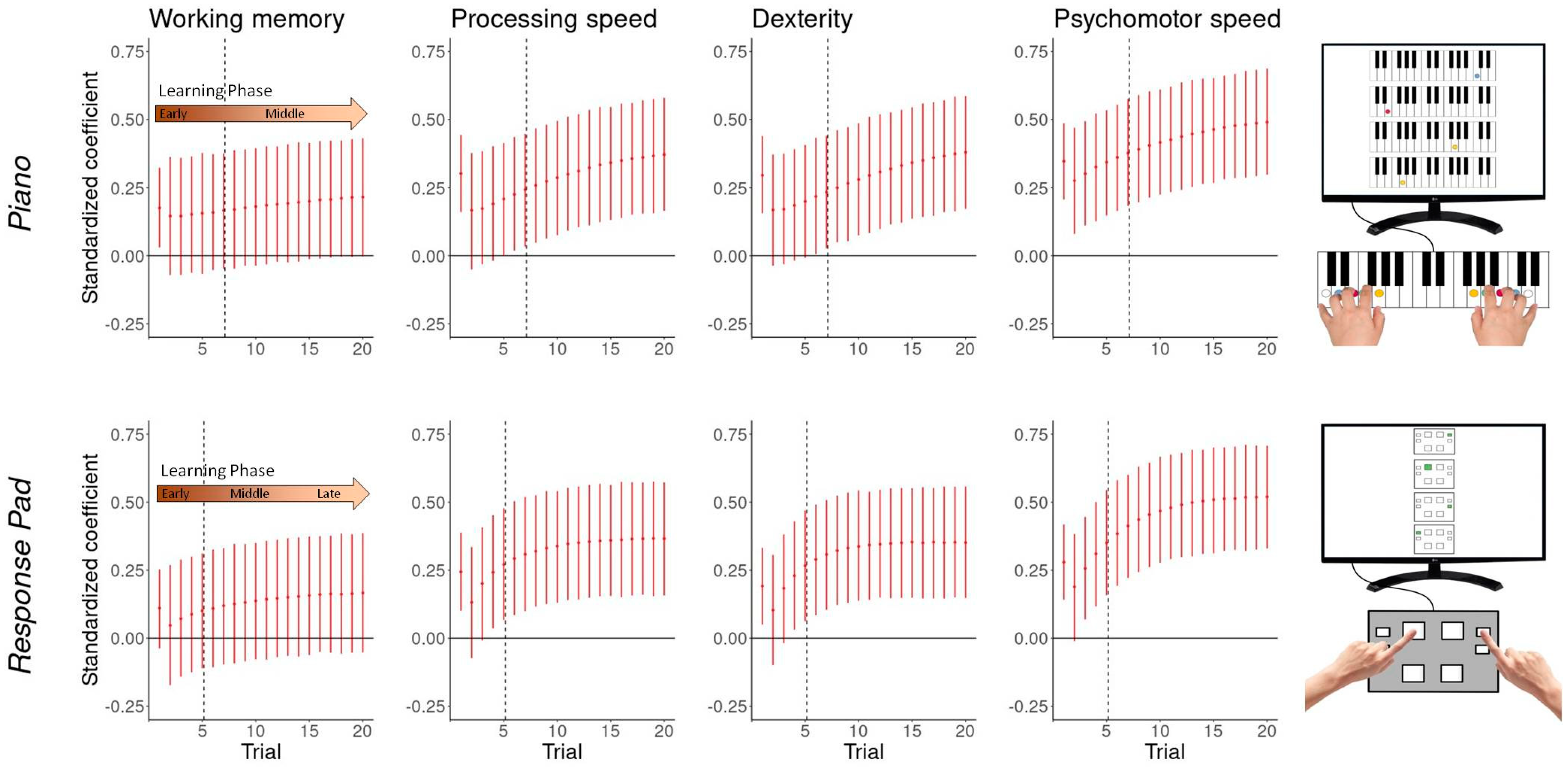
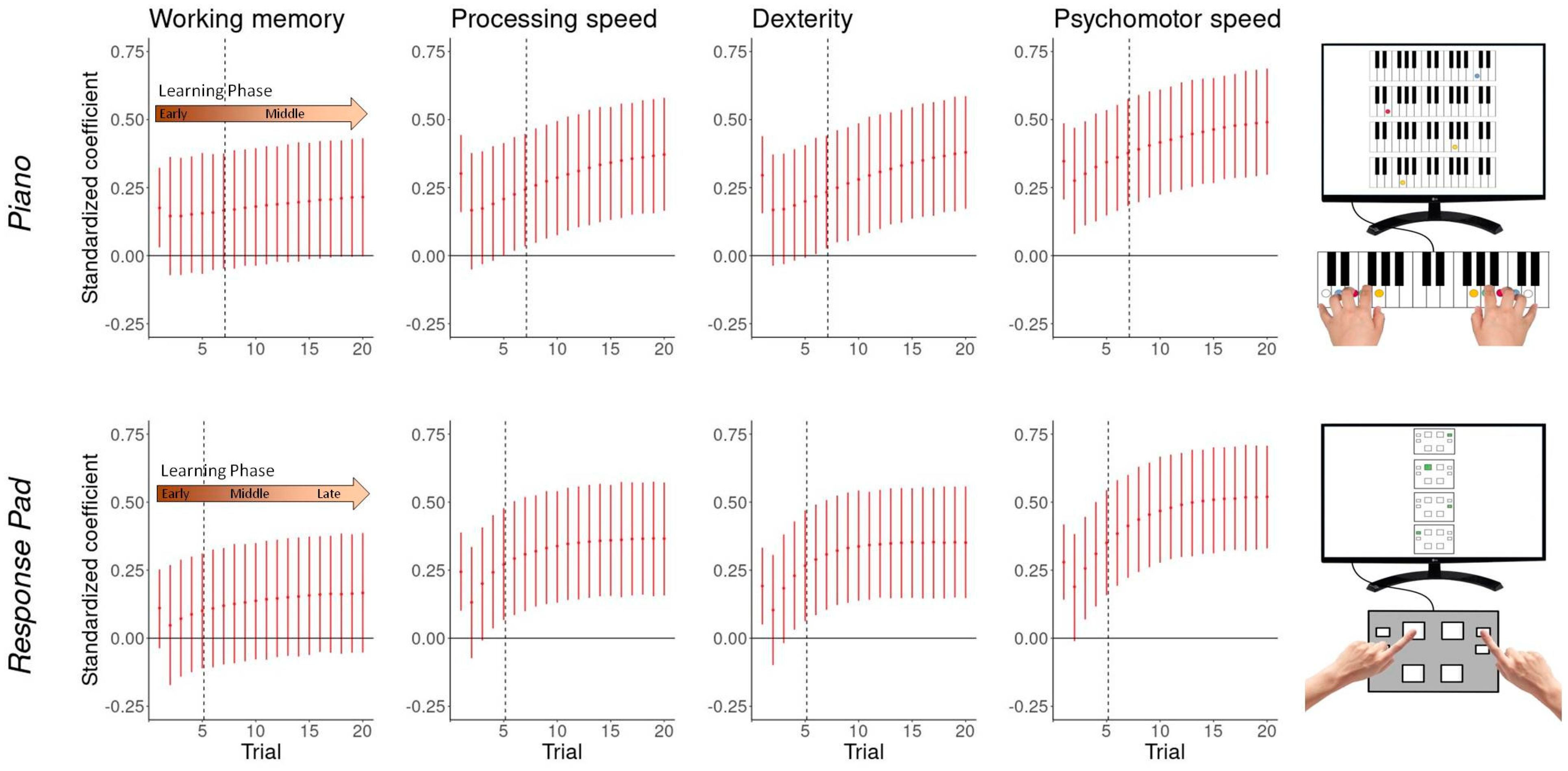
3.5. Performance-Cognito-Motor Relationship
3.6. Performance-Musicality Relationship
4. Conclusions
4.1. Implications for Interventions Promoting Healthy Aging
4.2. Strengths and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyon, J.; Benali, H. Reorganization and plasticity in the adult brain during learning of motor skills. Curr. Opin. Neurobiol. 2005, 15, 161–167. [Google Scholar] [CrossRef]
- Krakauer, J.W.; Hadjiosif, A.M.; Xu, J.; Wong, A.L.; Haith, A.M. Motor Learning. Compr. Physiol. 2019, 9, 613–663. [Google Scholar] [CrossRef]
- Goubault, E.; Verdugo, F.; Pelletier, J.; Traube, C.; Begon, M.; Dal Maso, F. Exhausting repetitive piano tasks lead to local forearm manifestation of muscle fatigue and negatively affect musical parameters. Sci. Rep. 2021, 11, 8117. [Google Scholar] [CrossRef]
- Doyon, J.; Bellec, P.; Amsel, R.; Penhune, V.; Monchi, O.; Carrier, J.; Lehéricy, S.; Benali, H. Contributions of the basal ganglia and functionally related brain structures to motor learning. Behav. Brain Res. 2009, 199, 61–75. [Google Scholar] [CrossRef]
- Doyon, J.; Gabitov, E.; Vahdat, S.; Lungu, O.; Boutin, A. Current issues related to motor sequence learning in humans. Curr. Opin. Behav. Sci. 2018, 20, 89–97. [Google Scholar] [CrossRef]
- Reybrouck, M.; Schiavio, A. Music performance as knowledge acquisition: A review and preliminary conceptual framework. Front. Psychol. 2024, 15, 1331806. [Google Scholar] [CrossRef]
- Ackerman, P.L. Determinants of individual differences during skill acquisition: Cognitive abilities and information processing. J. Exp. Psychol. Gen. 1988, 117, 288–318. [Google Scholar] [CrossRef]
- Fitts, P.M.; Posner, M.I. Human Performance; Brooks/Cole: Pacific Grove, CA, USA, 1967. [Google Scholar]
- Hikosaka, O.; Nakamura, K.; Sakai, K.; Nakahara, H. Central mechanisms of motor skill learning. Curr. Opin. Neurobiol. 2002, 12, 217–222. [Google Scholar] [CrossRef]
- Penhune, V.B.; Steele, C.J. Parallel contributions of cerebellar, striatal and M1 mechanisms to motor sequence learning. Behav. Brain Res. 2012, 226, 579–591. [Google Scholar] [CrossRef]
- Rasmussen, J. Skills, rules, and knowledge; signals, signs, and symbols, and other distinctions in human performance models. IEEE Trans. Syst. Man Cybern. 1983, SMC-13, 257–266. [Google Scholar] [CrossRef]
- Kim, J.W.; Ritter, F.E.; Koubek, R.J. An integrated theory for improved skill acquisition and retention in the three stages of learning. Theor. Issues Ergon. Sci. 2013, 14, 22–37. [Google Scholar] [CrossRef]
- Lindenberger, U.; Lövdén, M. Brain Plasticity in Human Lifespan Development: The Exploration–Selection–Refinement Model. Annu. Rev. Dev. Psychol. 2019, 1, 197–222. [Google Scholar] [CrossRef]
- Smith, E.T.; Skolasinska, P.; Qin, S.; Sun, A.; Fishwick, P.; Park, D.C.; Basak, C. Cognitive and structural predictors of novel task learning, and contextual predictors of time series of daily task performance during the learning period. Front. Aging Neurosci. 2022, 14, 936528. [Google Scholar] [CrossRef]
- Verwey, W.B. C-SMB 2.0: Integrating over 25 years of motor sequencing research with the Discrete Sequence Production task. Psychon. Bull. Rev. 2023. [Google Scholar] [CrossRef]
- Ackerman, P.L.; Cianciolo, A.T. Cognitive, perceptual-speed, and psychomotor determinants of individual differences during skill acquisition. J. Exp. Psychol. Appl. 2000, 6, 259–290. [Google Scholar] [CrossRef]
- Guadagnoli, M.A.; Lee, T.D. Challenge point: A framework for conceptualizing the effects of various practice conditions in motor learning. J. Mot. Behav. 2004, 36, 212–224. [Google Scholar] [CrossRef]
- Csikszentmihalyi, M.; Abuhamdeh, S.; Nakamura, J. Flow. In Flow and the Foundations of Positive Psychology: The Collected Works of Mihaly Csikszentmihalyi; Csikszentmihalyi, M., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 227–238. ISBN 978-94-017-9088-8. [Google Scholar]
- Sinnamon, S.; Moran, A.; O’Connell, M. Flow Among Musicians. J. Res. Music Educ. 2012, 60, 6–25. [Google Scholar] [CrossRef]
- Antonini Philippe, R.; Kosirnik, C.; Ortuño, E.; Biasutti, M. Flow and music performance: Professional musicians and music students’ views. Psychol. Music 2022, 50, 1023–1038. [Google Scholar] [CrossRef]
- Ackerman, P.L. Predicting individual differences in complex skill acquisition: Dynamics of ability determinants. J. Appl. Psychol. 1992, 77, 598–614. [Google Scholar] [CrossRef]
- Fisk, A.D.; Ackerman, P.L.; Schneider, W. 5. Automatic and Controlled Processing Theory and its Applications to Human Factors Problems. In Human Factors Psychology; Elsevier: Amsterdam, The Netherlands, 1987; pp. 159–197. ISBN 9780444703194. [Google Scholar]
- Voelkle, M.C.; Wittmann, W.W.; Ackerman, P.L. Abilities and skill acquisition: A latent growth curve approach. Learn. Individ. Differ. 2006, 16, 303–319. [Google Scholar] [CrossRef]
- Ackerman, P.L. Ability Determinants of Individual Differences in Skilled Performance. In Cognition and Intelligence: Identifying the Mechanisms of the Mind; Transferred to Digital Print; Sternberg, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2010; pp. 142–159. ISBN 9780521827447. [Google Scholar]
- Shinar, D.; Meir, M.; Ben-Shoham, I. How automatic is manual gear shifting? Hum. Factors 1998, 40, 647–654. [Google Scholar] [CrossRef]
- Herrebrøden, H.; Jensenius, A.R.; Espeseth, T.; Bishop, L.; Vuoskoski, J.K. Cognitive load causes kinematic changes in both elite and non-elite rowers. Hum. Mov. Sci. 2023, 90, 103113. [Google Scholar] [CrossRef] [PubMed]
- Monz, A.; Morbe, K.; Klein, M.; Schaefer, S. Mutual interference between memory encoding and motor skills: The influence of motor expertise. Front. Psychol. 2023, 14, 1196978. [Google Scholar] [CrossRef] [PubMed]
- Peel, N.M.; Alapatt, L.J.; Jones, L.V.; Hubbard, R.E. The Association Between Gait Speed and Cognitive Status in Community-Dwelling Older People: A Systematic Review and Meta-analysis. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Beaunieux, H.; Hubert, V.; Witkowski, T.; Pitel, A.-L.; Rossi, S.; Danion, J.-M.; Desgranges, B.; Eustache, F. Which processes are involved in cognitive procedural learning? Memory 2006, 14, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, J.A.; Taylor, A.E.; Lang, A.E. Procedural learning and neostriatal dysfunction in man. Brain 1988, 111 Pt 4, 941–959. [Google Scholar] [CrossRef] [PubMed]
- Cormier, D.C.; McGrew, K.S.; Bulut, O.; Funamoto, A. Revisiting the Relations Between the WJ-IV Measures of Cattell-Horn-Carroll (CHC) Cognitive Abilities and Reading Achievement During the School-Age Years. J. Psychoeduc. Assess. 2017, 35, 731–754. [Google Scholar] [CrossRef]
- Seidler, R.D.; Bo, J.; Anguera, J.A. Neurocognitive contributions to motor skill learning: The role of working memory. J. Mot. Behav. 2012, 44, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Worschech, F.; James, C.E.; Jünemann, K.; Sinke, C.; Krüger, T.H.C.; Scholz, D.S.; Kliegel, M.; Marie, D.; Altenmüller, E. Fine motor control improves in older adults after 1 year of piano lessons: Analysis of individual development and its coupling with cognition and brain structure. Eur. J. Neurosci. 2023, 57, 2040–2061. [Google Scholar] [CrossRef] [PubMed]
- Cormier, D.C.; Bulut, O.; McGrew, K.S.; Frison, J. The role of Cattell–Horn–Carroll (CHC) cognitive abilities in predicting writing achievement during the school-age years. Psychol. Schs. 2016, 53, 787–803. [Google Scholar] [CrossRef]
- Altenmüller, E.; Furuya, S. Apollos Gift and Curse: Making Music as a model for Adaptive and Maladaptive Plasticity. e-Neuroforum 2017, 23, 57–75. [Google Scholar] [CrossRef]
- Jabusch, H.-C.; Alpers, H.; Kopiez, R.; Vauth, H.; Altenmüller, E. The influence of practice on the development of motor skills in pianists: A longitudinal study in a selected motor task. Hum. Mov. Sci. 2009, 28, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Worschech, F.; Großbach, M.; Bläsing, B.; Altenmüller, E. Elevated Forearm Coactivation Levels and Higher Temporal Variability in String Players with Musicians’ Dystonia During Demanding Playing Conditions. Med. Probl. Perform. Art. 2020, 35, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Bugos, J.A.; Perlstein, W.M.; McCrae, C.S.; Brophy, T.S.; Bedenbaugh, P.H. Individualized piano instruction enhances executive functioning and working memory in older adults. Aging Ment. Health 2007, 11, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Degé, F.; Kerkovius, K. The effects of drumming on working memory in older adults. Ann. N. Y. Acad. Sci. 2018, 1423, 242–250. [Google Scholar] [CrossRef]
- Frischen, U.; Schwarzer, G.; Degé, F. Music lessons enhance executive functions in 6- to 7-year-old children. Learn. Instr. 2021, 74, 101442. [Google Scholar] [CrossRef]
- Burgoyne, A.P.; Harris, L.J.; Hambrick, D.Z. Predicting piano skill acquisition in beginners: The role of general intelligence, music aptitude, and mindset. Intelligence 2019, 76, 101383. [Google Scholar] [CrossRef]
- Kopiez, R.; Lee, J.I. Towards a general model of skills involved in sight reading music. Music Educ. Res. 2008, 10, 41–62. [Google Scholar] [CrossRef]
- Toril, P.; Reales, J.M.; Ballesteros, S. Video game training enhances cognition of older adults: A meta-analytic study. Psychol. Aging 2014, 29, 706–716. [Google Scholar] [CrossRef]
- Stojan, R.; Voelcker-Rehage, C. A Systematic Review on the Cognitive Benefits and Neurophysiological Correlates of Exergaming in Healthy Older Adults. J. Clin. Med. 2019, 8, 734. [Google Scholar] [CrossRef]
- Wayne, P.M.; Walsh, J.N.; Taylor-Piliae, R.E.; Wells, R.E.; Papp, K.V.; Donovan, N.J.; Yeh, G.Y. Effect of tai chi on cognitive performance in older adults: Systematic review and meta-analysis. J. Am. Geriatr. Soc. 2014, 62, 25–39. [Google Scholar] [CrossRef]
- Ludyga, S.; Gerber, M.; Pühse, U.; Looser, V.N.; Kamijo, K. Systematic review and meta-analysis investigating moderators of long-term effects of exercise on cognition in healthy individuals. Nat. Hum. Behav. 2020, 4, 603–612. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Risk Reduction of Cognitive Decline and Dementia: WHO Guidelines; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-155054-3. [Google Scholar]
- Román-Caballero, R.; Vadillo, M.A.; Trainor, L.J.; Lupiáñez, J. Please don’t stop the music: A meta-analysis of the cognitive and academic benefits of instrumental musical training in childhood and adolescence. Educ. Res. Rev. 2022, 35, 100436. [Google Scholar] [CrossRef]
- Sala, G.; Gobet, F. Does Far Transfer Exist? Negative Evidence from Chess, Music, and Working Memory Training. Curr. Dir. Psychol. Sci. 2017, 26, 515–520. [Google Scholar] [CrossRef]
- Sala, G.; Gobet, F. Cognitive and academic benefits of music training with children: A multilevel meta-analysis. Mem. Cognit. 2020, 48, 1429–1441. [Google Scholar] [CrossRef]
- Román-Caballero, R.; Arnedo, M.; Triviño, M.; Lupiáñez, J. Musical practice as an enhancer of cognitive function in healthy aging—A systematic review and meta-analysis. PLoS ONE 2018, 13, e0207957. [Google Scholar] [CrossRef] [PubMed]
- Sala, G.; Aksayli, N.D.; Tatlidil, K.S.; Tatsumi, T.; Gondo, Y.; Gobet, F. Near and Far Transfer in Cognitive Training: A Second-Order Meta-Analysis. Collabra Psychol. 2019, 5, 18. [Google Scholar] [CrossRef]
- Thorndike, E.L.; Woodworth, R.S. The influence of improvement in one mental function upon the efficiency of other functions. II. The estimation of magnitudes. Psychol. Rev. 1901, 8, 384–395. [Google Scholar] [CrossRef]
- Barnett, S.M.; Ceci, S.J. When and where do we apply what we learn? A taxonomy for far transfer. Psychol. Bull. 2002, 128, 612–637. [Google Scholar] [CrossRef]
- Birren, J.E.; Fisher, L.M. Aging and speed of behavior: Possible consequences for psychological functioning. Annu. Rev. Psychol. 1995, 46, 329–353. [Google Scholar] [CrossRef]
- Salthouse, T.A. The processing-speed theory of adult age differences in cognition. Psychol. Rev. 1996, 103, 403–428. [Google Scholar] [CrossRef]
- Müllensiefen, D.; Gingras, B.; Musil, J.; Stewart, L. The musicality of non-musicians: An index for assessing musical sophistication in the general population. PLoS ONE 2014, 9, e89642. [Google Scholar] [CrossRef] [PubMed]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in behavior made easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Tiffin, J.; Asher, E.J. The Purdue pegboard; norms and studies of reliability and validity. J. Appl. Psychol. 1948, 32, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, D. Wechsler Adult Intelligence Scale: Fourth Edition (WAIS-IV); [Assessment Instrument]; Pearson: San Antonio, TX, USA, 2008. [Google Scholar]
- Oswald, W.D. Zahlen-Verbindungs-Test ZVT.: 3., Überarbeitete und neu Normierte Auflage; Hogrefe Verlag GmbH & Co. KG: Göttingen, Germany, 2016. [Google Scholar]
- Lehrl, S.; Blaha, L. Messung des Arbeitsgedächtnisses: KAI-N; [Manual]; Vless: Ebersberg, Germany, 2000; ISBN 3-88562-091-x. [Google Scholar]
- Reitan, R.M.; Wolfson, D. The Halstead-Reitan Neuropsychological Test Battery: Theory and Clinical Interpretation, 2nd ed.; Neuropsychology Press: Tucson, Arizona, 1993; ISBN 093451514X. [Google Scholar]
- Milner, B. Interhemispheric differences in the localization of psychological processes in man. Br. Med. Bull. 1971, 27, 272–277. [Google Scholar] [CrossRef]
- Müller, R.A.; Kleinhans, N.; Pierce, K.; Kemmotsu, N.; Courchesne, E. Functional MRI of motor sequence acquisition: Effects of learning stage and performance. Brain Res. Cogn. Brain Res. 2002, 14, 277–293. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Soft. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Bürkner, P.-C. brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Soft. 2017, 80, 1–28. [Google Scholar] [CrossRef]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. performance: An R Package for Assessment, Comparison and Testing of Statistical Models. JOSS 2021, 6, 3139. [Google Scholar] [CrossRef]
- Heathcote, A.; Brown, S.; Mewhort, D.J. The power law repealed: The case for an exponential law of practice. Psychon. Bull. Rev. 2000, 7, 185–207. [Google Scholar] [CrossRef]
- Censor, N. Generalization of perceptual and motor learning: A causal link with memory encoding and consolidation? Neuroscience 2013, 250, 201–207. [Google Scholar] [CrossRef]
- Verwey, W.B.; Shea, C.H.; Wright, D.L. A cognitive framework for explaining serial processing and sequence execution strategies. Psychon. Bull. Rev. 2015, 22, 54–77. [Google Scholar] [CrossRef] [PubMed]
- Verwey, W.B.; Abrahamse, E.L.; Kleine, E.d. Cognitive processing in new and practiced discrete keying sequences. Front. Psychol. 2010, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Schwizer Ashkenazi, S.; Raiter-Avni, R.; Vakil, E. The benefit of assessing implicit sequence learning in pianists with an eye-tracked serial reaction time task. Psychol. Res. 2022, 86, 1426–1441. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, B.; van der Kamp, J.; Verneau, M.; Jongbloed-Pereboom, M.; Masters, R.S.W. Implicit and explicit learning: Applications from basic research to sports for individuals with impaired movement dynamics. Disabil. Rehabil. 2010, 32, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Slusarz, P.; Terry, C. The interaction of the explicit and the implicit in skill learning: A dual-process approach. Psychol. Rev. 2005, 112, 159–192. [Google Scholar] [CrossRef] [PubMed]
- Landau, S.M.; D’esposito, M. Sequence learning in pianists and nonpianists: An fMRI study of motor expertise. Cogn. Affect. Behav. Neurosci. 2006, 6, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Bugos, J.A. The benefits of music instruction on processing speed, verbal fluency, and cognitive control in aging. Music Educ. Res. Int. 2010, 4, 1–9. [Google Scholar]
- Bugos, J.A. The Effects of Bimanual Coordination in Music Interventions on Executive Functions in Aging Adults. Front. Integr. Neurosci. 2019, 13, 68. [Google Scholar] [CrossRef]
- Yeon, J.; Larson, A.S.; Rahnev, D.; D’esposito, M. Task learning is subserved by a domain-general brain network. Cereb. Cortex 2024, 34, bhae013. [Google Scholar] [CrossRef]
- Osman, A.M.; Jaffe, P.I.; Ng, N.F.; Kerlan, K.R.; Schafer, R.J. Transfer of learning: Analysis of dose-response functions from a large-scale, online, cognitive training dataset. PLoS ONE 2023, 18, e0281095. [Google Scholar] [CrossRef]
- Gold, C.; Solli, H.P.; Krüger, V.; Lie, S.A. Dose-response relationship in music therapy for people with serious mental disorders: Systematic review and meta-analysis. Clin. Psychol. Rev. 2009, 29, 193–207. [Google Scholar] [CrossRef]
- Ackerman, P.L. A correlational analysis of skill specificity: Learning, abilities, and individual differences. J. Exp. Psychol. Learn. Mem. Cogn. 1990, 16, 883–901. [Google Scholar] [CrossRef]
- Hamada, H.; Wen, W.; Kawasaki, T.; Yamashita, A.; Asama, H. Characteristics of EEG power spectra involved in the proficiency of motor learning. Front. Neurosci. 2023, 17, 1094658. [Google Scholar] [CrossRef]
- Noack, H.; Lövdén, M.; Schmiedek, F. On the validity and generality of transfer effects in cognitive training research. Psychol. Res. 2014, 78, 773–789. [Google Scholar] [CrossRef]
- McGrew, K.S. CHC theory and the human cognitive abilities project: Standing on the shoulders of the giants of psychometric intelligence research. Intelligence 2009, 37, 1–10. [Google Scholar] [CrossRef]
- Ericsson, K.A.; Krampe, R.T.; Tesch-Römer, C. The role of deliberate practice in the acquisition of expert performance. Psychol. Rev. 1993, 100, 363–406. [Google Scholar] [CrossRef]
- Schmid, D.; Erlacher, D.; Klostermann, A.; Kredel, R.; Hossner, E.-J. Sleep-dependent motor memory consolidation in healthy adults: A meta-analysis. Neurosci. Biobehav. Rev. 2020, 118, 270–281. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Worschech, F.; Passarotto, E.; Losch, H.; Oku, T.; Lee, A.; Altenmüller, E. What Does It Take to Play the Piano? Cognito-Motor Functions Underlying Motor Learning in Older Adults. Brain Sci. 2024, 14, 405. https://doi.org/10.3390/brainsci14040405
Worschech F, Passarotto E, Losch H, Oku T, Lee A, Altenmüller E. What Does It Take to Play the Piano? Cognito-Motor Functions Underlying Motor Learning in Older Adults. Brain Sciences. 2024; 14(4):405. https://doi.org/10.3390/brainsci14040405
Chicago/Turabian StyleWorschech, Florian, Edoardo Passarotto, Hannah Losch, Takanori Oku, André Lee, and Eckart Altenmüller. 2024. "What Does It Take to Play the Piano? Cognito-Motor Functions Underlying Motor Learning in Older Adults" Brain Sciences 14, no. 4: 405. https://doi.org/10.3390/brainsci14040405