
Abstinence and Fear Experienced during This Period Produce Distinct Cortical and Hippocampal Adaptations in Alcohol-Dependent Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Chronic Intermittent Ethanol Vapor Exposure (CIE)
2.3. Trace Fear Conditioning
2.4. Western Blotting
2.5. Golgi–Cox Staining and Neuron Morphology Analysis
2.6. Statistical Analyses
3. Results

3.1. Abstinence Differentially Effects the Expression of Synaptic Plasticity-Associated Proteins over the Time Course in the mPFC and the DG

3.2. Freezing during Baseline Session
3.3. Freezing Behavior during Acquisition of TFC
3.4. Freezing Behavior during Context Retrieval and during CS Retrieval in Fear Context
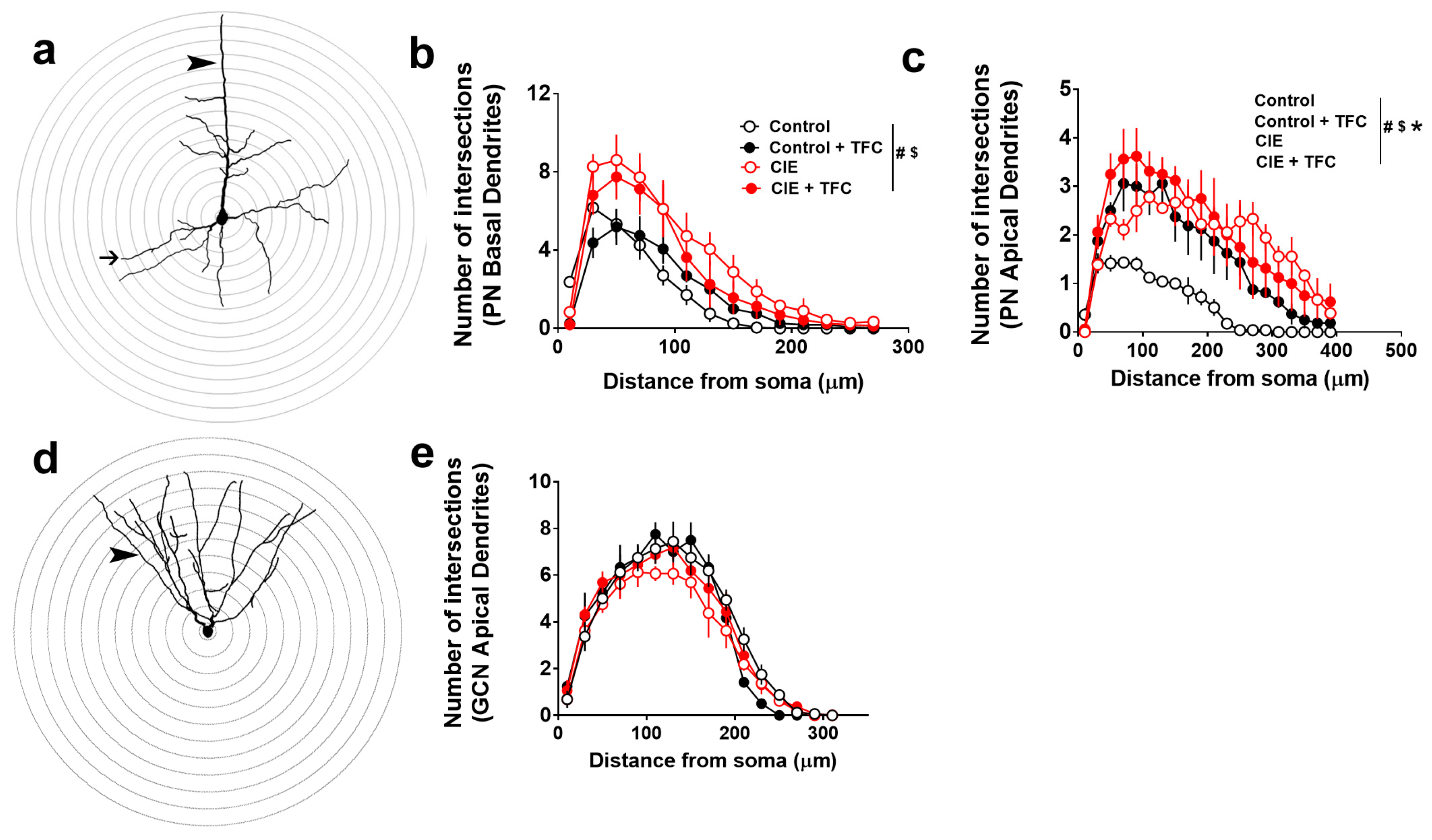
3.5. Dendritic Arborization of Pyramidal Neurons in the mPFC and GCNs in the DG

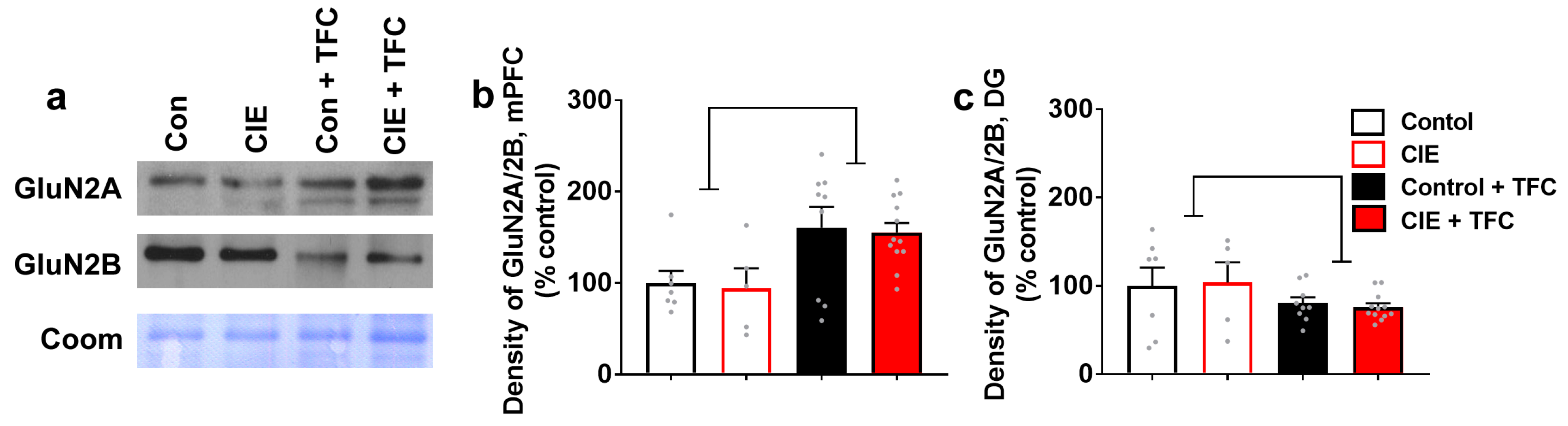
3.6. TFC Differentially Affects the Expression of GluN2A/2B Ratio in the mPFC and the DG

4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brandt, J.; Butters, N.; Ryan, C.; Bayog, R. Cognitive loss and recovery in long-term alcohol abusers. Arch. Gen. Psychiatry 1983, 40, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Glenn, S.W.; Parsons, O.A. Impaired efficiency in female alcoholics’ neuropsychological performance. J. Clin. Exp. Neuropsychol. 1991, 13, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, E.V.; Rosenbloom, M.J.; Lim, K.O.; Pfefferbaum, A. Longitudinal changes in cognition, gait, and balance in abstinent and relapsed alcoholic men: Relationships to changes in brain structure. Neuropsychology 2000, 14, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Bechara, A.; Dolan, S.; Denburg, N.; Hindes, A.; Anderson, S.W.; Nathan, P.E. Decision-making deficits, linked to a dysfunctional ventromedial prefrontal cortex, revealed in alcohol and stimulant abusers. Neuropsychologia 2001, 39, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Oscar-Berman, M.; Marinkovic, K. Alcoholism and the brain: An overview. Alcohol. Res. Health 2003, 27, 125–133. [Google Scholar] [PubMed]
- Rosenbloom, M.; Sullivan, E.V.; Pfefferbaum, A. Using magnetic resonance imaging and diffusion tensor imaging to assess brain damage in alcoholics. Alcohol. Res. Health 2003, 27, 146–152. [Google Scholar] [PubMed]
- Sullivan, E.V.; Rosenbloom, M.J.; Pfefferbaum, A. Pattern of motor and cognitive deficits in detoxified alcoholic men. Alcohol. Clin. Exp. Res. 2000, 24, 611–621. [Google Scholar] [CrossRef] [PubMed]
- De Bellis, M.D.; Narasimhan, A.; Thatcher, D.L.; Keshavan, M.S.; Soloff, P.; Clark, D.B. Prefrontal cortex, thalamus, and cerebellar volumes in adolescents and young adults with adolescent-onset alcohol use disorders and comorbid mental disorders. Alcohol. Clin. Exp. Res. 2005, 29, 1590–1600. [Google Scholar] [CrossRef] [PubMed]
- Fryer, S.L.; Jorgensen, K.W.; Yetter, E.J.; Daurignac, E.C.; Watson, T.D.; Shanbhag, H.; Krystal, J.H.; Mathalon, D.H. Differential brain response to alcohol cue distractors across stages of alcohol dependence. Biol. Psychol. 2013, 92, 282–291. [Google Scholar] [CrossRef]
- Sullivan, E.V.; Marsh, L.; Mathalon, D.H.; Lim, K.O.; Pfefferbaum, A. Anterior hippocampal volume deficits in nonamnesic, aging chronic alcoholics. Alcohol. Clin. Exp. Res. 1995, 19, 110–122. [Google Scholar] [CrossRef]
- Bengochea, O.; Gonzalo, L.M. Effect of chronic alcoholism on the human hippocampus. Histol. Histopathol. 1990, 5, 349–357. [Google Scholar] [PubMed]
- Durazzo, T.C.; Tosun, D.; Buckley, S.; Gazdzinski, S.; Mon, A.; Fryer, S.L.; Meyerhoff, D.J. Cortical Thickness, Surface Area, and Volume of the Brain Reward System in Alcohol Dependence: Relationships to Relapse and Extended Abstinence. Alcohol. Clin. Exp. Res. 2011, 35, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- UNODC. World Drug Report 2017; United Nations: Vienna, Austria, 2017; ISBN 978-92-1-148291-1. eISBN 978-92-1-060623-3. [Google Scholar]
- De Santis, S.; Bach, P.; Perez-Cervera, L.; Cosa-Linan, A.; Weil, G.; Vollstadt-Klein, S.; Hermann, D.; Kiefer, F.; Kirsch, P.; Ciccocioppo, R.; et al. Microstructural White Matter Alterations in Men With Alcohol Use Disorder and Rats With Excessive Alcohol Consumption During Early Abstinence. JAMA Psychiatry 2019, 76, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Somkuwar, S.S.; Fannon, M.J.; Staples, M.C.; Zamora-Martinez, E.R.; Navarro, A.I.; Kim, A.; Quigley, J.A.; Edwards, S.; Mandyam, C.D. Alcohol dependence-induced regulation of the proliferation and survival of adult brain progenitors is associated with altered BDNF-TrkB signaling. Brain Struct. Funct. 2016, 221, 4319–4335. [Google Scholar] [CrossRef] [PubMed]
- Avchalumov, Y.; Oliver, R.J.; Trenet, W.; Heyer Osorno, R.E.; Sibley, B.D.; Purohit, D.C.; Contet, C.; Roberto, M.; Woodward, J.J.; Mandyam, C.D. Chronic ethanol exposure differentially alters neuronal function in the medial prefrontal cortex and dentate gyrus. Neuropharmacology 2021, 185, 108438. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.S.; Barnet, R.C. Adolescent and adult rats differ in the amnesic effects of acute ethanol in two hippocampus-dependent tasks: Trace and contextual fear conditioning. Behav. Brain Res. 2016, 298 Pt A, 78–87. [Google Scholar] [CrossRef]
- Weitemier, A.Z.; Ryabinin, A.E. Alcohol-induced memory impairment in trace fear conditioning: A hippocampus-specific effect. Hippocampus 2003, 13, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, M.J.; Shin, Y.J.; Lindquist, D.H. Mitigation of postnatal ethanol-induced neuroinflammation ameliorates trace fear memory deficits in juvenile rats. Behav. Brain Res. 2018, 338, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.J. Ethanol disrupts fear conditioning in C57BL/6 mice. J. Psychopharmacol. 2003, 17, 77–81. [Google Scholar] [CrossRef]
- Fannon, M.J.; Mysore, K.K.; Williams, J.; Quach, L.W.; Purohit, D.C.; Sibley, B.D.; Sage-Sepulveda, J.S.; Kharidia, K.M.; Morales Silva, R.J.; Terranova, M.J.; et al. Hippocampal neural progenitor cells play a distinct role in fear memory retrieval in male and female CIE rats. Neuropharmacology 2018, 143, 239–249. [Google Scholar] [CrossRef]
- Bergstrom, H.C.; McDonald, C.G.; Smith, R.F. Alcohol exposure during adolescence impairs auditory fear conditioning in adult Long-Evans rats. Physiol. Behav. 2006, 88, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Staples, M.C.; Herman, M.A.; Lockner, J.W.; Avchalumov, Y.; Kharidia, K.M.; Janda, K.D.; Roberto, M.; Mandyam, C.D. Isoxazole-9 reduces enhanced fear responses and retrieval in ethanol-dependent male rats. J. Neurosci. Res. 2021, 99, 3047–3065. [Google Scholar] [CrossRef] [PubMed]
- O’Dell, L.E.; Roberts, A.J.; Smith, R.T.; Koob, G.F. Enhanced alcohol self-administration after intermittent versus continuous alcohol vapor exposure. Alcohol. Clin. Exp. Res. 2004, 28, 1676–1682. [Google Scholar] [CrossRef] [PubMed]
- Valdez, G.R.; Roberts, A.J.; Chan, K.; Davis, H.; Brennan, M.; Zorrilla, E.P.; Koob, G.F. Increased ethanol self-administration and anxiety-like behavior during acute ethanol withdrawal and protracted abstinence: Regulation by corticotropin-releasing factor. Alcohol. Clin. Exp. Res. 2002, 26, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- George, O.; Hope, B.T. Cortical and amygdalar neuronal ensembles in alcohol seeking, drinking and withdrawal. Neuropharmacology 2017, 122, 107–114. [Google Scholar] [CrossRef]
- Klenowski, P.M. Emerging role for the medial prefrontal cortex in alcohol-seeking behaviors. Addict. Behav. 2018, 77, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D.; Koob, G.F. The addicted brain craves new neurons: Putative role for adult-born progenitors in promoting recovery. Trends Neurosci. 2012, 35, 250–260. [Google Scholar] [CrossRef]
- Mandyam, C.D.; Villalpando, E.G.; Steiner, N.L.; Quach, L.W.; Fannon, M.J.; Somkuwar, S.S. Platelet Endothelial Cell Adhesion Molecule-1 and Oligodendrogenesis: Significance in Alcohol Use Disorders. Brain Sci. 2017, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Abernathy, K.; Chandler, L.J.; Woodward, J.J. Alcohol and the prefrontal cortex. Int. Rev. Neurobiol. 2010, 91, 289–320. [Google Scholar]
- Staples, M.C.; Kim, A.; Mandyam, C.D. Dendritic remodeling of hippocampal neurons is associated with altered NMDA receptor expression in alcohol dependent rats. Mol. Cell Neurosci. 2015, 65, 153–162. [Google Scholar] [CrossRef]
- Staples, M.C.; Mandyam, C.D. Thinking after Drinking: Impaired Hippocampal-Dependent Cognition in Human Alcoholics and Animal Models of Alcohol Dependence. Front. Psychiatry 2016, 7, 162. [Google Scholar] [CrossRef] [PubMed]
- Avchalumov, Y.; Mandyam, C.D. Synaptic Plasticity and its Modulation by Alcohol. Brain Plast. 2020, 6, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Macht, V.A.; Vetreno, R.P.; Crews, F.T. Cholinergic and Neuroimmune Signaling Interact to Impact Adult Hippocampal Neurogenesis and Alcohol Pathology across Development. Front. Pharmacol. 2022, 13, 849997. [Google Scholar] [CrossRef] [PubMed]
- Wooden, J.I.; Thompson, K.R.; Guerin, S.P.; Nawarawong, N.N.; Nixon, K. Consequences of adolescent alcohol use on adult hippocampal neurogenesis and hippocampal integrity. Int. Rev. Neurobiol. 2021, 160, 281–304. [Google Scholar] [PubMed]
- Hansson, A.C.; Nixon, K.; Rimondini, R.; Damadzic, R.; Sommer, W.H.; Eskay, R.; Crews, F.T.; Heilig, M. Long-term suppression of forebrain neurogenesis and loss of neuronal progenitor cells following prolonged alcohol dependence in rats. Int. J. Neuropsychopharmacol. 2010, 13, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Pian, J.P.; Criado, J.R.; Milner, R.; Ehlers, C.L. N-methyl-D-aspartate receptor subunit expression in adult and adolescent brain following chronic ethanol exposure. Neuroscience 2010, 170, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Criado, J.R.; Liu, T.; Ehlers, C.L.; Mathe, A.A. Prolonged chronic ethanol exposure alters neuropeptide Y and corticotropin-releasing factor levels in the brain of adult Wistar rats. Pharmacol. Biochem. Behav. 2011, 99, 104–111. [Google Scholar] [CrossRef]
- Richardson, H.N.; Chan, S.H.; Crawford, E.F.; Lee, Y.K.; Funk, C.K.; Koob, G.F.; Mandyam, C.D. Permanent impairment of birth and survival of cortical and hippocampal proliferating cells following excessive drinking during alcohol dependence. Neurobiol. Dis. 2009, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.E.; Ur, C.L.; Gruol, D.L. Chronic intermittent ethanol exposure enhances NMDA-receptor-mediated synaptic responses and NMDA receptor expression in hippocampal CA1 region. Brain Res. 2005, 1048, 69–79. [Google Scholar] [CrossRef]
- Beroun, A.; Nalberczak-Skora, M.; Harda, Z.; Piechota, M.; Ziolkowska, M.; Caly, A.; Pagano, R.; Radwanska, K. Generation of silent synapses in dentate gyrus correlates with development of alcohol addiction. Neuropsychopharmacology 2018, 43, 1989–1999. [Google Scholar] [CrossRef]
- Roberto, M.; Nelson, T.E.; Ur, C.L.; Gruol, D.L. Long-term potentiation in the rat hippocampus is reversibly depressed by chronic intermittent ethanol exposure. J. Neurophysiol. 2002, 87, 2385–2397. [Google Scholar] [CrossRef]
- Kroener, S.; Mulholland, P.J.; New, N.N.; Gass, J.T.; Becker, H.C.; Chandler, L.J. Chronic alcohol exposure alters behavioral and synaptic plasticity of the rodent prefrontal cortex. PLoS ONE 2012, 7, e37541. [Google Scholar] [CrossRef]
- Varodayan, F.P.; Pahng, A.R.; Davis, T.D.; Gandhi, P.; Bajo, M.; Steinman, M.Q.; Kiosses, W.B.; Blednov, Y.A.; Burkart, M.D.; Edwards, S.; et al. Chronic ethanol induces a pro-inflammatory switch in interleukin-1β regulation of GABAergic signaling in the medial prefrontal cortex of male mice. Brain Behav. Immun. 2023, 110, 125–139. [Google Scholar] [CrossRef]
- Varodayan, F.P.; Sidhu, H.; Kreifeldt, M.; Roberto, M.; Contet, C. Morphological and functional evidence of increased excitatory signaling in the prelimbic cortex during ethanol withdrawal. Neuropharmacology 2018, 133, 470–480. [Google Scholar] [CrossRef]
- Gilpin, N.W.; Richardson, H.N.; Cole, M.; Koob, G.F. Vapor inhalation of alcohol in rats. Curr. Protoc. Neurosci. 2008, 44, 9–29. [Google Scholar] [CrossRef]
- Somkuwar, S.S.; Villalpando, E.G.; Quach, L.W.; Head, B.P.; McKenna, B.S.; Scadeng, M.; Mandyam, C.D. Abstinence from ethanol dependence produces concomitant cortical gray matter abnormalities, microstructural deficits and cognitive dysfunction. Eur. Neuropsychopharmacol. 2021, 42, 22–34. [Google Scholar] [CrossRef]
- Jacobs, N.S.; Cushman, J.D.; Fanselow, M.S. The accurate measurement of fear memory in Pavlovian conditioning: Resolving the baseline issue. J. Neurosci. Methods 2010, 190, 235–239. [Google Scholar] [CrossRef]
- Kim, A.; Zamora-Martinez, E.R.; Edwards, S.; Mandyam, C.D. Structural reorganization of pyramidal neurons in the medial prefrontal cortex of alcohol dependent rats is associated with altered glial plasticity. Brain Struct. Funct. 2015, 220, 1705–1720. [Google Scholar] [CrossRef]
- Thacker, J.S.; Yeung, D.H.; Staines, W.R.; Mielke, J.G. Total protein or high-abundance protein: Which offers the best loading control for Western blotting? Anal. Biochem. 2016, 496, 76–78. [Google Scholar] [CrossRef]
- Welinder, C.; Ekblad, L. Coomassie staining as loading control in Western blot analysis. J. Proteome Res. 2011, 10, 1416–1419. [Google Scholar] [CrossRef]
- Haun, H.L.; Griffin, W.C.; Lopez, M.F.; Solomon, M.G.; Mulholland, P.J.; Woodward, J.J.; McGinty, J.F.; Ron, D.; Becker, H.C. Increasing Brain-Derived Neurotrophic Factor (BDNF) in medial prefrontal cortex selectively reduces excessive drinking in ethanol dependent mice. Neuropharmacology 2018, 140, 35–42. [Google Scholar] [CrossRef]
- McGinty, J.F.; Zelek-Molik, A.; Sun, W.L. Cocaine self-administration causes signaling deficits in corticostriatal circuitry that are reversed by BDNF in early withdrawal. Brain Res. 2015, 1628 Pt A, 82–87. [Google Scholar] [CrossRef]
- Navarro, A.I.; Mandyam, C.D. Protracted abstinence from chronic ethanol exposure alters the structure of neurons and expression of oligodendrocytes and myelin in the medial prefrontal cortex. Neuroscience 2015, 293, 35–44. [Google Scholar] [CrossRef]
- Somkuwar, S.S.; Fannon-Pavlich, M.J.; Ghofranian, A.; Quigley, J.A.; Dutta, R.R.; Galinato, M.H.; Mandyam, C.D. Wheel running reduces ethanol seeking by increasing neuronal activation and reducing oligodendroglial/neuroinflammatory factors in the medial prefrontal cortex. Brain Behav. Immun. 2016, 58, 357–368. [Google Scholar] [CrossRef]
- Nixon, K.; Kim, D.H.; Potts, E.N.; He, J.; Crews, F.T. Distinct cell proliferation events during abstinence after alcohol dependence: Microglia proliferation precedes neurogenesis. Neurobiol. Dis. 2008, 31, 218–229. [Google Scholar] [CrossRef]
- Heilig, M.; Egli, M.; Crabbe, J.C.; Becker, H.C. Acute withdrawal, protracted abstinence and negative affect in alcoholism: Are they linked? Addict. Biol. 2010, 15, 169–184. [Google Scholar] [CrossRef]
- Ron, D.; Wang, J. Frontiers in Neuroscience. The NMDA Receptor and Alcohol Addiction. In Biology of the NMDA Receptor; Van Dongen, A.M., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group, LLC: Boca Raton, FL, USA, 2009. [Google Scholar]
- Hu, X.J.; Ticku, M.K. Chronic ethanol treatment upregulates the NMDA receptor function and binding in mammalian cortical neurons. Brain Res. Mol. Brain Res. 1995, 30, 347–356. [Google Scholar] [CrossRef]
- Hu, X.J.; Follesa, P.; Ticku, M.K. Chronic ethanol treatment produces a selective upregulation of the NMDA receptor subunit gene expression in mammalian cultured cortical neurons. Brain Res. Mol. Brain Res. 1996, 36, 211–218. [Google Scholar] [CrossRef]
- Follesa, P.; Ticku, M.K. Chronic ethanol-mediated up-regulation of the N-methyl-D-aspartate receptor polypeptide subunits in mouse cortical neurons in culture. J. Biol. Chem. 1996, 271, 13297–13299. [Google Scholar] [CrossRef]
- Kalluri, H.S.; Mehta, A.K.; Ticku, M.K. Up-regulation of NMDA receptor subunits in rat brain following chronic ethanol treatment. Brain Res. Mol. Brain Res. 1998, 58, 221–224. [Google Scholar] [CrossRef]
- Cui, Z.; Feng, R.; Jacobs, S.; Duan, Y.; Wang, H.; Cao, X.; Tsien, J.Z. Increased NR2A:NR2B ratio compresses long-term depression range and constrains long-term memory. Sci. Rep. 2013, 3, 1036. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; Liang, J. Role of GABAA receptors in alcohol use disorders suggested by chronic intermittent ethanol (CIE) rodent model. Mol. Brain 2017, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Roberto, M.; Varodayan, F.P. Synaptic targets: Chronic alcohol actions. Neuropharmacology 2017, 122, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Beck, A.; Grüsser, S.M.; Grace, A.A.; Wrase, J. Identifying the neural circuitry of alcohol craving and relapse vulnerability. Addict. Biol. 2009, 14, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Murakoshi, H.; Shin, M.E.; Parra-Bueno, P.; Szatmari, E.M.; Shibata, A.C.E.; Yasuda, R. Kinetics of Endogenous CaMKII Required for Synaptic Plasticity Revealed by Optogenetic Kinase Inhibitor. Neuron 2017, 94, 37–47.e5. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Stoppini, L.; Miyamoto, E.; Muller, D. Long-term potentiation is associated with an increased activity of Ca2+/calmodulin-dependent protein kinase II. J. Biol. Chem. 1993, 268, 7863–7867. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, F.; Goodwani, S.; McCullumsmith, R.E.; Sari, Y. Role of glutamatergic system and mesocorticolimbic circuits in alcohol dependence. Prog. Neurobiol. 2018, 171, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Frankland, P.W.; Bontempi, B. The organization of recent and remote memories. Nat. Rev. Neurosci. 2005, 6, 119–130. [Google Scholar] [CrossRef]
- Doron, A.; Goshen, I. Investigating the transition from recent to remote memory using advanced tools. Brain Res. Bull. 2018, 141, 35–43. [Google Scholar] [CrossRef]
- Woods, N.I.; Kheirbek, M.A. The Small World of a Fear Memory. Neuron 2017, 94, 226–227. [Google Scholar] [CrossRef]
- Beeman, C.L.; Bauer, P.S.; Pierson, J.L.; Quinn, J.J. Hippocampus and medial prefrontal cortex contributions to trace and contextual fear memory expression over time. Learn. Mem. 2013, 20, 336–343. [Google Scholar] [CrossRef] [PubMed]
- McEchron, M.D.; Tseng, W.; Disterhoft, J.F. Neurotoxic lesions of the dorsal hippocampus disrupt auditory-cued trace heart rate (fear) conditioning in rabbits. Hippocampus 2000, 10, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Oommen, S.S.; Morrison, G.E.; Fanselow, M.S. Post-training excitotoxic lesions of the dorsal hippocampus attenuate forward trace, backward trace, and delay fear conditioning in a temporally specific manner. Hippocampus 2002, 12, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Ma, Q.D.; Tinsley, M.R.; Koch, C.; Fanselow, M.S. Inverse temporal contributions of the dorsal hippocampus and medial prefrontal cortex to the expression of long-term fear memories. Learn. Mem. 2008, 15, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Yoon, T.; Otto, T. Differential contributions of dorsal vs. ventral hippocampus to auditory trace fear conditioning. Neurobiol Learn. Mem. 2007, 87, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Pierson, J.L.; Pullins, S.E.; Quinn, J.J. Dorsal hippocampus infusions of CNQX into the dentate gyrus disrupt expression of trace fear conditioning. Hippocampus 2015, 25, 779–785. [Google Scholar] [CrossRef]
- Broadwater, M.; Spear, L.P. Age differences in fear retention and extinction in male Sprague-Dawley rats: Effects of ethanol challenge during conditioning. Behav. Brain Res. 2013, 252, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Fitzgerald, P.J.; MacPherson, K.P.; DeBrouse, L.; Colacicco, G.; Flynn, S.M.; Masneuf, S.; Pleil, K.E.; Li, C.; Marcinkiewcz, C.A.; et al. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-mediated fear extinction encoding. Nat. Neurosci. 2012, 15, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Melia, K.R.; Ryabinin, A.E.; Corodimas, K.P.; Wilson, M.C.; Ledoux, J.E. Hippocampal-dependent learning and experience-dependent activation of the hippocampus are preferentially disrupted by ethanol. Neuroscience 1996, 74, 313–322. [Google Scholar] [CrossRef]
- Czerniawski, J.; Ree, F.; Chia, C.; Otto, T. Dorsal versus ventral hippocampal contributions to trace and contextual conditioning: Differential effects of regionally selective NMDA receptor antagonism on acquisition and expression. Hippocampus 2012, 22, 1528–1539. [Google Scholar] [CrossRef]
- Gilmartin, M.R.; McEchron, M.D. Single neurons in the dentate gyrus and CA1 of the hippocampus exhibit inverse patterns of encoding during trace fear conditioning. Behav. Neurosci. 2005, 119, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, M.G.; Gould, T.J. Effects of drugs of abuse on hippocampal plasticity and hippocampus-dependent learning and memory: Contributions to development and maintenance of addiction. Learn. Mem. 2016, 23, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, J.; Alfonso-Loeches, S.; Guerri, C. Impact of the Innate Immune Response in the Actions of Ethanol on the Central Nervous System. Alcohol. Clin. Exp. Res. 2016, 40, 2260–2270. [Google Scholar] [CrossRef]
- Zorumski, C.F.; Mennerick, S.; Izumi, Y. Acute and chronic effects of ethanol on learning-related synaptic plasticity. Alcohol 2014, 48, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D. The Interplay between the Hippocampus and Amygdala in Regulating Aberrant Hippocampal Neurogenesis during Protracted Abstinence from Alcohol Dependence. Front. Psychiatry 2013, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Nixon, K. Alcohol and adult neurogenesis: Roles in neurodegeneration and recovery in chronic alcoholism. Hippocampus 2006, 16, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Barco, A.; Zablow, L.; Kandel, E.R.; Siegelbaum, S.A.; Zakharenko, S.S. Transient expansion of synaptically connected dendritic spines upon induction of hippocampal long-term potentiation. Proc. Natl. Acad. Sci. USA 2004, 101, 16665–16670. [Google Scholar] [CrossRef]
- George, O.; Sanders, C.; Freiling, J.; Grigoryan, E.; Vu, S.; Allen, C.D.; Crawford, E.; Mandyam, C.D.; Koob, G.F. Recruitment of medial prefrontal cortex neurons during alcohol withdrawal predicts cognitive impairment and excessive alcohol drinking. Proc. Natl. Acad. Sci. USA 2012, 109, 18156–18161. [Google Scholar] [CrossRef] [PubMed]
- Hoover, W.B.; Vertes, R.P. Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct. 2007, 212, 149–179. [Google Scholar] [CrossRef]
- Gabbott, P.; Warner, T.A.; Brown, J.; Salway, P.; Gabbott, T.; Busby, S. Amygdala afferents monosynaptically innervate corticospinal neurons in rat medial prefrontal cortex. J. Comp. Neurol. 2012, 520, 2440–2458. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Knapp, D.J.; Breese, G.R. Accentuated decrease in social interaction in rats subjected to repeated ethanol withdrawals. Alcohol. Clin. Exp. Res. 2002, 26, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.; Lacadie, C.M.; Tuit, K.; Hong, K.I.; Constable, R.T.; Sinha, R. Disrupted ventromedial prefrontal function, alcohol craving, and subsequent relapse risk. JAMA Psychiatry 2013, 70, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Loya, F.; Ma, Q.D.; Fanselow, M.S. Dorsal hippocampus NMDA receptors differentially mediate trace and contextual fear conditioning. Hippocampus 2005, 15, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Jarome, T.J.; Kwapis, J.L.; Werner, C.T.; Parsons, R.G.; Gafford, G.M.; Helmstetter, F.J. The timing of multiple retrieval events can alter GluR1 phosphorylation and the requirement for protein synthesis in fear memory reconsolidation. Learn. Mem. 2012, 19, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Huerta, P.T.; Sun, L.D.; Wilson, M.A.; Tonegawa, S. Formation of temporal memory requires NMDA receptors within CA1 pyramidal neurons. Neuron 2000, 25, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Holehonnur, R.; Phensy, A.J.; Kim, L.J.; Milivojevic, M.; Vuong, D.; Daison, D.K.; Alex, S.; Tiner, M.; Jones, L.E.; Kroener, S.; et al. Increasing the GluN2A/GluN2B Ratio in Neurons of the Mouse Basal and Lateral Amygdala Inhibits the Modification of an Existing Fear Memory Trace. J. Neurosci. 2016, 36, 9490–9504. [Google Scholar] [CrossRef]
- Stiedl, O.; Birkenfeld, K.; Palve, M.; Spiess, J. Impairment of conditioned contextual fear of C57BL/6J mice by intracerebral injections of the NMDA receptor antagonist APV. Behav. Brain Res. 2000, 116, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Monyer, H.; Burnashev, N.; Laurie, D.J.; Sakmann, B.; Seeburg, P.H. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron 1994, 12, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Ladagu, A.D.; Olopade, F.E.; Adejare, A.; Olopade, J.O. GluN2A and GluN2B N-Methyl-D-Aspartate Receptor (NMDARs) Subunits: Their Roles and Therapeutic Antagonists in Neurological Diseases. Pharmaceuticals 2023, 16, 1535. [Google Scholar] [CrossRef]
- Acutain, M.F.; Griebler Luft, J.; Vazquez, C.A.; Popik, B.; Cercato, M.C.; Epstein, A.; Salvetti, A.; Jerusalinsky, D.A.; de Oliveira Alvares, L.; Baez, M.V. Reduced Expression of Hippocampal GluN2A-NMDAR Increases Seizure Susceptibility and Causes Deficits in Contextual Memory. Front. Neurosci. 2021, 15, 644100. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steiner, N.L.; Purohit, D.C.; Tiefenthaler, C.M.; Mandyam, C.D. Abstinence and Fear Experienced during This Period Produce Distinct Cortical and Hippocampal Adaptations in Alcohol-Dependent Rats. Brain Sci. 2024, 14, 431. https://doi.org/10.3390/brainsci14050431
Steiner NL, Purohit DC, Tiefenthaler CM, Mandyam CD. Abstinence and Fear Experienced during This Period Produce Distinct Cortical and Hippocampal Adaptations in Alcohol-Dependent Rats. Brain Sciences. 2024; 14(5):431. https://doi.org/10.3390/brainsci14050431
Chicago/Turabian StyleSteiner, Noah L., Dvijen C. Purohit, Casey M. Tiefenthaler, and Chitra D. Mandyam. 2024. "Abstinence and Fear Experienced during This Period Produce Distinct Cortical and Hippocampal Adaptations in Alcohol-Dependent Rats" Brain Sciences 14, no. 5: 431. https://doi.org/10.3390/brainsci14050431