Auditory Reserve and the Legacy of Auditory Experience

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Legacy of Early Experience: Evidence for the Auditory Reserve
2.1. Evidence from Animal Models
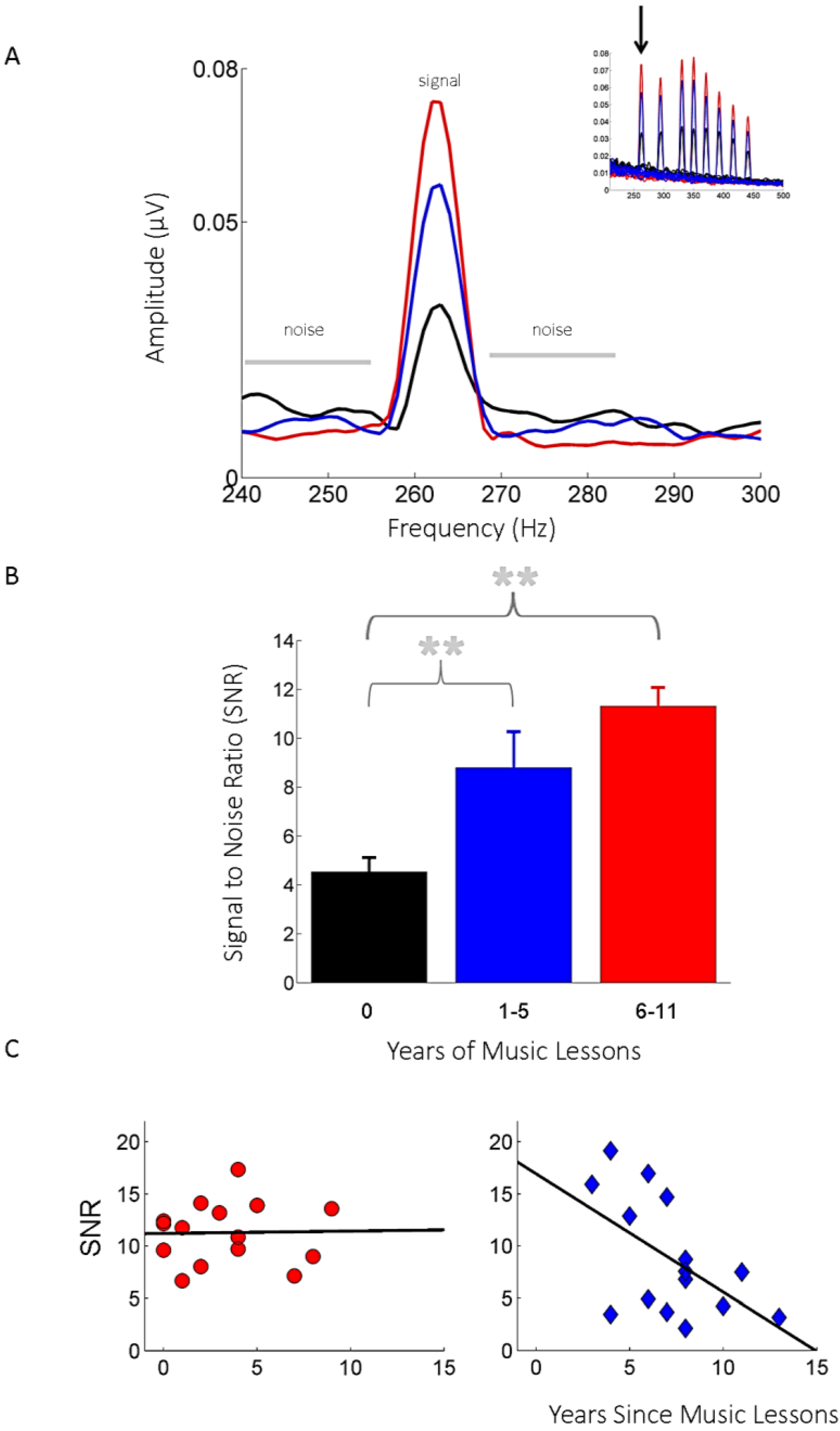
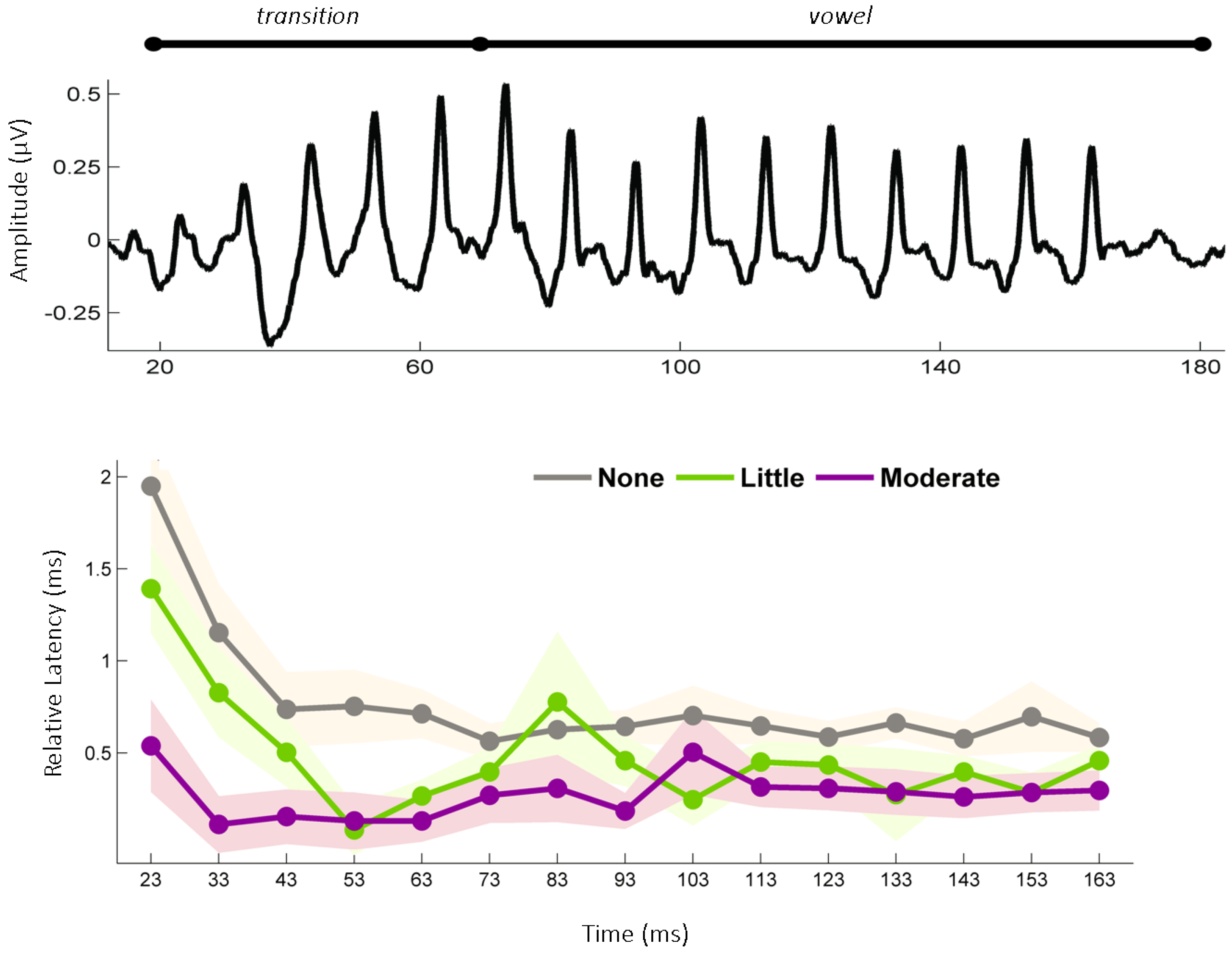
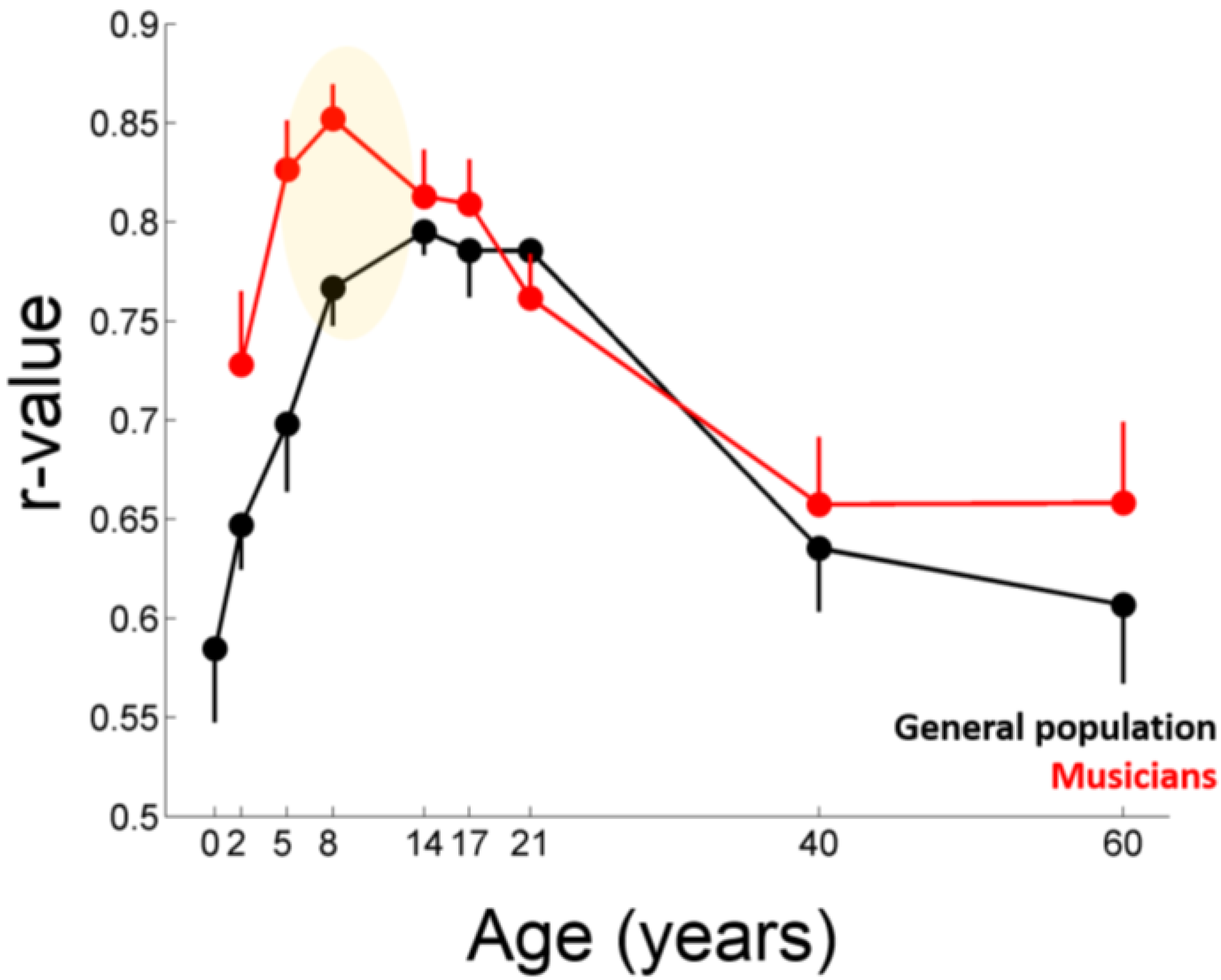
2.2. Evidence from Humans: Musicians
2.2.1. The Effects of Childhood Musical Training on the Mature Auditory System

2.2.2. The Effects of Childhood Musical Training on the Aging Auditory System

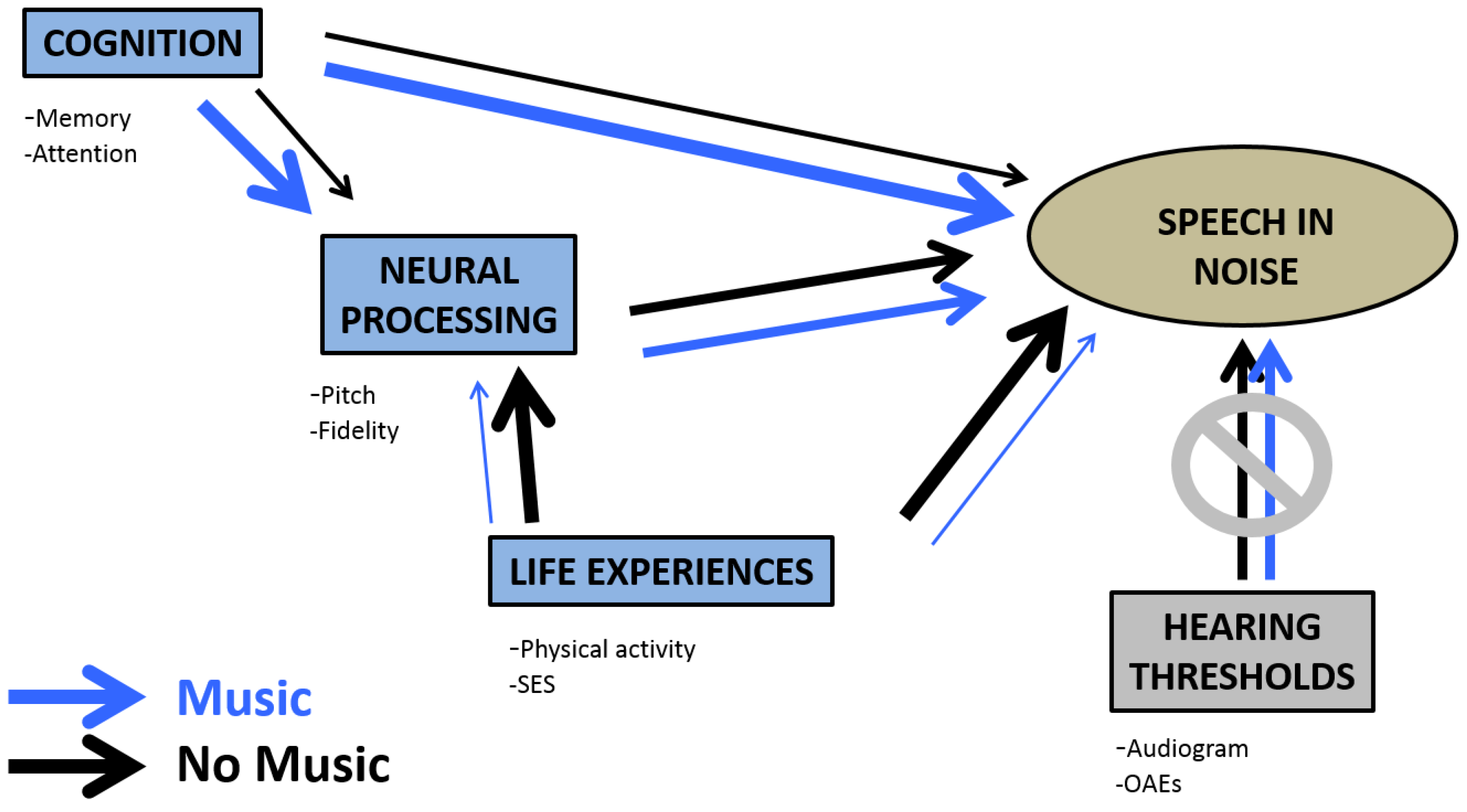
3. Auditory Reserve Defined
4. Exploring the Auditory Reserve
4.1. Auditory Reserve: The Role of Auditory Enrichment and Impoverishment
4.2. Activation of the Auditory Reserve
4.3. Activation of the Auditory Reserve: Evidence from Animal Models
4.4. Specificity of the Auditory Reserve: The Conditions that Allow an Earlier Experience to Affect Later Function
4.5. The Auditory Reserve and Sensitive Periods

4.6. Establishing the Auditory Reserve: The Role of in Utero Experiences
4.7. Interaction between the Auditory Reserve and Cognitive Reserve

5. Conclusions
Author Contributions
Acknowledgements
Conflicts of Interest
References
- Stern, Y. Cognitive reserve. Neuropsychologia 2009, 47, 2015–2028. [Google Scholar] [CrossRef]
- Steffener, J.; Stern, Y. Exploring the neural basis of cognitive reserve in aging. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 467–473. [Google Scholar] [CrossRef]
- Barulli, D.; Stern, Y. Efficiency, capacity, compensation, maintenance, plasticity: Emerging concepts in cognitive reserve. Trends Cogn. Sci. 2013, 17, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Steffener, J.; Barulli, D.; Habeck, C.; O’Shea, D.; Razlighi, Q.; Stern, Y. The role of education and verbal abilities in altering the effect of age-related gray matter differences on cognition. PLoS One 2014, 9, e91196. [Google Scholar]
- Bialystok, E.; Craik, F.I.M.; Freedman, M. Bilingualism as a protection against the onset of symptoms of dementia. Neuropsychologia 2007, 45, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.; Sacker, A. Lifetime antecedents of cognitive reserve. J. Clin. Exp. Neuropsychol. 2003, 25, 614–624. [Google Scholar] [CrossRef]
- Zhang, L.I.; Bao, S.; Merzenich, M.M. Persistent and specific influences of early acoustic environments on primary auditory cortex. Nat. Neurosci. 2001, 4, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.I.; Bao, S.; Merzenich, M.M. Disruption of primary auditory cortex by synchronous auditory inputs during a critical period. Proc. Natl. Acad. Sci. USA 2002, 99, 2309–2314. [Google Scholar] [CrossRef]
- Sanes, D.H.; Constantine-Paton, M. The sharpening of frequency tuning curves requires patterned activity during development in the mouse, Mus musculus. J. Neurosci. 1985, 5, 1152–1166. [Google Scholar] [PubMed]
- Oliver, D.L.; Izquierdo, M.A.; Malmierca, M.S. Persistent effects of early augmented acoustic environment on the auditory brainstem. Neuroscience 2011, 184, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Batkin, S.; Groth, H.; Watson, J.R.; Ansberry, M. Effects of auditory deprivation on the development of auditory sensitivity in albino rats. Electroencephalogr. Clin. Neurophysiol. 1970, 28, 351–359. [Google Scholar] [CrossRef]
- Kral, A.; Eggermont, J.J. What’s to lose and what’s to learn: Development under auditory deprivation, cochlear implants and limits of cortical plasticity. Brain Res. Rev. 2007, 56, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Sarro, E.C.; Sanes, D.H. The cost and benefit of juvenile training on adult perceptual skill. J. Neurosci. 2011, 31, 5383–5391. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Sarro, E.C.; Sanes, D.H. Auditory training during development mitigates a hearing loss-induced perceptual deficit. Front. Syst. Neurosci. 2014, 8. [Google Scholar] [CrossRef]
- Strait, D.L.; Kraus, N. Biological impact of auditory expertise across the life span: Musicians as a model of auditory learning. Hear. Res. 2013, 308, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Bidelman, G.M.; Krishnan, A. Effects of reverberation on brainstem representation of speech in musicians and non-musicians. Brain Res. 2010, 1355, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Bidelman, G.M.; Gandour, J.T.; Krishnan, A. Musicians demonstrate experience-dependent brainstem enhancement of musical scale features within continuously gliding pitch. Neurosci. Lett. 2011, 503, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Panda, M.R. Experience-dependent learning of auditory temporal resolution: Evidence from Carnatic-trained musicians. Neuroreport 2014, 25, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Kraus, N.; Chandrasekaran, B. Music training for the development of auditory skills. Nat. Rev. Neurosci. 2010, 11, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D. Why would Musical Training Benefit the Neural Encoding of Speech? The OPERA Hypothesis. Front. Psychol. 2011, 2, 142. [Google Scholar] [CrossRef] [PubMed]
- Zendel, B.R.; Alain, C. Musicians experience less age-related decline in central auditory processing. Psychol. Aging 2012, 27, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Skoe, E.; Kraus, N. Musical experience limits the degradative effects of background noise on the neural processing of sound. J. Neurosci. 2009, 29, 14100–14107. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Acceleration of age-related hearing loss by early noise exposure: Evidence of a misspent youth. J. Neurosci. 2006, 26, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Kraus, N.; Nicol, T. The Cognitive Auditory System: The Role of Learning in Shaping the Biology of the Auditory System. In Perspectives on Auditory Research; Popper, A.N., Fay, R.R., Eds.; Springer: New York, NY, USA, 2014; pp. 299–319. [Google Scholar]
- Samelli, A.G.; Matas, C.G.; Carvallo, R.M.; Gomes, R.F.; de Beija, C.S.; Magliaro, F.C.; Rabelo, C.M. Audiological and electrophysiological assessment of professional pop/rock musicians. Noise Health 2012, 14, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Strait, D.L.; O’Connell, A.S.; Kraus, N. Biological impact of preschool music classes on processing speech in noise. Dev. Cogn. Neurosci. 2013, 6C, 51–60. [Google Scholar] [CrossRef]
- Tierney, A.; Krizman, J.; Skoe, E.; Johnston, K.; Kraus, N. High school music classes enhance the neural processing of speech. Front. Educ. Psychol. 2013, 4, 855. [Google Scholar] [CrossRef]
- Moreno, S.; Bidelman, G.M. Examining neural plasticity and cognitive benefit through the unique lens of musical training. Hear. Res. 2014, 308, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.; Kraus, N. Hearing matters: Music training: An antidote for aging. Hear. J. 2013, 66, 52. [Google Scholar] [CrossRef]
- Parbery-Clark, A.; Anderson, S.; Hittner, E.; Kraus, N. Musical experience offsets age-related delays in neural timing. Neurobiol. Aging 2012, 33, 1483e1–1483e4. [Google Scholar] [CrossRef]
- Hanna-Pladdy, B.; MacKay, A. The relation between instrumental musical activity and cognitive aging. Neuropsychology 2011, 25, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Skoe, E.; Kraus, N. A little goes a long way: How the adult brain is shaped by musical training in childhood. J. Neurosci. 2012, 32, 11507–11510. [Google Scholar] [CrossRef] [PubMed]
- White-Schwoch, T.; Carr, K.W.; Anderson, S.; Strait, D.L.; Kraus, N. Older adults benefit from music training early in life: Biological evidence for long-term training-driven plasticity. J. Neurosci. 2013, 33, 17667–17674. [Google Scholar] [CrossRef] [PubMed]
- Krizman, J.; Skoe, E.; Marian, V.; Kraus, N. Bilingualism increases neural response consistency and attentional control: Evidence for sensory and cognitive coupling. Brain Lang. 2014, 128, 34–40. [Google Scholar] [CrossRef] [PubMed]
- McMahon, E.; Wintermark, P.; Lahav, A. Auditory brain development in premature infants: The importance of early experience. Ann. N. Y. Acad. Sci. 2012, 1252, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Chartrand, J.P.; Peretz, I.; Belin, P. Auditory recognition expertise and domain specificity. Brain Res. 2008, 1220, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Krizman, J.; Marian, V.; Shook, A.; Skoe, E.; Kraus, N. Subcortical encoding of sound is enhanced in bilinguals and relates to executive function advantages. Proc. Natl. Acad. Sci. USA 2012, 109, 7877–7881. [Google Scholar] [CrossRef] [PubMed]
- Fuller, C.D.; Galvin, J.J., 3rd; Maat, B.; Free, R.H.; Baskent, D. The musician effect: Does it persist under degraded pitch conditions of cochlear implant simulations? Front. Neurosci. 2014, 8, 179. [Google Scholar] [CrossRef]
- Sorqvist, P.; Hurtig, A.; Ljung, R.; Ronnberg, J. High second-language proficiency protects against the effects of reverberation on listening comprehension. Scand. J. Psychol. 2014, 55, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Anderson, S.; Kraus, N. Musicians change their tune: How hearing loss alters the neural code. Hear. Res. 2013, 302, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, F.; Hu, H.; Sun, X.; Kilgard, M.P.; Merzenich, M.M.; Zhou, X. Environmental acoustic enrichment promotes recovery from developmentally degraded auditory cortical processing. J. Neurosci. 2014, 34, 5406–5415. [Google Scholar] [CrossRef] [PubMed]
- Gravel, J.S.; Wallace, I.F.; Ruben, R.J. Auditory Consequences of Early Mild Hearing Loss Associated with Otitis Media. Acta Otolaryngol. 1996, 116, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Gourevitch, B.; Edeline, J.M.; Occelli, F.; Eggermont, J.J. Is the din really harmless? Long-term effects of non-traumatic noise on the adult auditory system. Nat. Rev. Neurosci. 2014, 15, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Skoe, E.; Krizman, J.; Kraus, N. The impoverished brain: Disparities in maternal education affect the neural response to sound. J. Neurosci. 2013, 33, 17221–17231. [Google Scholar] [CrossRef] [PubMed]
- Conway, C.M.; Pisoni, D.B.; Kronenberger, W.G. The importance of sound for cognitive sequencing abilities: The auditory scaffolding hypothesis. Curr. Dir. Psychol. Sci. 2009, 18, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.T.; McPherson, G.E.; Peretz, I.; Berkovic, S.F.; Wilson, S.J. The genetic basis of music ability. Front. Psychol. 2014, 5, 658. [Google Scholar] [CrossRef] [PubMed]
- Dohn, A.; Garza-Villarreal, E.A.; Chakravarty, M.M.; Hansen, M.; Lerch, J.P.; Vuust, P. Gray- and White-Matter Anatomy of Absolute Pitch Possessors. Cereb. Cortex 2013. [Google Scholar] [CrossRef]
- Gervain, J.; Vines, B.W.; Chen, L.M.; Seo, R.J.; Hensch, T.K.; Werker, J.F.; Young, A.H. Valproate reopens critical-period learning of absolute pitch. Front. Syst. Neurosci. 2013, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Rogenmoser, L.; Elmer, S.; Jancke, L. Absolute Pitch: Evidence for Early Cognitive Facilitation during Passive Listening as Revealed by Reduced P3a Amplitudes. J. Cogn. Neurosci. 2014. [Google Scholar] [CrossRef]
- Saffran, J.R.; Griepentrog, G.J. Absolute pitch in infant auditory learning: Evidence for developmental reorganization. Dev. Psychol. 2001, 37, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.J. Absolute pitch: A model for understanding the influence of genes and development on neural and cognitive function. Nat. Neurosci. 2003, 6, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Skoe, E.; Banai, K.; Kraus, N. Perception of speech in noise: Neural correlates. J. Cogn. Neurosci. 2011, 23, 2268–2279. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; McIntosh, A.R.; Tulving, E.; Nyberg, L.; Grady, C.L. Age-related differences in effective neural connectivity during encoding and recall. Neuroreport 1997, 8, 3479–3483. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.C.; Ettlinger, M.; Sheppard, J.P.; Gunasekera, G.M.; Dhar, S. Neuroanatomical characteristics and speech perception in noise in older adults. Ear Hear. 2010, 31, 471–479. [Google Scholar] [CrossRef]
- Stern, Y.; Habeck, C.; Moeller, J.; Scarmeas, N.; Anderson, K.E.; Hilton, H.J.; Flynn, J.; Sackeim, H.; van Heertum, R. Brain Networks Associated with Cognitive Reserve in Healthy Young and Old Adults. Cereb. Cortex 2005, 15, 394–402. [Google Scholar] [PubMed]
- Warraich, Z.; Kleim, J.A. Neural plasticity: The biological substrate for neurorehabilitation. PM R. 2010, 2, S208–S219. [Google Scholar] [CrossRef]
- Knudsen, E.I. Capacity for plasticity in the adult owl auditory system expanded by juvenile experience. Science 1998, 279, 1531–1533. [Google Scholar] [CrossRef]
- Linkenhoker, B.A.; von der Ohe, C.G.; Knudsen, E.I. Anatomical traces of juvenile learning in the auditory system of adult barn owls. Nat. Neurosci. 2005, 8, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Musacchia, G.; Sams, M.; Skoe, E.; Kraus, N. Musicians have enhanced subcortical auditory and audiovisual processing of speech and music. Proc. Natl. Acad. Sci. USA 2007, 104, 15894–15898. [Google Scholar] [CrossRef] [PubMed]
- Strait, D.L.; Chan, K.; Ashley, R.; Kraus, N. Specialization among the specialized: Auditory brainstem function is tuned in to timbre. Cortex 2012, 48, 360–362. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Skoe, E.; Lam, C.; Kraus, N. Musician Enhancement for Speech-in-Noise. Ear Hear. 2009, 30, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.C.; Skoe, E.; Russo, N.M.; Dees, T.; Kraus, N. Musical experience shapes human brainstem encoding of linguistic pitch patterns. Nat. Neurosci. 2007, 10, 420–422. [Google Scholar] [PubMed]
- Strait, D.L.; Kraus, N.; Skoe, E.; Ashley, R. Musical experience and neural efficiency: Effects of training on subcortical processing of vocal expressions of emotion. Eur. J. Neurosci. 2009, 29, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Skoe, E.; Kraus, N. Human Subcortical Auditory Function Provides a New Conceptual Framework for Considering Modularity: A Commentary of Isabelle Peretz’ Keynote Paper. In Language and Music as Cognitive Systems; Rebuschat, P., Ed.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Long, M.H. Maturational Constraints on Language Development. Stud. Second Lang. Acquis. 1990, 12, 251–285. [Google Scholar] [CrossRef]
- Seliger, H.W. Implications of a Multiple Critical Periods Hypothesis for Second Language Learning; Second language acquisition research: Issues and implications; Ritchie, W., Ed.; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Snyder, T.D.; Dillow, S.A. Elementary and Secondary Education. In Digest of Education Statistics, 2011 (NCES 2012–001); Institute of Education Sciences: Washington, DC, USA, 2012. [Google Scholar]
- Penhune, V.B. Sensitive periods in human development: Evidence from musical training. Cortex 2011, 47, 1126–1137. [Google Scholar] [CrossRef]
- Flege, J.E.; Yeni-Komshian, G.H.; Liu, S. Age Constraints on Second-Language Acquisition. J. Mem. Lang. 1999, 41, 78–104. [Google Scholar] [CrossRef]
- Kral, A.; Sharma, A. Developmental neuroplasticity after cochlear implantation. Trends Neurosci. 2012, 35, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Dorman, M.F.; Kral, A. The influence of a sensitive period on central auditory development in children with unilateral and bilateral cochlear implants. Hear. Res. 2005, 203, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Tillein, J.; Heid, S.; Lang, E.; Hartmann, R.; Kral, A. Development of brainstem-evoked responses in congenital auditory deprivation. Neural. Plast. 2012, 2012. [Google Scholar] [CrossRef]
- Skoe, E.; Krizman, J.; Anderson, S.; Kraus, N. Stability and Plasticity of Auditory Brainstem Function Across the Lifespan. Cereb. Cortex 2013. [Google Scholar] [CrossRef]
- Skoe, E.; Kraus, N. Musical training heightens auditory brainstem function during sensitive periods in development. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, E.; White-Schwoch, T.; Woodruff Carr, K.; Skoe, E.; Kraus, N. Continued maturation of the click-evoked auditory brainstem response in preschoolers. J. Am. Acad. Audiol. 2014, in press. [Google Scholar]
- Takesian, A.E.; Kotak, V.C.; Sanes, D.H. Age-dependent effect of hearing loss on cortical inhibitory synapse function. J. Neurophysiol. 2012, 107, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Brainard, M.S.; Knudsen, E.I. Sensitive periods for visual calibration of the auditory space map in the barn owl optic tectum. J. Neurosci. 1998, 18, 3929–3942. [Google Scholar] [PubMed]
- Zhou, X.; Panizzutti, R.; de Villers-Sidani, E.; Madeira, C.; Merzenich, M.M. Natural restoration of critical period plasticity in the juvenile and adult primary auditory cortex. J. Neurosci. 2011, 31, 5625–5634. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Yang, C.H.; Hsu, K.S.; Ming, G.L.; Song, H. A critical period for enhanced synaptic plasticity in newly generated neurons of the adult brain. Neuron 2007, 54, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Birnholz, J.C.; Benacerraf, B.R. The development of human fetal hearing. Science 1983, 222, 516–518. [Google Scholar] [CrossRef]
- Hepper, P.G.; Shahidullah, B.S. Development of fetal hearing. Arch. Dis. Child. 1994, 71, F81–F87. [Google Scholar] [CrossRef] [PubMed]
- Partanen, E.; Kujala, T.; Tervaniemi, M.; Huotilainen, M. Prenatal music exposure induces long-term neural effects. PLoS One 2013, 8, e78946. [Google Scholar]
- Fava, E.; Hull, R.; Bortfeld, H. Linking behavioral and neurophysiological indicators of perceptual tuning to language. Front. Psychol. 2011, 2, 174. [Google Scholar] [CrossRef] [PubMed]
- Ullal-Gupta, S.; der Nederlanden, C.M.V.B.; Tichko, P.; Lahav, A.; Hannon, E.E. Linking prenatal experience to the emerging musical mind. Front. Syst. Neurosci. 2013, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Trammer, R.M.; Aust, G.; Koster, K.; Obladen, M. Narcotic and nicotine effects on the neonatal auditory system. Acta Paediatr. 1992, 81, 962–965. [Google Scholar] [CrossRef] [PubMed]
- McFadden, D. Masculinization effects in the auditory system. Arch. Sex. Behav. 2002, 31, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Weikum, W.M.; Oberlander, T.F.; Hensch, T.K.; Werker, J.F. Prenatal exposure to antidepressants and depressed maternal mood alter trajectory of infant speech perception. Proc. Natl. Acad. Sci. USA 2012, 109, 17221–17227. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, M.; Gonzalez-Frankenberger, B.; Tremblay, J.; Vannasing, P.; Martinez-Montes, E.; Belin, P.; Beland, R.; Francoeur, D.; Carceller, A.M.; Wallois, F.; Lassonde, M. Mother and stranger: an electrophysiological study of voice processing in newborns. Cereb. Cortex 2011, 21, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.R.; Yaffe, K.; Xia, J.; Xue, Q.L.; Harris, T.B.; Purchase-Helzner, E.; Satterfield, S.; Ayonayon, H.N.; Ferrucci, L.; Simonsick, E.M. Hearing loss and cognitive decline in older adults. JAMA Intern. Med. 2013, 173, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Conway, C.M.; Pisoni, D.B.; Anaya, E.M.; Karpicke, J.; Henning, S.C. Implicit sequence learning in deaf children with cochlear implants. Dev. Sci. 2011, 14, 69–82. [Google Scholar] [CrossRef]
- Rocha-Muniz, C.N.; Zachi, E.C.; Teixeira, R.A.; Ventura, D.F.; Befi-Lopes, D.M.; Schochat, E. Association between language development and auditory processing disorders. Braz. J. Otorhinolaryngol. 2014, 80, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.H.; Granot, R.Y.; Ahissar, M. The enigma of dyslexic musicians. Neuropsychologia 2014, 54, 28–40. [Google Scholar]
- Anderson, S.; White-Schwoch, T.; Parbery-Clark, A.; Kraus, N. A dynamic auditory-cognitive system supports speech-in-noise perception in older adults. Hear. Res. 2013, 300, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Bialystok, E. Reshaping the mind: The benefits of bilingualism. Can. J. Exp. Psychol. 2011, 65, 229–235. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoe, E.; Kraus, N. Auditory Reserve and the Legacy of Auditory Experience. Brain Sci. 2014, 4, 575-593. https://doi.org/10.3390/brainsci4040575
Skoe E, Kraus N. Auditory Reserve and the Legacy of Auditory Experience. Brain Sciences. 2014; 4(4):575-593. https://doi.org/10.3390/brainsci4040575
Chicago/Turabian StyleSkoe, Erika, and Nina Kraus. 2014. "Auditory Reserve and the Legacy of Auditory Experience" Brain Sciences 4, no. 4: 575-593. https://doi.org/10.3390/brainsci4040575
APA StyleSkoe, E., & Kraus, N. (2014). Auditory Reserve and the Legacy of Auditory Experience. Brain Sciences, 4(4), 575-593. https://doi.org/10.3390/brainsci4040575