
Early Life Stress, Nicotinic Acetylcholine Receptors and Alcohol Use Disorders

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Alcohol Use Disorders: What’s All the Stress About?
2. The Two-Way Interplay Between Stress and Alcohol Controls Alcohol Consumption
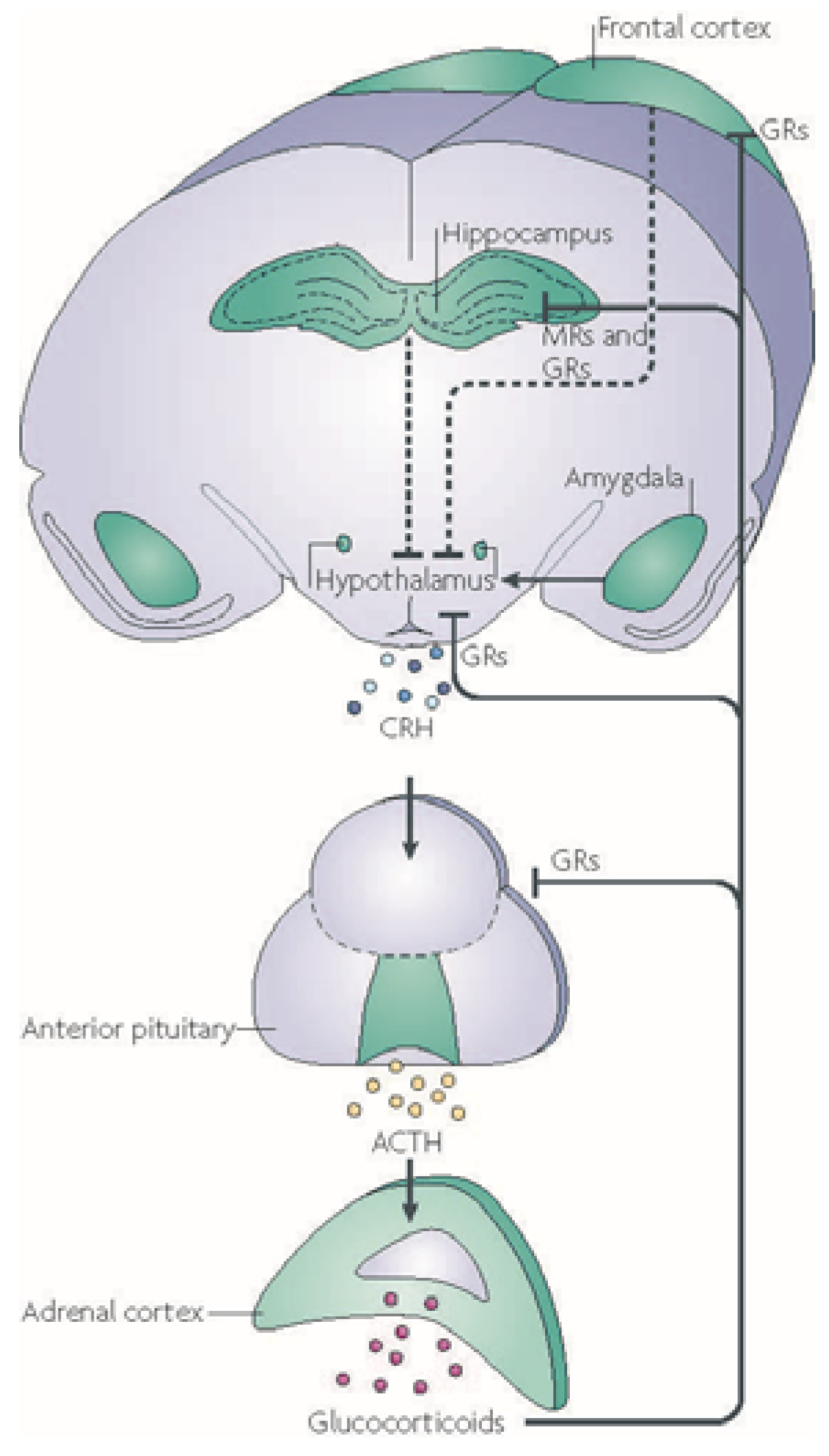
3. Stress Changes How the Brain Functions

4. AUDs and the Nucleus Accumbens
5. Early Life Stress Causes Neuroplastic Changes in the Nucleus Accumbens
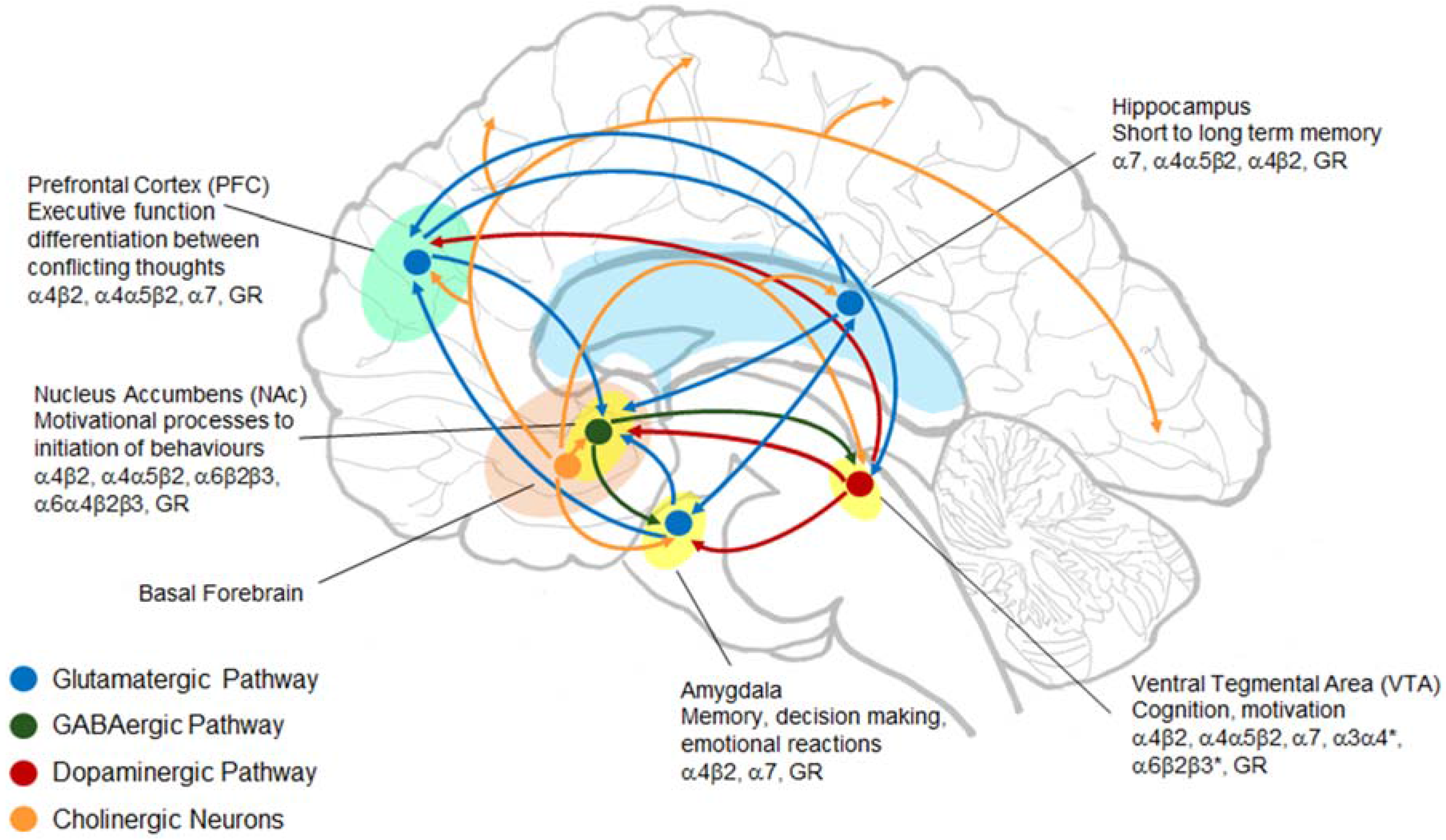
6. Nucleus Accumbens Regulates Cholinergic Output to PFC
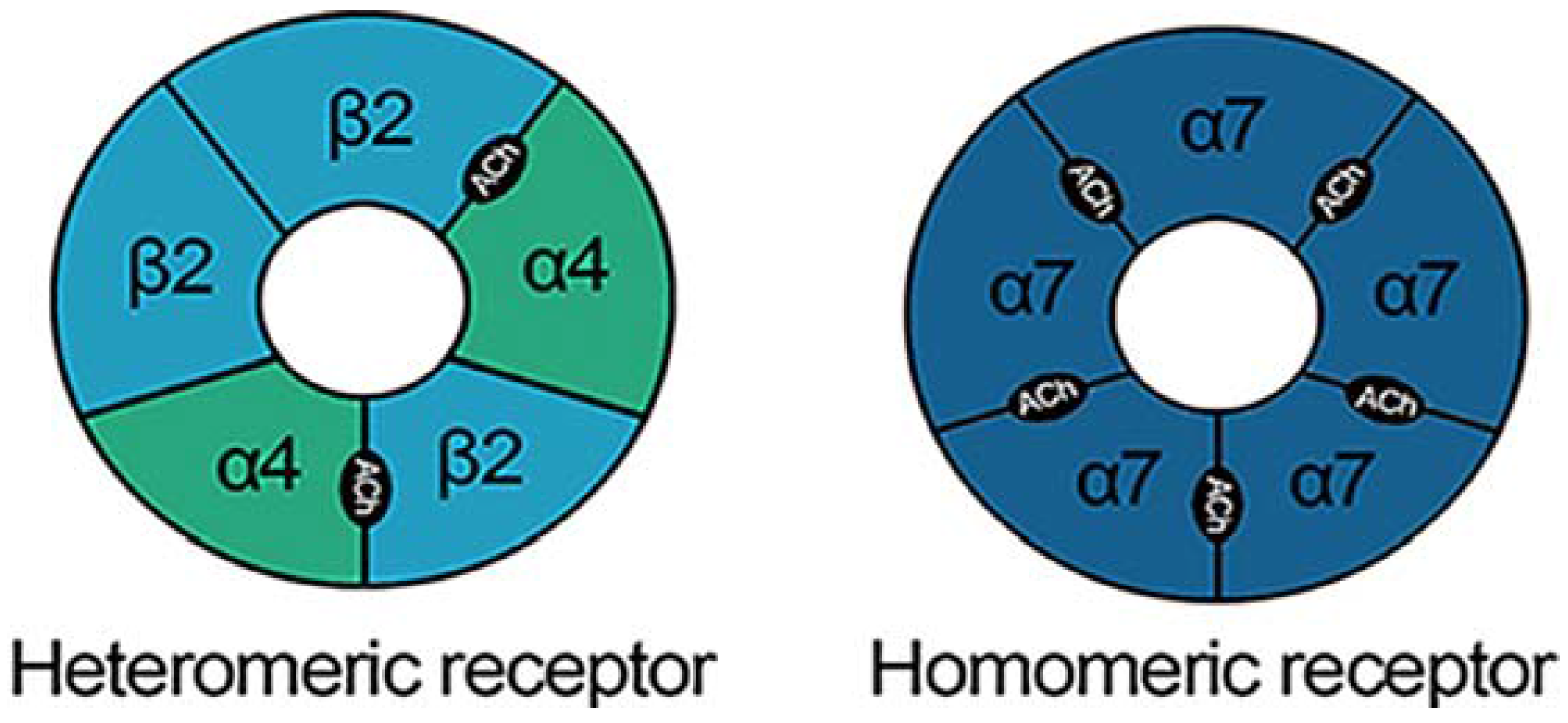
7. What are nAChRs and How are They Involved in AUDs?


8. Nicotinic Acetylcholine Receptors are Modulated by Alcohol and Stress
9. nAChR Subtypes and Their Role in Stress and AUDs
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Srinivasan, S.; Shariff, M.; Bartlett, S.E. The role of the glucocorticoids in developing resilience to stress and addiction. Front. Psychiatry 2013, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Jastreboff, A.M. Stress as a common risk factor for obesity and addiction. Biol. Psychiatry 2013, 73, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Haass-Koffler, C.L.; Bartlett, S.E. Stress and addiction: Contribution of the corticotropin releasing factor (crf) system in neuroplasticity. Front. Mol. Neurosci. 2012, 5, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, S.J.; Murrough, J.W.; Han, M.H.; Charney, D.S.; Nestler, E.J. Neurobiology of resilience. Nat. Neurosci. 2012, 15, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Enoch, M.A. The role of early life stress as a predictor for alcohol and drug dependence. Psychopharmacology 2011, 214, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Enoch, M.A. The influence of gene-environment interactions on the development of alcoholism and drug dependence. Curr. Psychiatry Rep. 2012, 14, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.T.; Back, S.E. Childhood trauma, posttraumatic stress disorder, and alcohol dependence. Alcohol Res. Curr. Rev. 2012, 34, 408–413. [Google Scholar]
- Daoura, L.; Haaker, J.; Nylander, I. Early environmental factors differentially affect voluntary ethanol consumption in adolescent and adult male rats. Alcohol. Clin. Exp. Res. 2011, 35, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Anisman, H.; Zaharia, M.D.; Meaney, M.J.; Merali, Z. Do early-life events permanently alter behavioral and hormonal responses to stressors? Int. J. Dev. Neurosci. 1998, 16, 149–164. [Google Scholar] [CrossRef]
- Levine, S. Developmental determinants of sensitivity and resistance to stress. Psychoneuroendocrinology 2005, 30, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Chassin, L.; Pillow, D.R.; Curran, P.J.; Molina, B.S.; Barrera, M., Jr. Relation of parental alcoholism to early adolescent substance use: A test of three mediating mechanisms. J. Abnorm. Psychol. 1993, 102, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Contoreggi, C.; Lee, M.R.; Chrousos, G. Addiction and corticotropin-releasing hormone type 1 receptor antagonist medications. Ann. NY Acad. Sci. 2013, 1282, 107–118. [Google Scholar] [CrossRef] [PubMed]
- See, R.E.; Waters, R.P. Pharmacologically-induced stress: A cross-species probe for translational research in drug addiction and relapse. Am. J. Transl. Res. 2010, 3, 81–89. [Google Scholar] [PubMed]
- Shalev, U.; Erb, S.; Shaham, Y. Role of crf and other neuropeptides in stress-induced reinstatement of drug seeking. Brain Res. 2010, 1314, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Simms, J.A.; Nielsen, C.K.; Li, R.; Bartlett, S.E. Intermittent access ethanol consumption dysregulates crf function in the hypothalamus and is attenuated by the crf-r1 antagonist, cp-376395. Addict. Biol. 2014, 19, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Spanagel, R.; Noori, H.R.; Heilig, M. Stress and alcohol interactions: Animal studies and clinical significance. Trends Neurosci. 2014, 37, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Breese, G.R.; Sinha, R.; Heilig, M. Chronic alcohol neuroadaptation and stress contribute to susceptibility for alcohol craving and relapse. Pharmacol. Ther. 2011, 129, 149–171. [Google Scholar] [CrossRef] [PubMed]
- Haass-Koffler, C.L.; Leggio, L.; Kenna, G.A. Pharmacological approaches to reducing craving in patients with alcohol use disorders. CNS Drugs 2014, 28, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Adinoff, B.; Iranmanesh, A.; Veldhuis, J.; Fisher, L. Disturbances of the stress response: The role of the hpa axis during alcohol withdrawal and abstinence. Alcohol Health Res. World 1998, 22, 67–72. [Google Scholar] [PubMed]
- Bhutada, P.; Mundhada, Y.; Ghodki, Y.; Dixit, P.; Umathe, S.; Jain, K. Acquisition, expression, and reinstatement of ethanol-induced conditioned place preference in mice: Effects of exposure to stress and modulation by mecamylamine. J. Psychopharmacol. (Oxf., Engl.) 2012, 26, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Groeneweg, F.L.; Karst, H.; de Kloet, E.R.; Joels, M. Rapid non-genomic effects of corticosteroids and their role in the central stress response. J. Endocrinol. 2011, 209, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Arias, A.J.; Sewell, R.A. Pharmacogenetically driven treatments for alcoholism. CNS Drugs 2012, 26, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Heilig, M.; Goldman, D.; Berrettini, W.; O’Brien, C.P. Pharmacogenetic approaches to the treatment of alcohol addiction. Nat. Rev. Neurosci. 2011, 12, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Vengeliene, V.; Bilbao, A.; Molander, A.; Spanagel, R. Neuropharmacology of alcohol addiction. Br. J. Pharmacol. 2008, 154, 299–315. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Allostasis and allostatic load: Implications for neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- McEwen, B.S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R. Hormones and the stressed brain. Ann. NY Acad. Sci. 2004, 1018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.L.; Russell, M.; Skinner, J.B.; Frone, M.R.; Mudar, P. Stress and alcohol use: Moderating effects of gender, coping, and alcohol expectancies. J. Abnorm. Psychol. 1992, 101, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.M.; Hersen, M.; Eisler, R.M.; Hilsman, G. Effects of social stress on operant drinking of alcoholics and social drinkers. Behav. Res. Ther. 1974, 12, 67–72. [Google Scholar] [CrossRef]
- Adriana, D.-A.; Alejandro, D.-M.; Leonila Rosa, D.-M. The complex interplay of genetics, epigenetics, and environment in the predisposition to alcohol dependence. Salud Ment. 2011, 34, 157–166. [Google Scholar]
- Zannas, A.S.; West, A.E. Epigenetics and the regulation of stress vulnerability and resilience. Neuroscience 2014, 264, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Roman, E.; Nylander, I. The impact of emotional stress early in life on adult voluntary ethanol intake-results of maternal separation in rats. Stress (Amst., Neth.) 2005, 8, 157–174. [Google Scholar] [CrossRef]
- Brenhouse, H.C.; Lukkes, J.L.; Andersen, S.L. Early life adversity alters the developmental profiles of addiction-related prefrontal cortex circuitry. Brain Sci. 2013, 3, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Choy, K.H.; van den Buuse, M. Attenuated disruption of prepulse inhibition by dopaminergic stimulation after maternal deprivation and adolescent corticosterone treatment in rats. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2008, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Goff, B.; Gee, D.G.; Telzer, E.H.; Humphreys, K.L.; Gabard-Durnam, L.; Flannery, J.; Tottenham, N. Reduced nucleus accumbens reactivity and adolescent depression following early-life stress. Neuroscience 2013, 249, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.T.; Langenecker, S.A.; Kennedy, S.E.; Zubieta, J.K.; Heitzeg, M.M. Fmri bold responses to negative stimuli in the prefrontal cortex are dependent on levels of recent negative life stress in major depressive disorder. Psychiatry Res. 2010, 183, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, A.N.; Locke, J.L.; McCool, B.A.; Weiner, J.L.; Jones, S.R. Social isolation rearing increases nucleus accumbens dopamine and norepinephrine responses to acute ethanol in adulthood. Alcohol. Clin. Exp. Res. 2014, 38, 2770–2779. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.R.; Staudinger, K.; Scheck, L.; Olive, M.F. The effects of maternal separation on adult methamphetamine self-administration, extinction, reinstatement, and mecp2 immunoreactivity in the nucleus accumbens. Front. Psychiatry 2013, 4, 55. [Google Scholar] [CrossRef] [PubMed]
- Pena, C.J.; Neugut, Y.D.; Calarco, C.A.; Champagne, F.A. Effects of maternal care on the development of midbrain dopamine pathways and reward-directed behavior in female offspring. Eur. J. Neurosci. 2014, 39, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Post, R.M. Mechanisms of illness progression in the recurrent affective disorders. Neurotox. Res. 2010, 18, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Silveira, P.P.; Portella, A.K.; Assis, S.A.; Nieto, F.B.; Diehl, L.A.; Crema, L.M.; Peres, W.; Costa, G.; Scorza, C.; Quillfeldt, J.A.; et al. Early life experience alters behavioral responses to sweet food and accumbal dopamine metabolism. Int. J. Dev. Neurosci. 2010, 28, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Fentress, H.M.; Hoversten, M.T.; Zhang, L.; Hebda-Bauer, E.K.; Watson, S.J.; Seasholtz, A.F.; Akil, H. Early-life forebrain glucocorticoid receptor overexpression increases anxiety behavior and cocaine sensitization. Biol. Psychiatry 2012, 71, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Yorgason, J.T.; Espana, R.A.; Konstantopoulos, J.K.; Weiner, J.L.; Jones, S.R. Enduring increases in anxiety-like behavior and rapid nucleus accumbens dopamine signaling in socially isolated rats. Eur. J. Neurosci. 2013, 37, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; An, S.; Tai, F.; Wang, J.; Wu, R.; Wang, B. Early social deprivation impairs pair bonding and alters serum corticosterone and the nacc dopamine system in mandarin voles. Psychoneuroendocrinology 2013, 38, 3128–3138. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Nelson, C.L.; Bruno, J.P. Cortical cholinergic transmission and cortical information processing in schizophrenia. Schizophr. Bull. 2005, 31, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Agoglia, A.E.; Sharko, A.C.; Psilos, K.E.; Holstein, S.E.; Reid, G.T.; Hodge, C.W. Alcohol alters the activation of erk1/2, a functional regulator of binge alcohol drinking in adult c57bl/6j mice. Alcohol. Clin. Exp. Res. 2015, 39, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Crews, F.T. Adolescent intermittent ethanol exposure enhances ethanol activation of the nucleus accumbens while blunting the prefrontal cortex responses in adult rat. Neuroscience 2015, 293, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, A.N.; Rose, J.H.; Huggins, K.N.; Konstantopoulos, J.K.; Jones, S.R. Chronic intermittent ethanol exposure reduces presynaptic dopamine neurotransmission in the mouse nucleus accumbens. Drug Alcohol Depend. 2015, 150, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Griffin, W.C.; Ramachandra, V.S.; Knackstedt, L.A.; Becker, H.C. Repeated cycles of chronic intermittent ethanol exposure increases basal glutamate in the nucleus accumbens of mice without affecting glutamate transport. Front. Pharmacol. 2015, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- McGuier, N.S.; Padula, A.E.; Mulholland, P.J.; Chandler, L.J. Homer2 deletion alters dendritic spine morphology but not alcohol-associated adaptations in glun2b-containing N-methyl-d-aspartate receptors in the nucleus accumbens. Front. Pharmacol. 2015, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Alsio, J.; Morud, J.; Ericson, M.; Robbins, T.W.; Soderpalm, B. Ethanol impairment of spontaneous alternation behaviour and associated changes in medial prefrontal glutamatergic gene expression precede putative markers of dependence. Pharmacol. Biochem. Behav. 2015, 132, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Uys, J.D.; McGuier, N.S.; Gass, J.T.; Griffin, W.C., 3rd; Ball, L.E.; Mulholland, P.J. Chronic intermittent ethanol exposure and withdrawal leads to adaptations in nucleus accumbens core postsynaptic density proteome and dendritic spines. Addict. Biol. 2015. [Google Scholar] [CrossRef]
- Peterson, V.L.; McCool, B.A.; Hamilton, D.A. Effects of ethanol exposure and withdrawal on dendritic morphology and spine density in the nucleus accumbens core and shell. Brain Res. 2015, 1594, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Dumontier, S.; DeRoode, D.; Sahota, P.; Thakkar, M.M. Nicotine infusion in the wake-promoting basal forebrain enhances alcohol-induced activation of nucleus accumbens. Alcohol. Clin. Exp. Res. 2014, 38, 2590–2596. [Google Scholar] [CrossRef] [PubMed]
- Ramaker, M.J.; Strong-Kaufman, M.N.; Ford, M.M.; Phillips, T.J.; Finn, D.A. Effect of nucleus accumbens shell infusions of ganaxolone or gaboxadol on ethanol consumption in mice. Psychopharmacology 2014, 232, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Tizabi, Y.; Copeland, R.L., Jr.; Louis, V.A.; Taylor, R.E. Effects of combined systemic alcohol and central nicotine administration into ventral tegmental area on dopamine release in the nucleus accumbens. Alcohol. Clin. Exp. Res. 2002, 26, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, S.E.; Quadros, I.M.; Takahashi, S.; Suchecki, D. Long maternal separation accelerates behavioural sensitization to ethanol in female, but not in male mice. Behav. Brain Res. 2007, 184, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Slotten, H.A.; Kalinichev, M.; Hagan, J.J.; Marsden, C.A.; Fone, K.C. Long-lasting changes in behavioural and neuroendocrine indices in the rat following neonatal maternal separation: Gender-dependent effects. Brain Res. 2006, 1097, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Renard, G.M.; Rivarola, M.A.; Suarez, M.M. Gender-dependent effects of early maternal separation and variable chronic stress on vasopressinergic activity and glucocorticoid receptor expression in adult rats. Dev. Neurosci. 2010, 32, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Cservenka, A.; Gillespie, A.J.; Michael, P.G.; Nagel, B.J. Family history density of alcoholism relates to left nucleus accumbens volume in adolescent girls. J. Stud. Alcohol Drugs 2015, 76, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Del Arco, A.; Mora, F. Neurotransmitters and prefrontal cortex-limbic system interactions: Implications for plasticity and psychiatric disorders. J. Neural Trans. (Vienna, Aus.: 1996) 2009, 116, 941–952. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, P.; Grace, A.A. Dysfunctions in multiple interrelated systems as the neurobiological bases of schizophrenic symptom clusters. Schizophr. Bull. 1998, 24, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Saddoris, M.P.; Sugam, J.A.; Cacciapaglia, F.; Carelli, R.M. Rapid dopamine dynamics in the accumbens core and shell: Learning and action. Front. Biosci. (Elite ed.) 2013, 5, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Bock, J.; Riedel, A.; Braun, K. Differential changes of metabolic brain activity and interregional functional coupling in prefronto-limbic pathways during different stress conditions: Functional imaging in freely behaving rodent pups. Front. Cell. Neurosci. 2012, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Brake, W.G.; Zhang, T.Y.; Diorio, J.; Meaney, M.J.; Gratton, A. Influence of early postnatal rearing conditions on mesocorticolimbic dopamine and behavioural responses to psychostimulants and stressors in adult rats. Eur. J. Neurosci. 2004, 19, 1863–1874. [Google Scholar] [CrossRef] [PubMed]
- Oreland, S.; Raudkivi, K.; Oreland, L.; Harro, J.; Arborelius, L.; Nylander, I. Ethanol-induced effects on the dopamine and serotonin systems in adult wistar rats are dependent on early-life experiences. Brain Res. 2011, 1405, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Kalpachidou, T.; Raftogianni, A.; Zografou, E.; Tzanou, A.; Pondiki, S.; Stylianopoulou, F. Rat dams exposed repeatedly to a daily brief separation from the pups exhibit increased maternal behavior, decreased anxiety and altered levels of receptors for estrogens (eralpha, erbeta), oxytocin and serotonin (5-ht1a) in their brain. Psychoneuroendocrinology 2015, 52, 212–228. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, S.E.; Quadros, I.M.; Machado, R.B.; Suchecki, D. Sex-dependent effects of maternal separation on plasma corticosterone and brain monoamines in response to chronic ethanol administration. Neuroscience 2013, 253, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Greisen, M.H.; Bolwig, T.G.; Wortwein, G. Cholecystokinin tetrapeptide effects on hpa axis function and elevated plus maze behaviour in maternally separated and handled rats. Behav. Brain Res. 2005, 161, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Plotsky, P.M.; Meaney, M.J. Early, postnatal experience alters hypothalamic corticotropin-releasing factor (crf) mrna, median eminence crf content and stress-induced release in adult rats. Mol. Brain Res. 1993, 18, 195–200. [Google Scholar] [CrossRef]
- Francis, D.D.; Diorio, J.; Plotsky, P.M.; Meaney, M.J. Environmental enrichment reverses the effects of maternal separation on stress reactivity. J. Neurosci. 2002, 22, 7840–7843. [Google Scholar] [PubMed]
- Nylander, I.; Roman, E. Is the rodent maternal separation model a valid and effective model for studies on the early-life impact on ethanol consumption? Psychopharmacology 2013, 229, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Skalkidou, A.; Hellgren, C.; Comasco, E.; Sylven, S.; Sundstrom Poromaa, I. Biological aspects of postpartum depression. Women’s health 2012, 8, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Bodo, C.; Rissman, E.F. New roles for estrogen receptor beta in behavior and neuroendocrinology. Front. Neuroendocrinol. 2006, 27, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Romens, S.E.; McDonald, J.; Svaren, J.; Pollak, S.D. Associations between early life stress and gene methylation in children. Child Dev. 2015, 86, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Oitzl, M.S.; Champagne, D.L.; van der Veen, R.; de Kloet, E.R. Brain development under stress: Hypotheses of glucocorticoid actions revisited. Neurosci. Biobehav. Rev. 2010, 34, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Alt, S.R.; Turner, J.D.; Klok, M.D.; Meijer, O.C.; Lakke, E.A.; Derijk, R.H.; Muller, C.P. Differential expression of glucocorticoid receptor transcripts in major depressive disorder is not epigenetically programmed. Psychoneuroendocrinology 2010, 35, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Turecki, G.; Meaney, M.J. Effects of the social environment and stress on glucocorticoid receptor gene methylation: A systematic review. Biological psychiatry 2014. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Szyf, M. Environmental programming of stress responses through DNA methylation: Life at the interface between a dynamic environment and a fixed genome. Dialogues Clin. Neurosci. 2005, 7, 103–123. [Google Scholar] [PubMed]
- Weaver, I.C. Epigenetic effects of glucocorticoids. Semin. Fetal Neonatal Med. 2009, 14, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Fish, E.W.; Shahrokh, D.; Bagot, R.; Caldji, C.; Bredy, T.; Szyf, M.; Meaney, M.J. Epigenetic programming of stress responses through variations in maternal care. Ann. NY Acad. Sci. 2004, 1036, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Witzmann, S.R.; Turner, J.D.; Meriaux, S.B.; Meijer, O.C.; Muller, C.P. Epigenetic regulation of the glucocorticoid receptor promoter 1(7) in adult rats. Epigenetics 2012, 7, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Krebs, M.O.; Bleich, S.; Frieling, H. Epigenetics and depression: Current challenges and new therapeutic options. Curr. Opin. Psychiatry 2010, 23, 588–592. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O.; Kato, T. Epigenetics in mood disorders. Environ. Health Prev. Med. 2008, 13, 16–24. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O. Epigenomic mechanisms of early adversity and hpa dysfunction: Considerations for ptsd research. Front. Psychiatry 2013, 4, 110. [Google Scholar] [CrossRef] [PubMed]
- Van der Knaap, L.J.; Riese, H.; Hudziak, J.J.; Verbiest, M.M.; Verhulst, F.C.; Oldehinkel, A.J.; van Oort, F.V. Glucocorticoid receptor gene (nr3c1) methylation following stressful events between birth and adolescence. The trails study. Transl. Psychiatry 2014, 4, e381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kloet, E.R.; Fitzsimons, C.P.; Datson, N.A.; Meijer, O.C.; Vreugdenhil, E. Glucocorticoid signaling and stress-related limbic susceptibility pathway: About receptors, transcription machinery and microrna. Brain Res. 2009, 1293, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Sofuoglu, M.; Mooney, M. Cholinergic functioning in stimulant addiction: Implications for medications development. CNS Drugs 2009, 23, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Avena, N.M.; Rada, P.V. Cholinergic modulation of food and drug satiety and withdrawal. Physiol. Behav. 2012, 106, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Mark, G.P.; Shabani, S.; Dobbs, L.K.; Hansen, S.T. Cholinergic modulation of mesolimbic dopamine function and reward. Physiol. Behav. 2011, 104, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.A.; Neves, J.; Vilela, M.; Sousa, S.; Cruz, C.; Madeira, M.D. Chronic alcohol consumption leads to neurochemical changes in the nucleus accumbens that are not fully reversed by withdrawal. Neurotoxicol. Teratol. 2014, 44, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Correa, M. The mysterious motivational functions of mesolimbic dopamine. Neuron 2012, 76, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Levita, L.; Dalley, J.W.; Robbins, T.W. Nucleus accumbens dopamine and learned fear revisited: A review and some new findings. Behav. Brain Res. 2002, 137, 115–127. [Google Scholar] [CrossRef]
- Floresco, S.B. The nucleus accumbens: An interface between cognition, emotion, and action. Ann. Rev. Psychol. 2015, 66, 25–52. [Google Scholar] [CrossRef] [PubMed]
- Carlezon, W.A., Jr.; Thomas, M.J. Biological substrates of reward and aversion: A nucleus accumbens activity hypothesis. Neuropharmacology 2009, 56 (Suppl. S1), 122–132. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Mesulam, M.M.; Mufson, E.J.; Levey, A.I.; Wainer, B.H. Cholinergic innervation of cortex by the basal forebrain: Cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (substantia innominata), and hypothalamus in the rhesus monkey. J. Comp. Neurol. 1983, 214, 170–197. [Google Scholar] [CrossRef] [PubMed]
- Rye, D.B.; Wainer, B.H.; Mesulam, M.M.; Mufson, E.J.; Saper, C.B. Cortical projections arising from the basal forebrain: A study of cholinergic and noncholinergic components employing combined retrograde tracing and immunohistochemical localization of choline acetyltransferase. Neuroscience 1984, 13, 627–643. [Google Scholar] [CrossRef]
- Blomqvist, O.; Ericson, M.; Engel, J.A.; Soderpalm, B. Accumbal dopamine overflow after ethanol: Localization of the antagonizing effect of mecamylamine. Eur. J. Pharmacol. 1997, 334, 149–156. [Google Scholar] [CrossRef]
- Soderpalm, B.; Lof, E.; Ericson, M. Mechanistic studies of ethanol’s interaction with the mesolimbic dopamine reward system. Pharmacopsychiatry 2009, 42 (Suppl. S1), 87–94. [Google Scholar] [CrossRef] [PubMed]
- Doyon, W.M.; Dong, Y.; Ostroumov, A.; Thomas, A.M.; Zhang, T.A.; Dani, J.A. Nicotine decreases ethanol-induced dopamine signaling and increases self-administration via stress hormones. Neuron 2013, 79, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Ericson, M.; Blomqvist, O.; Engel, J.A.; Soderpalm, B. Voluntary ethanol intake in the rat and the associated accumbal dopamine overflow are blocked by ventral tegmental mecamylamine. Eur. J. Pharmacol. 1998, 358, 189–196. [Google Scholar] [CrossRef]
- Larsson, A.; Edstrom, L.; Svensson, L.; Soderpalm, B.; Engel, J.A. Voluntary ethanol intake increases extracellular acetylcholine levels in the ventral tegmental area in the rat. Alcohol Alcohol. 2005, 40, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Zoli, M.; Clementi, F. Brain nicotinic acetylcholine receptors: Native subtypes and their relevance. Trends Pharmacol. Sci. 2006, 27, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Mansvelder, H.D.; Keath, J.R.; McGehee, D.S. Synaptic mechanisms underlie nicotine-induced excitability of brain reward areas. Neuron 2002, 33, 905–919. [Google Scholar] [CrossRef]
- Soderpalm, B.; Ericson, M. Neurocircuitry involved in the development of alcohol addiction: The dopamine system and its access points. Curr. Top. Behav. Neurosci. 2013, 13, 127–161. [Google Scholar] [PubMed]
- Tabakoff, B.; Hoffman, P.L. The neurobiology of alcohol consumption and alcoholism: An integrative history. Pharmacol. Biochem. Behav. 2013, 113, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Hopur52009. Nicotinic_receptors.Png. Available online: http://upload.wikimedia.org/wikipedia/commons/f/f1/Nicotinic_receptors.png (accessed on 2 March 2015).
- Josephine, T.M.S.; Holgate, J.; Bartlett, S.E. Effects of alcohol on nicotinic acetylcholine receptors and impact on addiction. Neuropathol. Addict. 2014, in press. [Google Scholar]
- Zaborszky, L.; Hoemke, L.; Mohlberg, H.; Schleicher, A.; Amunts, K.; Zilles, K. Stereotaxic probabilistic maps of the magnocellular cell groups in human basal forebrain. NeuroImage 2008, 42, 1127–1141. [Google Scholar] [CrossRef] [PubMed]
- Zaborszky, L.; Pang, K.; Somogyi, J.; Nadasdy, Z.; Kallo, I. The basal forebrain corticopetal system revisited. Ann. NY Acad. Sci. 1999, 877, 339–367. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Prendergast, M.A. Cholinergic receptor system as a target for treating alcohol abuse and dependence. Recent Pat. CNS Drug Discov. 2012, 7, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Feduccia, A.A.; Chatterjee, S.; Bartlett, S.E. Neuronal nicotinic acetylcholine receptors: Neuroplastic changes underlying alcohol and nicotine addictions. Front. Mol. Neurosci. 2012, 5, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çelik, T.; Kayir, H.; Ceyhan, M.; Demirtaş, S.; Coşar, A.; Uzbay, I.T. Cpp and amlodipine alter the decrease in basal acetylcholine and choline release by audiogenic stimulus in hippocampus of ethanol-withdrawn rats in vivo. Brain Res. Bull. 2004, 64, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Casamenti, F.; Scali, C.; Vannucchi, M.G.; Bartolini, L.; Pepeu, G. Long-term ethanol consumption by rats: Effect on acetylcholine release in vivo, choline acetyltransferase activity, and behavior. Neuroscience 1993, 56, 465–471. [Google Scholar] [CrossRef]
- Mitchell, J.M.; Teague, C.H.; Kayser, A.S.; Bartlett, S.E.; Fields, H.L. Varenicline decreases alcohol consumption in heavy-drinking smokers. Psychopharmacology 2012, 223, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Steensland, P.; Simms, J.A.; Holgate, J.; Coe, J.W.; Hurst, R.S.; Shaffer, C.L.; Lowe, J.; Rollema, H.; Bartlett, S.E. Partial agonists of the alpha3beta4* neuronal nicotinic acetylcholine receptor reduce ethanol consumption and seeking in rats. Neuropsychopharmacology 2011, 36, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Steensland, P.; Simms, J.A.; Holgate, J.; Richards, J.K.; Bartlett, S.E. Varenicline, an alpha4beta2 nicotinic acetylcholine receptor partial agonist, selectively decreases ethanol consumption and seeking. Proc. Natl. Acad. Sci. USA 2007, 104, 12518–12523. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, A.; Jerlhag, E.; Liljequist, S.; Engel, J. Effects of subunit selective nach receptors on operant ethanol self-administration and relapse-like ethanol-drinking behavior. Psychopharmacology 2009, 203, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Markett, S.; Melchers, M.; Montag, C. Interaction of the cholinergic system and the hypothalamic-pituitary-adrenal axis as a risk factor for depression: Evidence from a genetic association study. Neuroreport 2012, 23, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Aisa, B.; Gil-Bea, F.J.; Marcos, B.; Tordera, R.; Lasheras, B.; del Rio, J.; Ramirez, M.J. Neonatal stress affects vulnerability of cholinergic neurons and cognition in the rat: Involvement of the hpa axis. Psychoneuroendocrinology 2009, 34, 1495–1505. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Yamaguchi-Shima, N.; Shimizu, T.; Arai, J.; Lianyi, L.; Wakiguchi, H.; Yokotani, K. Role of brain nicotinic acetylcholine receptor in centrally administered corticotropin-releasing factor-induced elevation of plasma corticosterone in rats. European Journal of Pharmacol. 2008, 587, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Loomis, S.; Gilmour, G. Corticosterone urinalysis and nicotinic receptor modulation in rats. J. Neurosci. Methods 2010, 188, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Kim, J.W.; Song, J.Y.; Park, S.; Lee, H.J.; Chung, J.H. Association of polymorphisms in nicotinic acetylcholine receptor alpha 4 subunit gene (chrna4), mu-opioid receptor gene (oprm1), and ethanol-metabolizing enzyme genes with alcoholism in korean patients. Alcohol 2004, 34, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.M.; Andrud, K.M.; Burke, M.B.; Pearson, J.N.; Kreisler, A.D.; Stevens, K.E.; Leonard, S.; Adams, C.E. The effects of prenatal stress on alpha4 beta2 and alpha7 hippocampal nicotinic acetylcholine receptor levels in adult offspring. Dev. Neurobiol. 2013, 73, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Cservenka, A.; Casimo, K.; Fair, D.A.; Nagel, B.J. Resting state functional connectivity of the nucleus accumbens in youth with a family history of alcoholism. Psychiatry Res. 2014, 221, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Weiland, B.J.; Welsh, R.C.; Yau, W.Y.; Zucker, R.A.; Zubieta, J.K.; Heitzeg, M.M. Accumbens functional connectivity during reward mediates sensation-seeking and alcohol use in high-risk youth. Drug Alcohol Depend. 2013, 128, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.M.; Meda, S.A.; Thomas, A.D.; Potenza, M.N.; Krystal, J.H.; Worhunsky, P.; Stevens, M.C.; O’Malley, S.; Book, G.A.; Reynolds, B.; et al. Individuals family history positive for alcoholism show functional magnetic resonance imaging differences in reward sensitivity that are related to impulsivity factors. Biol. Psychiatry 2011, 69, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Markett, S.; Montag, C.; Reuter, M. The nicotinic acetylcholine receptor gene chrna4 is associated with negative emotionality. Emotion (Washington, DC) 2011, 11, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Cippitelli, A.; Brunori, G.; Gaiolini, K.A.; Zaveri, N.T.; Toll, L. Pharmacological stress is required for the anti-alcohol effect of the alpha3beta4* nachr partial agonist at-1001. Neuropharmacology 2015, 93c, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Baier, C.J.; Pallares, M.E.; Adrover, E.; Monteleone, M.C.; Brocco, M.A.; Barrantes, F.J.; Antonelli, M.C. Prenatal restraint stress decreases the expression of alpha-7 nicotinic receptor in the brain of adult rat offspring. Stress (Amst., Neth.) 2015. [Google Scholar] [CrossRef] [PubMed]
- Taslim, N.; Soderstrom, K.; Dar, M.S. Role of mouse cerebellar nicotinic acetylcholine receptor (nachr) alpha(4)beta(2)- and alpha(7) subtypes in the behavioral cross-tolerance between nicotine and ethanol-induced ataxia. Behav. Brain Res. 2011, 217, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Pichat, P.; Bergis, O.E.; Terranova, J.P.; Urani, A.; Duarte, C.; Santucci, V.; Gueudet, C.; Voltz, C.; Steinberg, R.; Stemmelin, J.; et al. Ssr180711, a novel selective alpha7 nicotinic receptor partial agonist: (ii) Efficacy in experimental models predictive of activity against cognitive symptoms of schizophrenia. Neuropsychopharmacology 2007, 32, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Robles, N.; Sabria, J. Ethanol consumption produces changes in behavior and on hippocampal alpha7 and alpha4beta2 nicotinic receptors. J. Mol. Neurosci. MN 2006, 30, 119–120. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holgate, J.Y.; Bartlett, S.E. Early Life Stress, Nicotinic Acetylcholine Receptors and Alcohol Use Disorders. Brain Sci. 2015, 5, 258-274. https://doi.org/10.3390/brainsci5030258
Holgate JY, Bartlett SE. Early Life Stress, Nicotinic Acetylcholine Receptors and Alcohol Use Disorders. Brain Sciences. 2015; 5(3):258-274. https://doi.org/10.3390/brainsci5030258
Chicago/Turabian StyleHolgate, Joan Y., and Selena E. Bartlett. 2015. "Early Life Stress, Nicotinic Acetylcholine Receptors and Alcohol Use Disorders" Brain Sciences 5, no. 3: 258-274. https://doi.org/10.3390/brainsci5030258