
Morphology of Donor and Recipient Nerves Utilised in Nerve Transfers to Restore Upper Limb Function in Cervical Spinal Cord Injury

 ,
,
Abstract
:1. Introduction
2. Patients
3. Materials and Methods
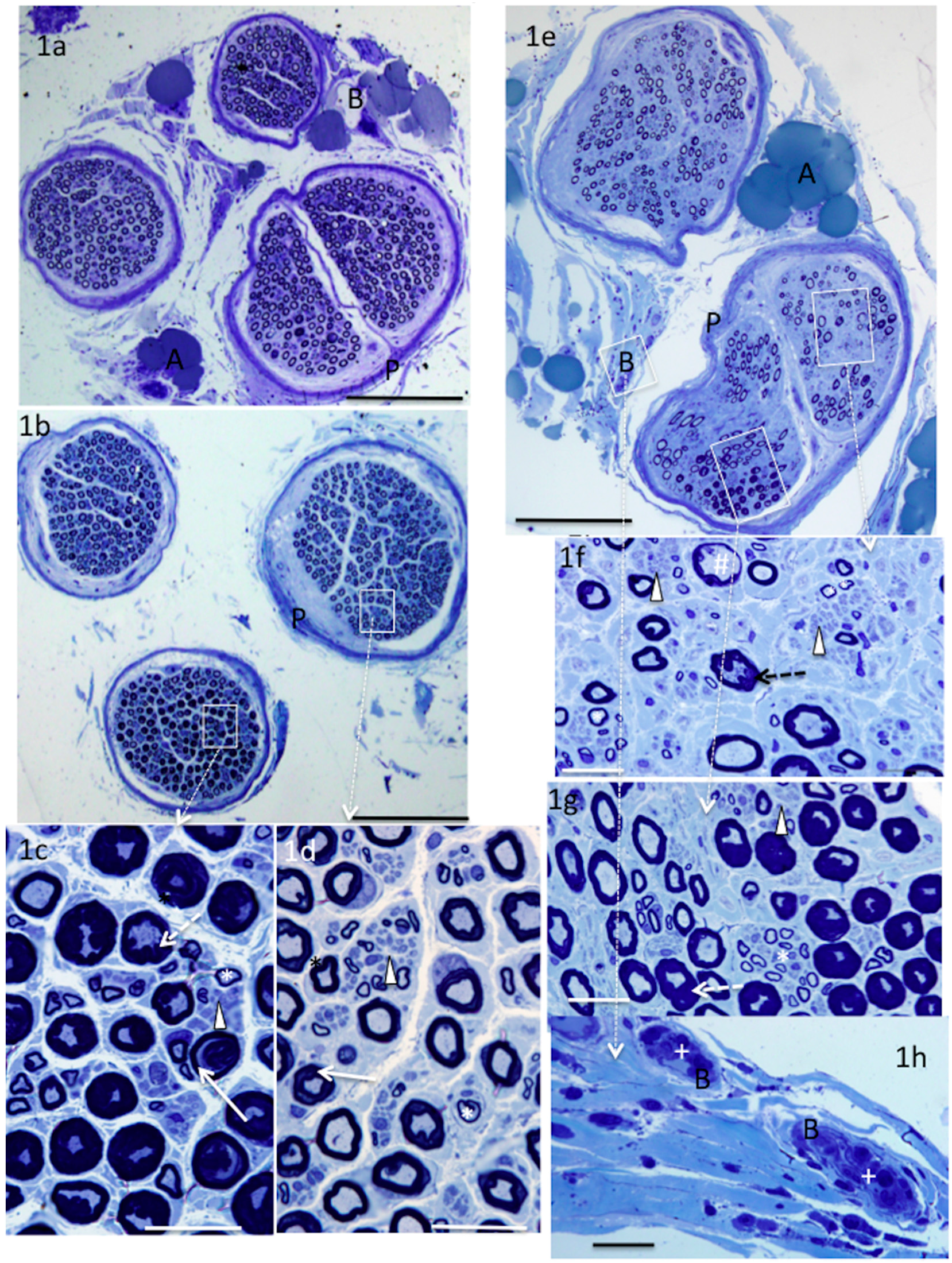
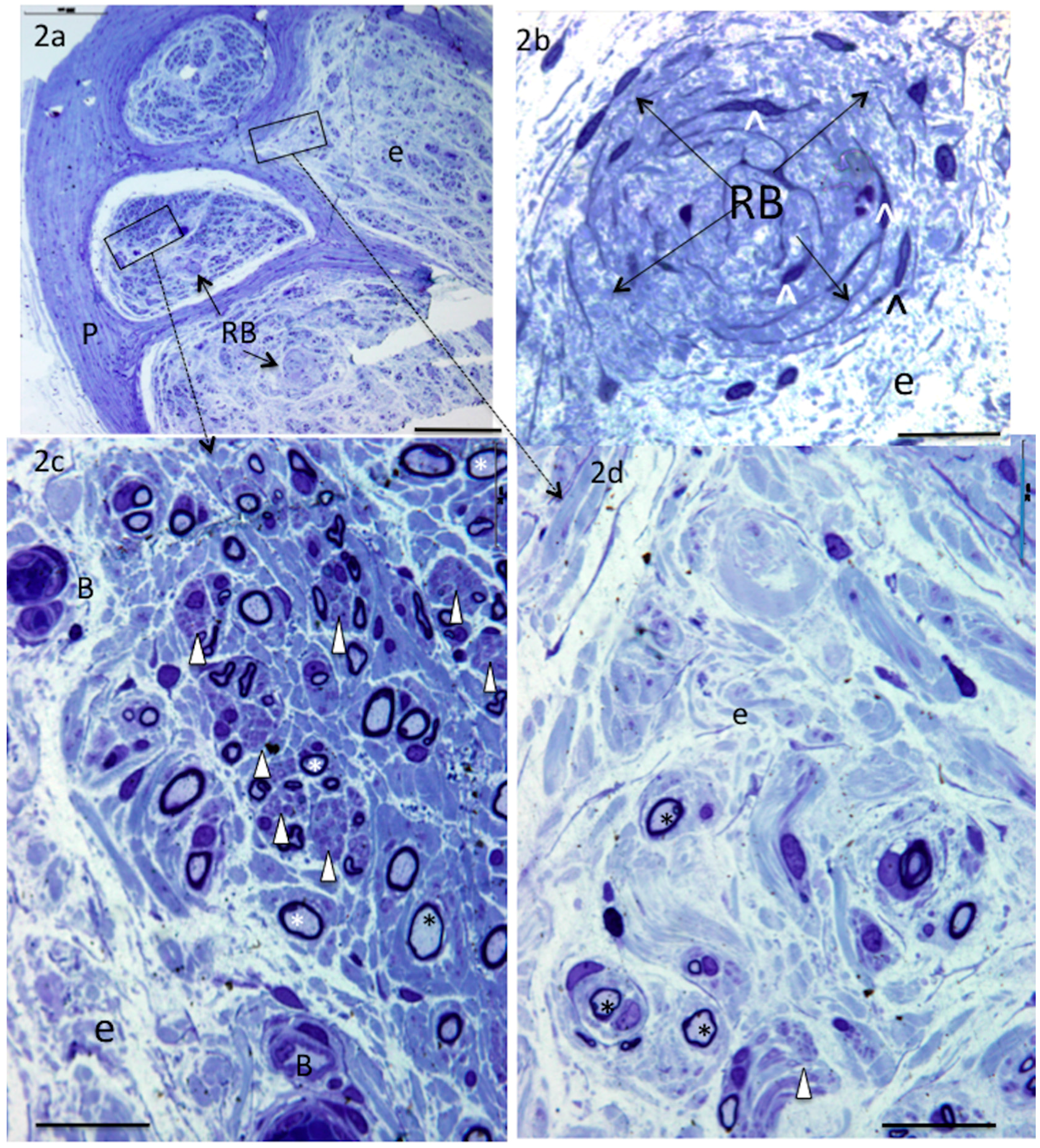
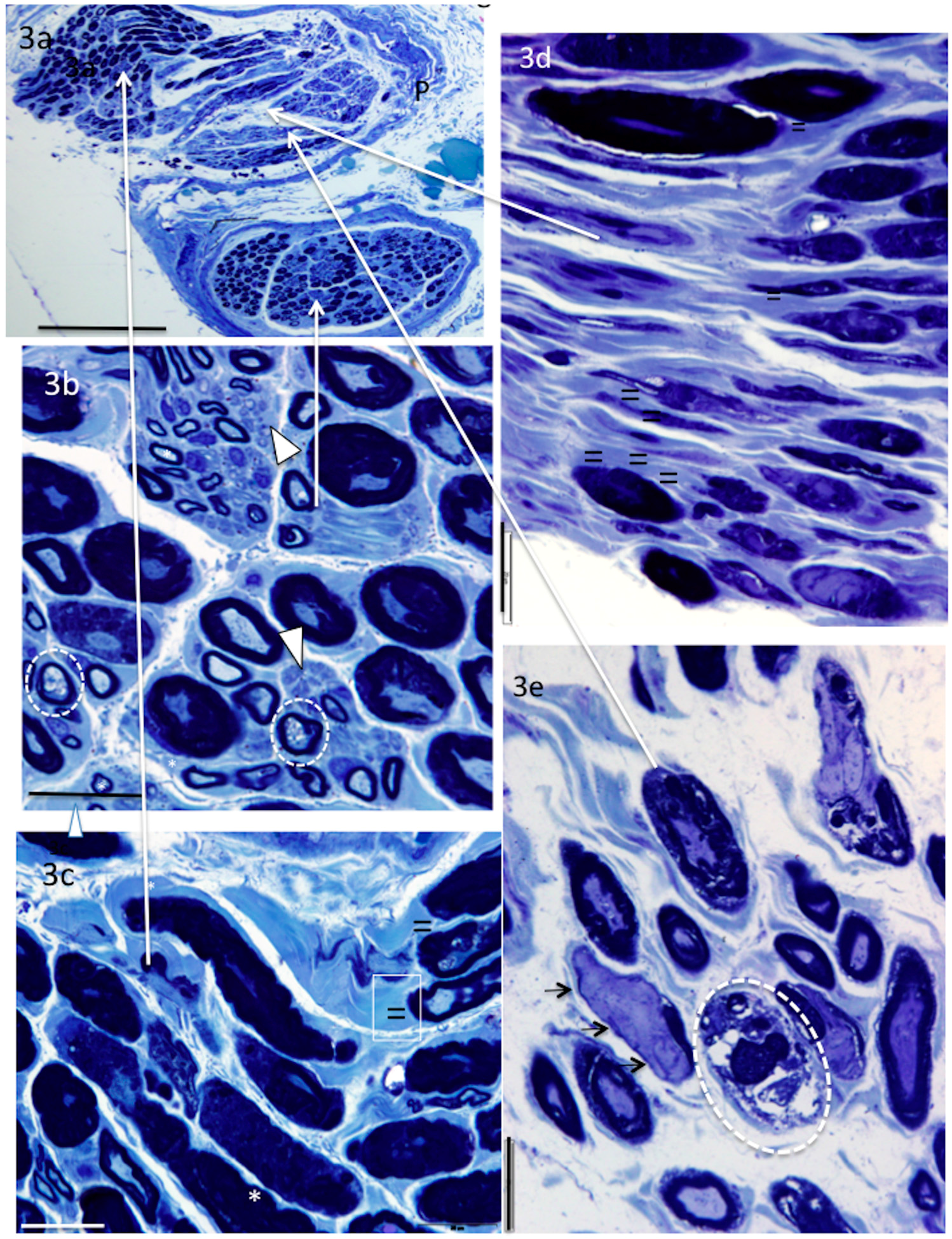
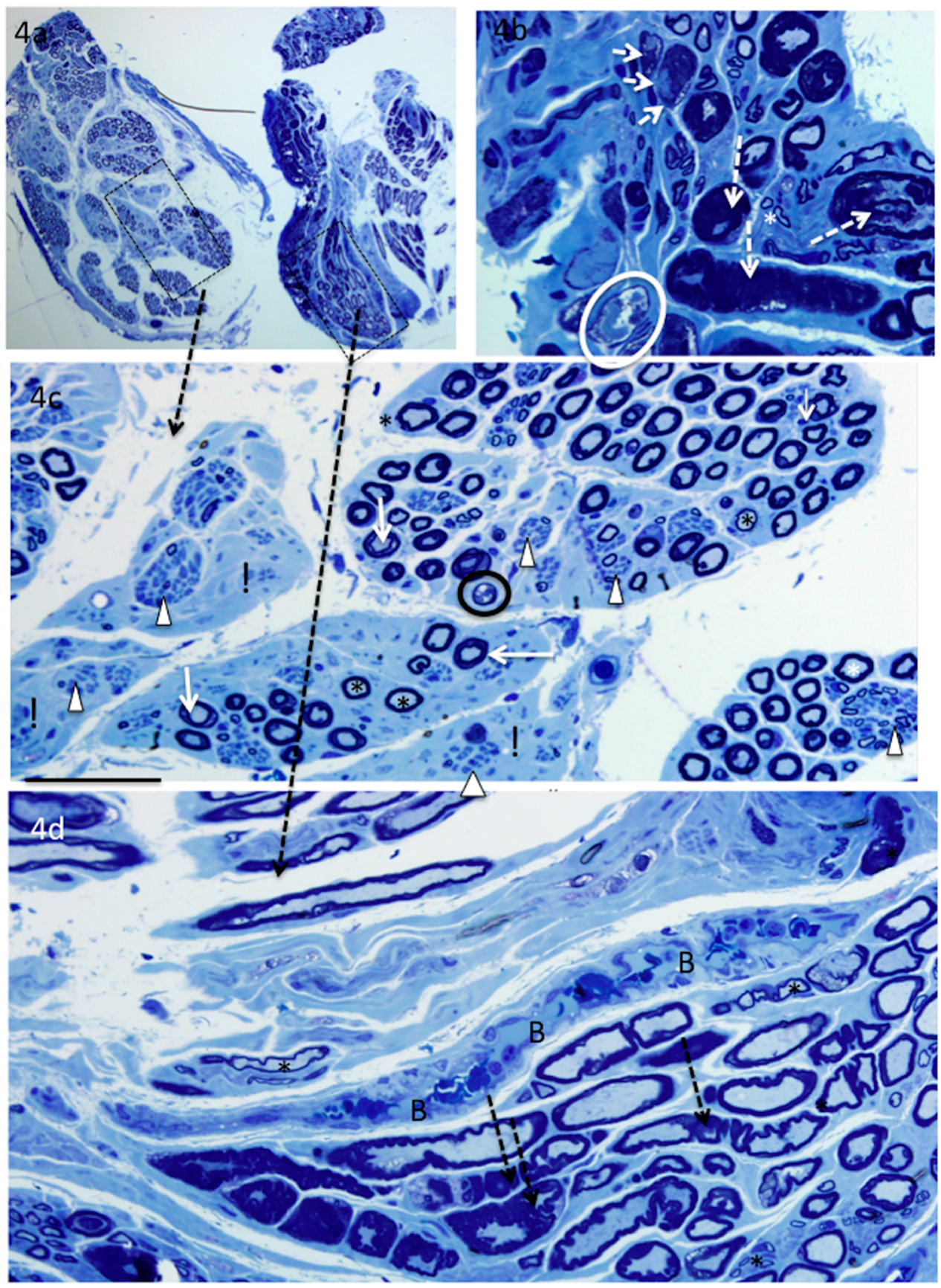
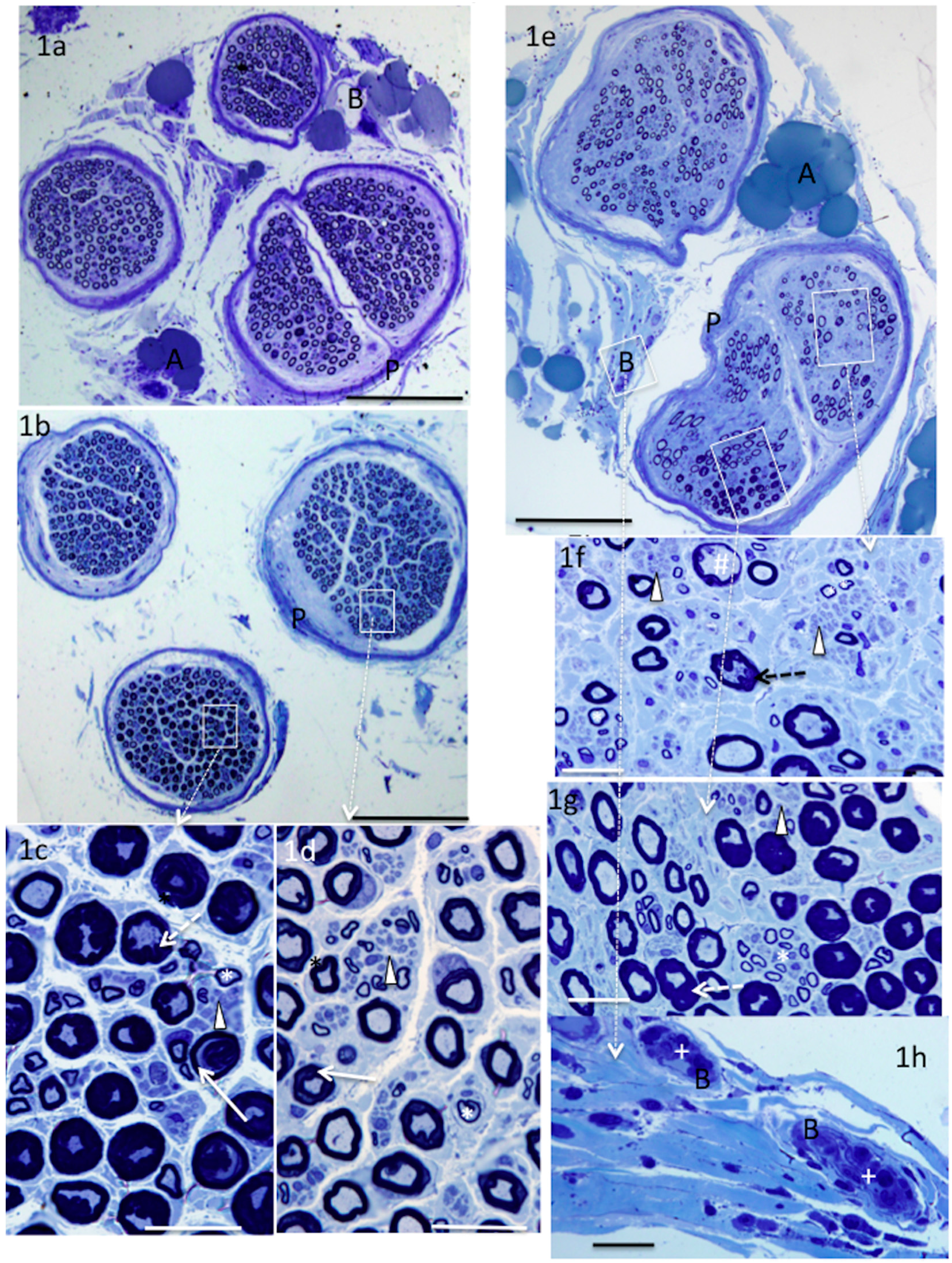
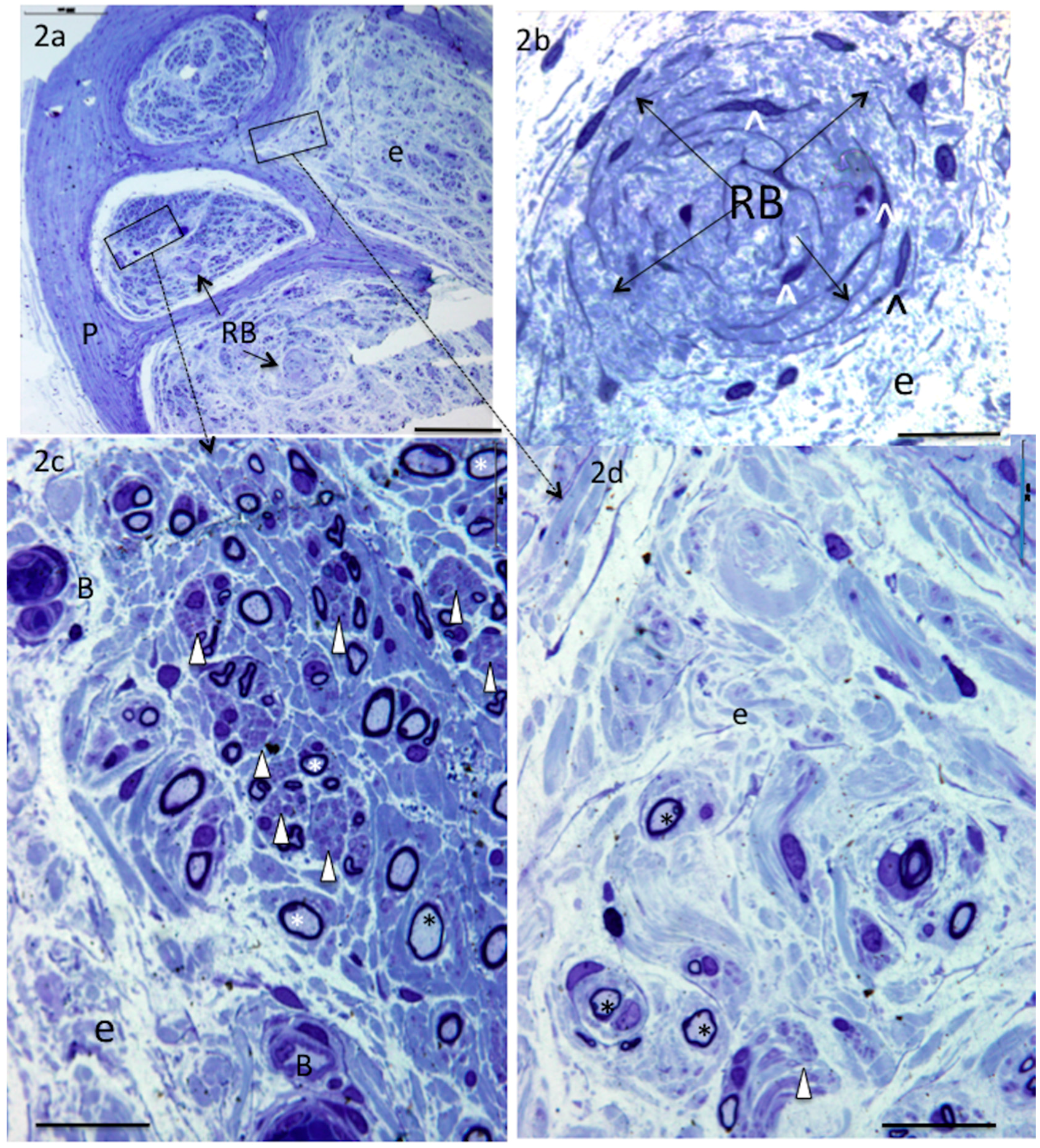
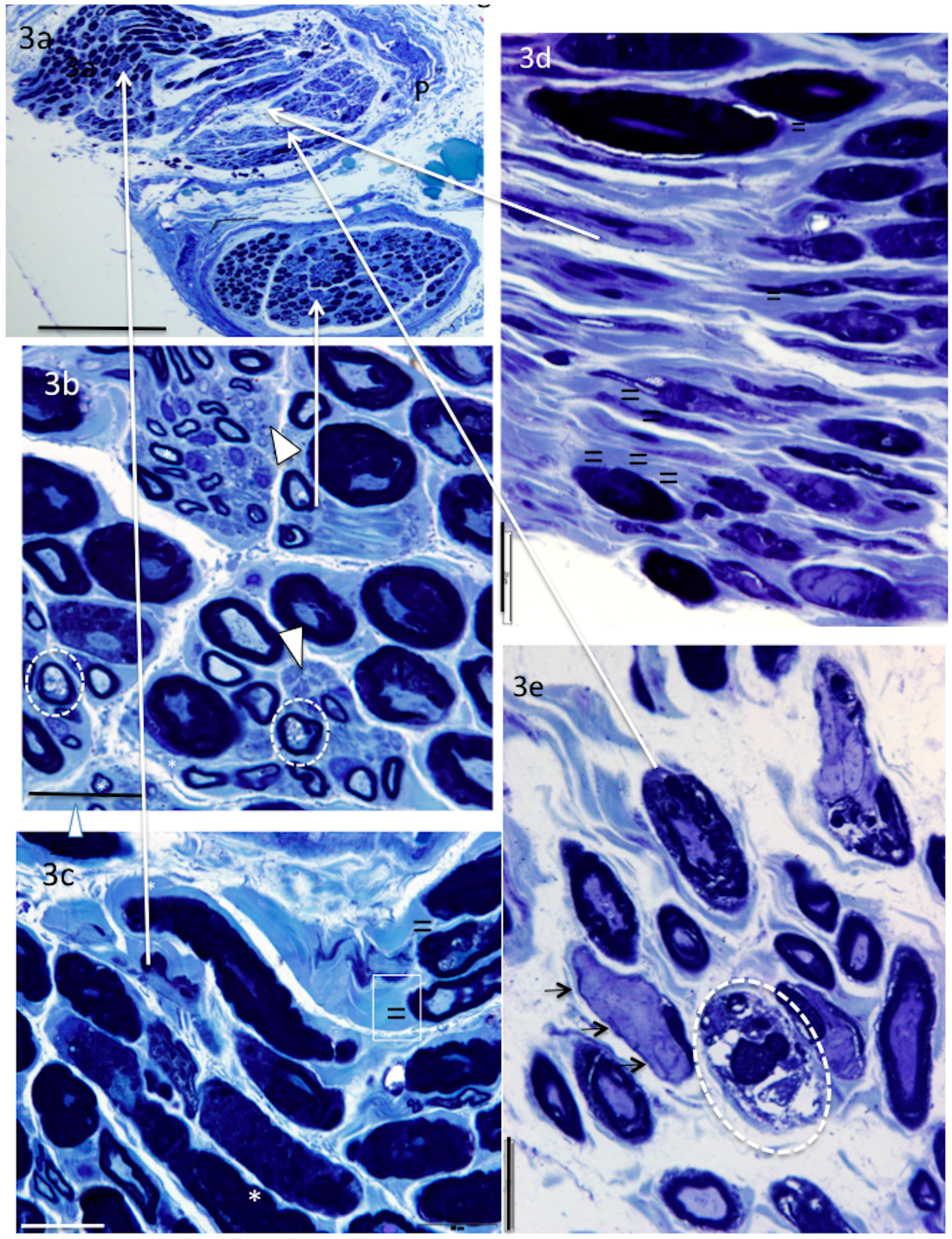
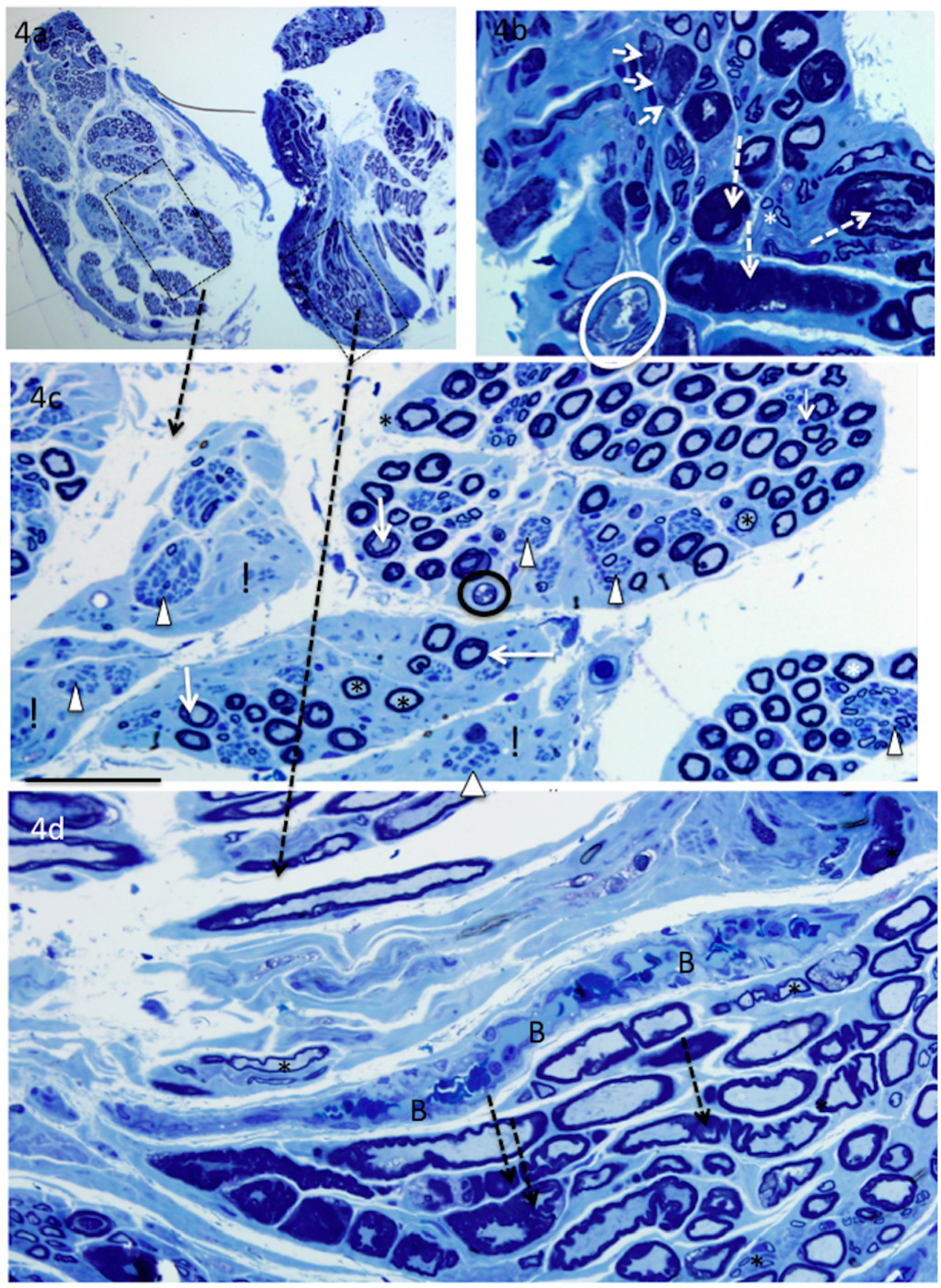
4. Results
5. Donor Nerves
5.1. Supinator (n = 4)
5.2. Brachialis (n = 1) and ECRB (n = 1)
5.3. Teres minor (n = 3) and Pax (n = 2)
6. Recipient Nerves
6.1. PIN (n = 4) and AIN (n = 2)
6.2. Triceps (n = 3)
7. Discussion
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Zyl, N.; Hahn, J.B.; Cooper, C.A.; Weymouth, M.D.; Flood, S.J.; Galea, M.P. Upper limb reinnervation in c6 tetraplegia using a triple nerve transfer: Case report. J. Hand Surg. Am. 2014, 39, 1779–1783. [Google Scholar] [CrossRef] [PubMed]
- Cain, S.; Gohritz, A.; Fridén, J.; van Zyl, N. Review of upper extremity nerve transfer in cervical spinal cord injury. J. Brachial Plex. Peripher. Nerve Inj. 2015, 10, e34–e42. [Google Scholar] [CrossRef]
- Bertelli, J.A.; Ghizoni, M.F.; Tacca, C.P. Transfer of the teres minor motor branch for triceps reinnervation in tetraplegia. J. Neurosurg. 2011, 114, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, J.A.; Ghizoni, M.F. Transfer of supinator motor branches to the posterior interosseous nerve in C7-T1 brachial plexus palsy. J. Neurosurg. 2010, 113, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, J.A.; Mendes Lehm, V.L.; Tacca, C.P.; Winkelmann Duarte, E.C.; Ghizoni, M.F.; Duarte, H. Transfer of the distal terminal motor branch of the extensor carpi radialis brevis to the nerve of the flexor pollicis longus: An anatomic study and clinical application in a tetraplegic patient. Neurosurgery 2012, 70, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, S.E.; Yee, A.; Ray, W.Z. Nerve transfers for the restoration of hand function after spinal cord injury. J. Neurosurg. 2012, 117, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M. Nerve transfers in tetraplegia i: Background and technique. Surg. Neurol. Int. 2011, 2, 121. [Google Scholar] [CrossRef] [PubMed]
- Council, M.R. Aids to the Examination of the Peripheral Nervous System; Saunders Ltd.: London, UK, 1976. [Google Scholar]
- Brown, J.M.; Vivio, N.; Sheean, G.L. The clinical practice of reconstructive neurosurgery. Clin. Neurol. Neurosurg. 2012, 114, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Coulet, B.; Allieu, Y.; Chammas, M. Injured metamere and functional surgery of the tetraplegic upper limb. Hand Clin. 2002, 18, 399–412. [Google Scholar] [CrossRef]
- Tankisi, H.; Pugdahl, K.; Rasmussen, M.M.; Clemmensen, D.; Rawashdeh, Y.F.; Christensen, P.; Krogh, K.; Fuglsang-Frederiksen, A. Peripheral nervous system involvement in chronic spinal cord injury. Muscle Nerve 2015, 52, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Kirshblum, S.; Lim, S.; Garstang, S.; Millis, S. Electrodiagnostic changes of the lower limbs in subjects with chronic complete cervical spinal cord injury. Arch. Phys. Med. Rehabil. 2001, 82, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Kakulas, B.A. Neuropathology: The foundation for new treatments in spinal cord injury. Spinal Cord 2004, 42, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Fox, I.K.; Davidge, K.M.; Novak, C.B.; Hoben, G.; Kahn, L.C.; Juknis, N.; Ruvinskaya, R.; Mackinnon, S.E. Use of peripheral nerve transfers in tetraplegia: Evaluation of feasibility and morbidity. Hand (N. Y.) 2015, 10, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, J.A.; Tacca, C.P.; Winkelmann Duarte, E.C.; Ghizoni, M.F.; Duarte, H. Transfer of axillary nerve branches to reconstruct elbow extension in tetraplegics: A laboratory investigation of surgical feasibility. Microsurgery 2011, 31, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, S.E.; Dellon, A.L.; Hudson, A.R.; Hunter, D.A. Chronic human nerve compression a histolgical assessment. Neuropathol. Appl. Neurobiol. 1986, 12, 547–565. [Google Scholar] [CrossRef] [PubMed]
- Renaut, J. Recherche sur quelque points particulier d’histologie des nerfs. Arch. Physiol. (Paris) 1881, 8, 846–859. [Google Scholar]
- Asbury, A.K. Renaut bodies. A forgotten endoneurial structure. J. Neuropathol. Exp. Neurol. 1973, 32, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Pina-Oviedo, S.; Del Valle, L.; Baquera-Heredia, J.; Ortiz-Hidalgo, C. Immunohistochemical characterization of Renaut bodies in superficial digital nerves: Further evidence supporting their perineurial cell origin. J. Peripher. Nerv. Syst. 2009, 14, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, D.; Neary, D.; Eames, R.A. Renaut body distributiion at sites of human peripheral nerve entrapment. J. Neurol. Sci. 1981, 49, 19–29. [Google Scholar] [CrossRef]
- Neary, D.; Ochoa, J.; Gilliatt, R.W. Subclinical entrapment neuropathy in man. J. Neurol. Sci. 1975, 24, 283–298. [Google Scholar] [CrossRef]
- Gitlin, G. Concerning the ganglioform enlargement (‘psuedoganglion’) on the nerve to teres minor muscle. Anatomy 1957, 91, 466–470. [Google Scholar]
- Linker, C.S.; Helms, C.A.; Fritz, R.C. Quadrilateral space syndrome: Findings at MR imaging. Radiology 1993, 188, 675–676. [Google Scholar] [CrossRef] [PubMed]
- Cothran, R.L., Jr.; Helms, C. Quadrilateral space syndrome: Incidence of imaging findings in a population referred for MRI of the shoulder. AJR Am. J. Roentgenol. 2005, 184, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Hamada, T.; Toshitaka, N.; Yasuyuli, S.; Katsumi, S. Myxoid and globular degeneration of nerves in the shoulder joint. J. Clin. Orthop. Relat. Res. 1995, 320, 55–64. [Google Scholar] [CrossRef]
- Hashimoto, T.; Hijioka, A.; Hamada, T.; Sasaguri, Y.; Suzuki, K. Quantitative study of myxoid and hyaline-globular degeneration of nerves in the shoulder joint. Clin. Orthop. Relat. Res. 1995, 19, 97–102. [Google Scholar]
- Sunderland, S. A classification of peripheral nerve injuries producing loss of function. Brain 1951, 74, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Castro, E.; Navarro, X. Peripheral nerve alterations after spinal cord injury in the adult rat. Spinal Cord 2013, 51, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Trojaborg, W.; Hays, A.P.; van den Berg, L.; Younger, D.S.; Latov, N. Motor conduction parameters in neuropathies associated with anti-mag antibodies and other types of demyelinating and axonal neuropathies. Muscle Nerve 1995, 18, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Sha, D.; Mohammed, A.A.; Asress, S.; Glass, J.D.; Chin, L.S.; Li, L. Motor and sensory neuropathy due to myelin infolding and paranodal damage in a transgenic mouse model of charcot-marie-tooth disease type 1c. Hum. Mol. Genet. 2013, 22, 1755–1770. [Google Scholar] [CrossRef] [PubMed]
- Maier, M.; Berger, P.; Suter, U. Understanding schwann cell-neurone interactions: The key to charcot-marie-tooth disease? J. Anat. 2002, 200, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Berger, P.; Niemann, A.; Suter, U. Schwann cells and the pathogenesis of inherited motor and sensory neuropathies (charcot-marie-tooth disease). Glia 2006, 54, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, F.; Pisciotta, C.; Reilly, M.M.; Tozza, S.; Schenone, A.; Fabrizi, G.M.; Cavallaro, T.; Vita, G.; Padua, L.; Gemignani, F.; et al. Nerve conduction velocity in CMT1A: What else can we tell? Eur. J. Neurol. 2016, 23, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Pareyson, D.; Scaioli, V.; Laura, M. Clinical and electrophysiological aspects of charcot-marie-tooth disease. Neuromol. Med. 2006, 8, 3–22. [Google Scholar] [CrossRef]
- Lev-Ram, V.; Ellisman, M.H. Axonal activation-induced calcium transients in myelinating schwann cells, sources, and mechanisms. J. Neurosci. 1995, 15, 2628–2637. [Google Scholar] [PubMed]
- Reist, N.E.; Smith, S.J. Neurally evoked calcium transients in terminal schwann cells at the neuromuscular junction. Proc. Natl. Acad. Sci. USA 1992, 89, 7625–7629. [Google Scholar] [CrossRef] [PubMed]
- Popko, B. Myelin maintenance: Axonal support required. Nat. Neurosci. 2010, 13, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Dahlstrom, A.; Heiwall, P.O.; Booj, S.; Dahllof, A.G. The influence of supraspinal impulse activity on the intra-axonal transport of acetylcholine, choline acetyltransferase and acetylcholinesterase in rat motorneurons. Acta Physiol. Scand. 1978, 103, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kiernan, M.C.; Macefield, V.G.; Lee, B.B.; Lin, C.S. Short-term peripheral nerve stimulation ameliorates axonal dysfunction after spinal cord injury. J. Neurophysiol. 2015, 113, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.A.; Burns, A.S.; Carrion-Jones, M.; Dillingham, T.R. Electrophysiological dysfunction in the peripheral nervous system following spinal cord injury. PM R 2011, 3, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Van De Meent, H.; Hosman, A.J.; Hendriks, J.; Zwarts, M.; Schubert, M. Severe degeneration of peripheral motor axons after spinal cord injury: A European multicenter study in 345 patients. Neurorehabil. Neural Repair 2010, 24, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Blaik, Z.; McGarry, J.; Daura, R. Peripheral neuropathy in spinal cord injured patients. Electromyogr. Clin. Neurophysiol. 1989, 29, 469–472. [Google Scholar] [PubMed]
- Kaelan, C.; Jacobsen, P.F.; Kakulas, B.A. An investigation of possible transynaptic neuronal degeneration in human spinal cord injury. J. Neurol. Sci. 1988, 86, 231–237. [Google Scholar] [CrossRef]
- Bjugn, R.; Nyengaard, J.R.; Rosland, J.H. Spinal cord transection—No loss of distal ventral horn neurons. Modern stereological techniques reveal no transneuronal changes in the ventral horns of the mouse lumbar spinal cord after thoracic cord transection. Exp. Neurol. 1997, 148, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Laurence, T.N.; Pugel, A.V.; Teasdall, R.D. Peripheral nerve involvement in spinal cord injury: An electromyographic study. Arch. Phys. Med. Rehabil. 1978, 59, 309–313. [Google Scholar] [PubMed]
- Lin, C.S.; Macefield, V.G.; Elam, M.; Wallin, B.G.; Engel, S.; Kiernan, M.C. Axonal changes in spinal cord injured patients distal to the site of injury. Brain 2007, 130, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K.C.; Hull, T.C.; Delaney, G.A.; Potter, P.J.; Sequeira, K.A.; Campbell, K.; Popovich, P.G. Elevated serum titers of proinflammatory cytokines and cns autoantibodies in patients with chronic spinal cord injury. J. Neurotrauma 2002, 19, 753–761. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Number | Patient Details |
|---|---|
| 3 | Male age 36 years |
| TPI 46 weeks | |
| C5 ASIA B | |
| 6 | Female Age 33 |
| TPI 46 weeks | |
| C5 ASIA A (C6 zone of partial preservation) | |
| 7 | Male age 23 years |
| TPI 46 weeks | |
| C5 ASIA A (C5 zone of partial preservation) |
| Spec No | Nerve | Axon Density | % Abnormal Myelin | Myelin | UM | Renaut body | Inflammation | Orientation |
|---|---|---|---|---|---|---|---|---|
| Recipient nerves | ||||||||
| 3L | PIN | + | T | |||||
| 3R | PIN | 89% | + | TL | ||||
| 6R | PIN | 100% | + | TL | ||||
| 7R | PIN | 58% | + | T | ||||
| 3R | AIN | |||||||
| 6R | AIN | 58% | + | TL | ||||
| 3R | TRI | - | +++ | T | ||||
| 6R | TRI | +++ | TL | |||||
| 7R | TRI | 69% | +++ | T | ||||
| Donor nerves | ||||||||
| 3L | SUP | - | + | T | ||||
| 3R | SUP | - | + | T | ||||
| 6R | SUP | + | T | |||||
| 7R | SUP | - | + | T | ||||
| 3R | BRA | - | + | T | ||||
| 6R | ECRB | - | ++ | T | ||||
| 3R | TM | - | ++ | T | ||||
| 6R | TM | - | +++ | T | ||||
| 7R | TM | - | ++ | TL | ||||
| 6R | PAX | - | +++ | TL | ||||
| 7R | PAX | - | + | T | ||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, A.; Van Zyl, N.; Weymouth, M.; Flood, S.; Nunn, A.; Cooper, C.; Hahn, J.; Galea, M.P. Morphology of Donor and Recipient Nerves Utilised in Nerve Transfers to Restore Upper Limb Function in Cervical Spinal Cord Injury. Brain Sci. 2016, 6, 42. https://doi.org/10.3390/brainsci6040042
Messina A, Van Zyl N, Weymouth M, Flood S, Nunn A, Cooper C, Hahn J, Galea MP. Morphology of Donor and Recipient Nerves Utilised in Nerve Transfers to Restore Upper Limb Function in Cervical Spinal Cord Injury. Brain Sciences. 2016; 6(4):42. https://doi.org/10.3390/brainsci6040042
Chicago/Turabian StyleMessina, Aurora, Natasha Van Zyl, Michael Weymouth, Stephen Flood, Andrew Nunn, Catherine Cooper, Jodie Hahn, and Mary P. Galea. 2016. "Morphology of Donor and Recipient Nerves Utilised in Nerve Transfers to Restore Upper Limb Function in Cervical Spinal Cord Injury" Brain Sciences 6, no. 4: 42. https://doi.org/10.3390/brainsci6040042