Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
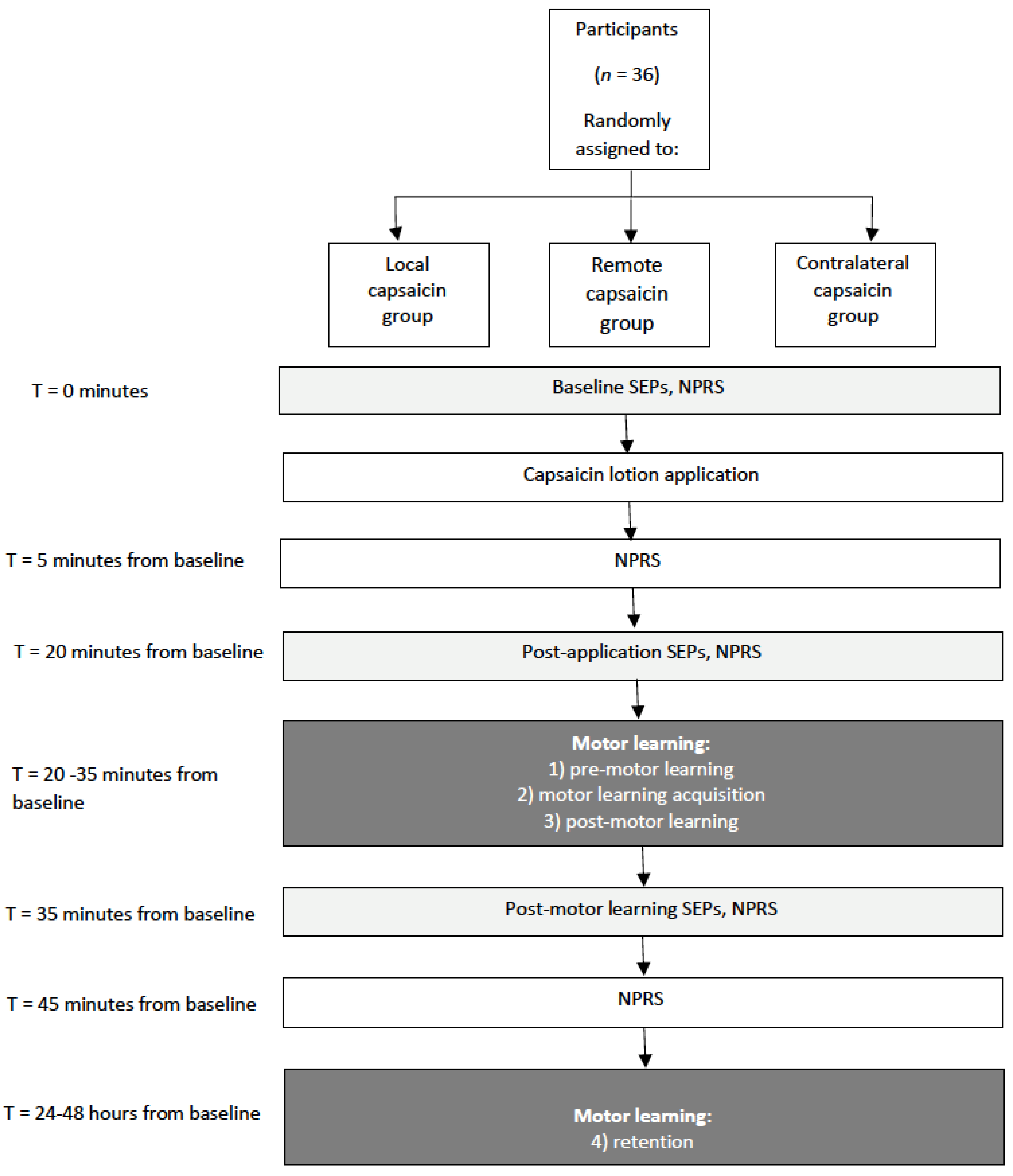
2. Materials and Methods
2.1. Methods Overview
2.2. Outcome Measures
2.3. Stimulation of Median Nerve to Elicit SEPs
2.4. SEP Recording Parameters
2.5. Motor Training Task
2.6. Pain
2.7. Data Analysis
3. Results
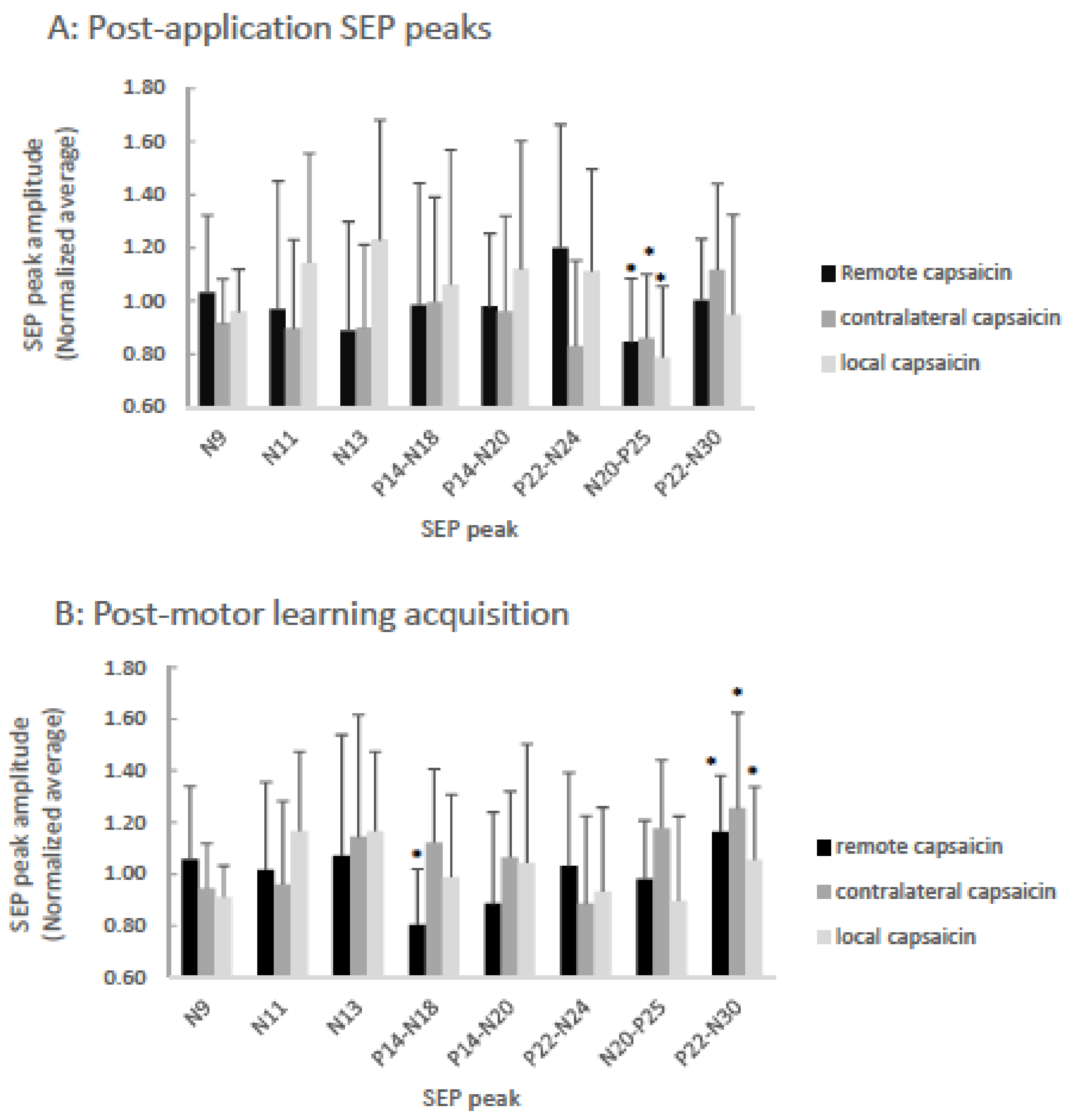
3.1. SEP Peaks
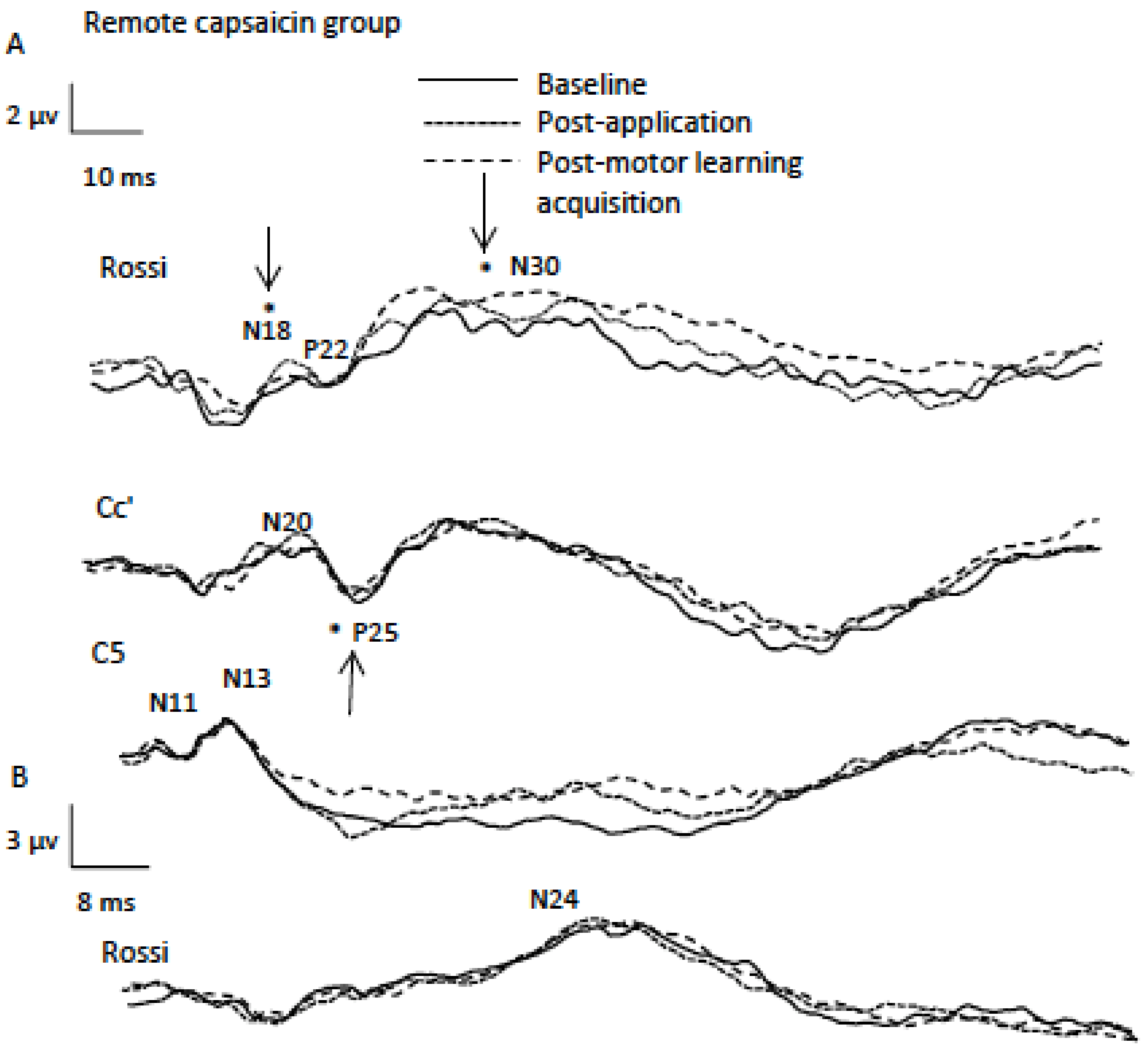
3.1.1. P25
3.1.2. N18
3.1.3. N30
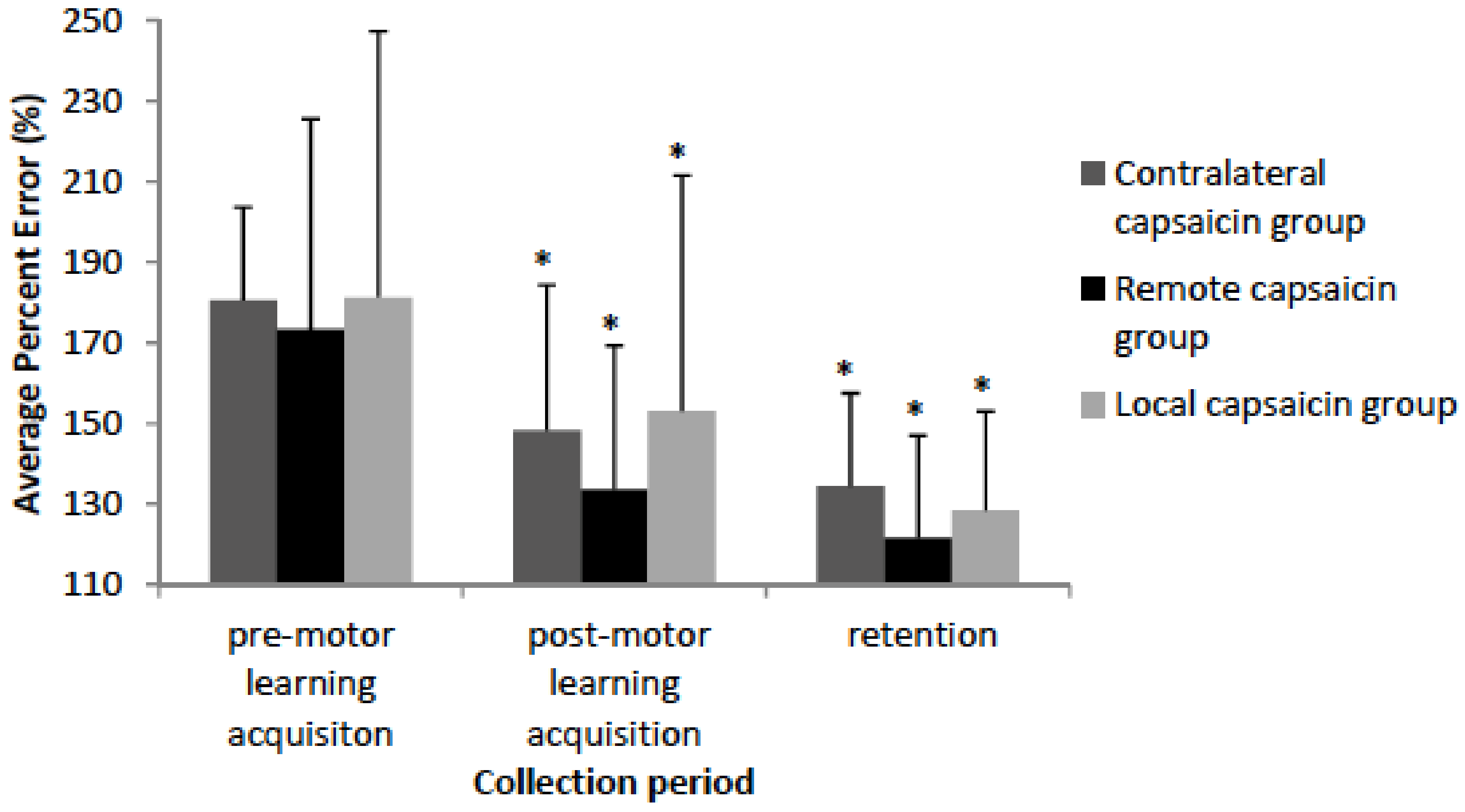
3.2. Motor Performance
3.2.1. Accuracy
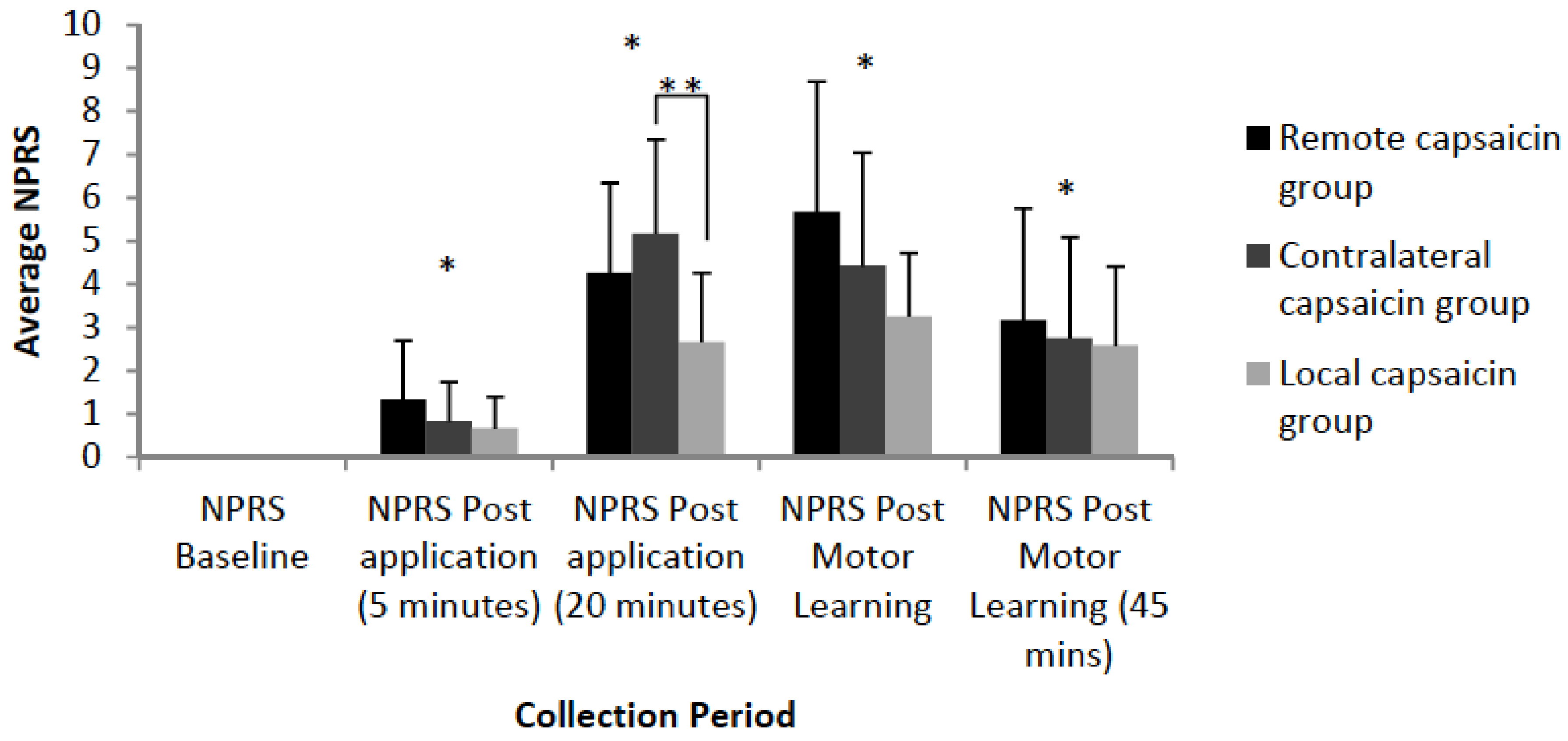
3.3. Pain Ratings
4. Discussion
4.1. SEP Peaks
4.1.1. Post Application SEP peaks: P25
4.1.2. Post-motor Learning Acquisition SEP Peaks: N18 and N30
4.2. Behavioural Data
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hodges, P.W.; Smeets, R.J. Interaction Between Pain, Movement, and Physical Activity: Short-term Benefits, Long-term Consequences, and Targets for Treatment. Clin. J. Pain 2015, 31, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, S.; Romaniello, A.; Wang, K.; Svensson, P.; Sessle, B.J.; Arendt-Nielsen, L. The effects of intra-oral pain on motor cortex neuroplasticity associated with short-term novel tongue-protrusion training in humans. Pain 2007, 132, 169. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.; Crown, E.; Grau, J. Nociceptive plasticity inhibits adaptive learning in the spinal cord. Neuroscience 2006, 141, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Hook, M.A.; Huie, J.R.; Grau, J.W. Peripheral inflammation undermines the plasticity of the isolated spinal cord. Behav. Neurosci. 2008, 122, 233. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, M.C.; Roosink, M.; Mercier, C. Effect of local versus remote tonic heat pain during training on acquisition and retention of a finger-tapping sequence task. Exp. Brain Res. 2016, 234, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Bouffard, J.; Bouyer, L.J.; Roy, J.-S.; Mercier, C. Tonic pain experienced during locomotor training impairs retention despite normal performance during acquisition. J. Neurosci. 2014, 34, 9190–9195. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The effect of local versus remote experimental pain on motor learning and sensorimotor integration using a complex typing task. Pain 2016, 157, 1682–1695. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.; Srbely, J.; Yielder, P. The effect of experimental pain on motor training performance and sensorimotor integration. Exp. Brain. Res. 2014, 232, 2879–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGaughy, J.; Dalley, J.; Morrison, C.; Everitt, B.; Robbins, T. Selective behavioral and neurochemical effects of cholinergic lesions produced by intrabasalis infusions of 192 IgG-saporin on attentional performance in a five-choice serial reaction time task. J. Neurosci. 2002, 22, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Culberson, A.; Packowski, C.; Chiba, A.A.; Tuszynski, M.H. Lesions of the basal forebrain cholinergic system impair task acquisition and abolish cortical plasticity associated with motor skill learning. Neuron 2003, 38, 819–829. [Google Scholar] [CrossRef]
- Rosenkranz, K.; Rothwell, J.C. The effect of sensory input and attention on the sensorimotor organization of the hand area of the human motor cortex. J. Physiol. 2004, 561, 307–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, K.; Wycislo, M.; Classen, J. Modulation of associative human motor cortical plasticity by attention. J. Neurophysiol. 2004, 92, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.D.; Gordon, N.C.; Smith, R.; Fields, H.L. Post-operative pain: Effect of extent of injury and attention. Brain Res. 1982, 234, 500–504. [Google Scholar] [CrossRef]
- Rode, S.; Salkovskis, P.M.; Jack, T. An experimental study of attention, labelling and memory in people suffering from chronic pain. Pain 2001, 94, 193–203. [Google Scholar] [CrossRef]
- McCaul, K.D.; Malott, J.M. Distraction and coping with pain. Psychol. Bull. 1984, 95, 516. [Google Scholar] [CrossRef] [PubMed]
- Miron, D.; Duncan, G.H.; Bushnell, M.C. Effects of attention on the intensity and unpleasantness of thermal pain. Pain 1989, 39, 345–352. [Google Scholar] [CrossRef]
- Bushnell, M.C.; Duncan, G.H.; Dubner, R.; Jones, R.L.; Maixner, W. Attentional influences on noxious and innocuous cutaneous heat detection in humans and monkeys. J. Neurosci. 1985, 5, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Magerl, W. Multiple mechanisms of secondary hyperalgesia. Prog. Brain Res. 2000, 129, 331–341. [Google Scholar] [PubMed]
- Koltzenburg, M.; Torebjörk, H.E.; Wahren, L.K. Nociceptor modulated central sensitization causes mechanical hyperalgesia in acute chemogenic and chronic neuropathic pain. Brain 1994, 117, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Maihöfner, C.; Handwerker, H.O. Differential coding of hyperalgesia in the human brain: A functional MRI study. Neuroimage 2005, 28, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Torebjörk, H.; Lundberg, L.; LaMotte, R. Central changes in processing of mechanoreceptive input in capsaicin-induced secondary hyperalgesia in humans. J. Physiol. 1992, 448, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Schmelz, M.; Forster, C.; Ringkamp, M.; Torebjork, E.; Handwerker, H. Novel classes of responsive and unresponsive C nociceptors in human skin. J. Neurosci. 1995, 15, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Shiri, R.; Viikari-Juntura, E.; Varonen, H.; Heliövaara, M. Prevalence and determinants of lateral and medial epicondylitis: A population study. Am. J. Epidemiol. 2006, 164, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Aminoff, M.; Curio, G.; Guerit, J.; Kakigi, R.; Mauguiere, F.; Rossini, P.; Treede, R.; Garcia-Larrea, L. Recommendations for the clinical use of somatosensory-evoked potentials. Clin. Neurophysiol. 2008, 119, 1705–1719. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; della Volpe, R.; Ginanneschi, F.; Ulivelli, M.; Bartalini, S.; Spidalieri, R.; Rossi, A. Early somatosensory processing during tonic muscle path in humans: Relation to loss of proprioception and motor defensive strategies. Clin. Neurophysiol. 2003, 114, 1351–1358. [Google Scholar] [CrossRef]
- Schabrun, S.M.; Jones, E.; Kloster, J.; Hodges, P.W. The temporal association between changes in primary sensory cortex and corticomotor output during muscle pain. Neurosci. 2013, 235, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The interactive effect of acute pain and motor learning acquisition on sensorimotor integration and motor learning outcomes. J. Neurophysiol. 2016, 116, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E. GPOWER: A Priori-, Post Hoc-, and Compromise Power Analyses for MS-DOS; Bonn University: Bonn, Germany, 1992. [Google Scholar]
- Mauguiere, F.; Allison, T.; Babiloni, C.; Buchner, H.; Eisen, A.; Goodin, D.; Jones, S.; Kakigi, R.; Matsuoka, S.; Nuwer, M. Somatosensory evoked potentials. The International Federation of Clinical Neurophysiology. Electroencephalogr. Clin. Neurophysiol. Suppl. 1999, 52, 79–90. [Google Scholar] [PubMed]
- Haavik, H.; Murphy, B. Selective changes in cerebellar-cortical processing following motor training. Exp. Brain Res. 2013, 231, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Yamada, T.; Aihara, M.; Kokubun, Y.; Noguchi, Y.; Matsubara, M.; Malcolm, H. The effects of stimulus rates upon median, ulnar and radial nerve somatosensory evoked potentials. Electroencephalogr. Clin. Neurophysiol. 1994, 92, 518–526. [Google Scholar] [CrossRef]
- Abbadie, C.; Trafton, J.; Liu, H.; Mantyh, P.W.; Basbaum, A.I. Inflammation increases the distribution of dorsal horn neurons that internalize the neurokinin-1 receptor in response to noxious and non-noxious stimulation. J. Neurosci. 1997, 17, 8049–8060. [Google Scholar] [CrossRef] [PubMed]
- Nuwer, M.R.; Aminoff, M.; Desmedt, J.; Eisen, A.A.; Goodin, D.; Matsuoka, S.; Mauguière, F.; Shibasaki, H.; Sutherling, W.; Vibert, J.F. IFCN recommended standards for short latency somatosensory evoked potentials. Report of an IFCN committee. International Federation of Clinical Neurophysiology. Electroencephalogr. Clin. Neurophysiol. 1994, 91, 6. [Google Scholar] [CrossRef]
- Holland, L.; Murphy, B.; Passmore, S.; Yielder, P. Time course of corticospinal excitability changes following a novel motor training task. Neurosci. Lett. 2015, 591, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Dolphin, N.W.; Crue, B.L., Jr. Pain: Clinical Manual for Nursing Practice. Clin. J. Pain 1989, 5, 363. [Google Scholar] [CrossRef]
- Streiner, D.L.; Norman, G.R. Biostatistics: The Bare Essentials, 3rd ed.; B.C. Decker Inc.: Hamilton, CA, USA, 2008. [Google Scholar]
- Jensen, M.P. A neuropsychological model of pain: Research and clinical implications. J. Pain 2010, 11, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Kaas, J.H.; Nelson, R.J.; Sur, M.; Lin, C.-S.; Merzenich, M.M. Multiple representations of the body within the primary somatosensory cortex of primates. Science 1979, 204, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Burton, H.; Fabri, M.; Alloway, K. Cortical areas within the lateral sulcus connected to cutaneous representations in areas 3B and 1: A revised interpretation of the second somatosensory area in macaque monkeys. J. Comp. Neurol. 1995, 355, 539–562. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.; Gigli, G.; Marciani, M.; Zarola, F.; Caramia, M. Non-invasive evaluation of input-output characteristics of sensorimotor cerebral areas in healthy humans. Electroencephalogr. Clin. Neurophysiol. 1987, 68, 88–100. [Google Scholar] [CrossRef]
- Molinari, M.; Restuccia, D.; Leggio, M.G. State estimation, response prediction, and cerebellar sensory processing for behavioral control. Cerebellum 2009, 8, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.H.; Parsons, L.M.; Bower, J.M.; Xiong, J.; Li, J.; Fox, P.T. Cerebellum implicated in sensory acquisition and discrimination rather than motor control. Science 1996, 272, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Jueptner, M.; Weiller, C. A review of differences between basal ganglia and cerebellar control of movements as revealed by functional imaging studies. Brain 1998, 121, 1437–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albus, J.S. A theory of cerebellar function. Math. Biosci. 1971, 10, 25–61. [Google Scholar] [CrossRef] [Green Version]
- Ito, M. Cerebellar long-term depression: Characterization, signal transduction, and functional roles. Physiol. Rev. 2001, 81, 1143–1195. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Bushnell, M.C.; Treede, R.D.; Zubieta, J.K. Human brain mechanisms of pain perception and regulation in health and disease. Eur. J. Pain 2005, 9, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Borsook, D. Pain and motor system plasticity. Pain 2007, 132, 8. [Google Scholar] [CrossRef] [PubMed]
- Legrain, V.; Van Damme, S.; Eccleston, C.; Davis, K.D.; Seminowicz, D.A.; Crombez, G. A neurocognitive model of attention to pain: Behavioral and neuroimaging evidence. Pain 2009, 144, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Desimone, R.; Duncan, J. Neural mechanisms of selective visual attention. Annu. Rev. Neurosci. 1995, 18, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Sonoo, M.; Sakuta, M.; Shimpo, T.; Genba, K.; Mannen, T. Widespread N18 in median nerve SEP is preserved in a pontine lesion. Electroencephalogr. Clin. Neurophysiol. 1991, 80, 238–240. [Google Scholar] [CrossRef]
- Noel, P.; Ozaki, I.; Desmedt, J. Origin of N18 and P14 far-fields of median nerve somatosensory evoked potentials studied in patients with a brain-stem lesion. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 167–170. [Google Scholar] [CrossRef]
- Society, A.C.N. Guideline 5: Guidelines for standard electrode position nomenclature. Am. J. Electroneurodiagn. Technol. 2006, 46, 222. [Google Scholar]
- Tamraz, J.C.; Comair, Y.G. The brainstem and cerebellum. In Atlas of Regional Anatomy of the Brain Using MRI: With Functional Correlations; Springer: Berlin, Germany, 2006; pp. 227–255. [Google Scholar]
- Moulton, E.; Schmahmann, J.; Becerra, L.; Borsook, D. The cerebellum and pain: Integrator or active participator? Brain Res. Rev. 2010, 65, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Glickstein, M.; May, J.G.; Mercier, B.E. Corticopontine projection in the macaque: The distribution of labelled cortical cells after large injections of horseradish peroxidase in the pontine nuclei. J. Comp. Neurol. 1985, 235, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Strick, P.L.; Dum, R.P.; Fiez, J.A. Cerebellum and nonmotor function. J. Comp. Neurol. 2009, 32, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Black, J.E.; Isaacs, K.R.; Anderson, B.J.; Alcantara, A.A.; Greenough, W.T. Learning causes synaptogenesis, whereas motor activity causes angiogenesis, in cerebellar cortex of adult rats. Proc. Natl. Acad. Sci. USA 1990, 87, 5568–5572. [Google Scholar] [CrossRef] [PubMed]
- Kleim, J.A. Synaptic Plasticity in the Cerebellum of the Adult Rat Following Complex Motor Skill Acquisition; University of Illinois at Urbana-Champaign: Champaign, IL, USA, 1994. [Google Scholar]
- Kleim, J.P.; Bender, R.; Kirsch, R.; Meichsner, C.; Paessens, A.; Rösner, M.; Rübsamen-Waigmann, H.; Kaiser, R.; Wichers, M.; Schneweis, K.E. Preclinical evaluation of HBY097, a new nonnucleoside reverse transcriptase inhibitor of human immunodeficiency virus type 1 replication. Antimicrob. Agents Chemother. 1995, 39, 2253–2257. [Google Scholar] [CrossRef] [PubMed]
- Classen, J.; Liepert, J.; Wise, S.P.; Hallett, M.; Cohen, L.G. Rapid plasticity of human cortical movement representation induced by practice. J. Neurosci. 1998, 79, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Helmchen, C.; Mohr, C.; Erdmann, C.; Petersen, D.; Nitschke, M. Differential cerebellar activation related to perceived pain intensity during noxious thermal stimulation in humans: A functional magnetic resonance imaging study. Neurosci. Lett. 2003, 335, 202–206. [Google Scholar] [CrossRef]
- Kanovsky, P.; Bares, M.; Rektor, I. The selective gating of the N30 cortical component of the somatosensory evoked potentials of median nerve is different in the mesial and dorsolateral frontal cortex: Evidence from intracerebral recordings. Clin. Neurophysiol. 2003, 114, 981–991. [Google Scholar] [CrossRef]
- Cebolla, A.; Palmero-Soler, E.; Dan, B.; Cheron, G. Frontal phasic and oscillatory generators of the N30 somatosensory evoked potential. Neuroimage 2011, 54, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Yielder, P.; Murphy, B. Do pursuit movement tasks lead to differential changes in early somatosensory evoked potentials related to motor learning compared with typing tasks? J. Neurophysiol. 2015, 113, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrew, D.; Yielder, P.; Murphy, B. Pursuit movement tasks lead to differential changes in early somatosensory evoked potentials related to motor learning in comparison to typing tasks. J. Neurophysiol. 2014. [Google Scholar] [CrossRef]
- Rossini, P.M.; Babiloni, F.; Babiloni, C.; Ambrosini, A.; Onorati, P.; Urbano, A. Topography of spatially enhanced human shortlatency somatosensory evoked potentials. Neuroreport 1997, 8, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Legon, W.; Dionne, J.K.; Meehan, S.K.; Staines, W.R. Non-dominant hand movement facilitates the frontal N30 somatosensory evoked potential. BMC Neurosci. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Kóbor, I.; Gál, V.; Vidnyánszky, Z. Attentional modulation of perceived pain intensity in capsaicin-induced secondary hyperalgesia. Exp. Brain. Res. 2009, 195, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Houlihan, M.E.; McGrath, P.J.; Connolly, J.F.; Stroink, G.; Finley, G.A.; Dick, B.; Phi, T.-T. Assessing the effect of pain on demands for attentional resources using ERPs. Int. J. Psychophysiol. 2004, 51, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Veldhuijzen, D.S.; Kenemans, J.L.; de Bruin, C.M.; Olivier, B.; Volkerts, E.R. Pain and attention: Attentional disruption or distraction? J. Pain 2006, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Eccleston, C. Chronic pain and distraction: An experimental investigation into the role of sustained and shifting attention in the processing of chronic persistent pain. Behav. Res. Ther. 1995, 33, 391–405. [Google Scholar] [CrossRef]
- Juottonen, K.; Gockel, M.; Silén, T.; Hurri, H.; Hari, R.; Forss, N. Altered central sensorimotor processing in patients with complex regional pain syndrome. Pain 2002, 98, 315–323. [Google Scholar] [CrossRef]
- Grigsby, J.; Rosenberg, N.L.; Busenbark, D. Chronic Pain is Associated with Deficits in Information Processing 1. Percept. Mot. Skills 1995, 81, 403–410. [Google Scholar] [CrossRef] [PubMed]
- McCracken, L.M.; Hoskins, J.; Eccleston, C. Concerns about medication and medication use in chronic pain. J. Pain 2006, 7, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.; Farmer, M.; Baliki, M.; Apkarian, A.V. Chronic pain: The role of learning and brain plasticity. Restor. Neurol. Neurosci. 2014, 32, 129–139. [Google Scholar] [PubMed]
- Robbins, T.W. Arousal systems and attentional processes. Biol. Psychol. 1997, 45, 57–71. [Google Scholar] [CrossRef]
- Deco, G.; Thiele, A. Attention–oscillations and neuropharmacology. Eur. J. Neurosci. 2009, 30, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Tracy, J.I.; Mohamed, F.; Faro, S.; Tiver, R.; Pinus, A.; Bloomer, C.; Pyrros, A.; Harvan, J. The effect of autonomic arousal on attentional focus. Neuroreport 2000, 11, 4037–4042. [Google Scholar] [CrossRef] [PubMed]
- Coull, J.T. Neural correlates of attention and arousal: Insights from electrophysiology, functional neuroimaging and psychopharmacology. Prog. Neurobiol. 1998, 55, 343–361. [Google Scholar] [CrossRef]
- Seminowicz, D.A.; Davis, K.D. Interactions of pain intensity and cognitive load: The brain stays on task. Cereb. Cortex 2007, 17, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Meyer, R.A.; Campbell, J.N. Myelinated mechanically insensitive afferents from monkey hairy skin: Heat-response properties. J. Neurophysiol. 1998, 80, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Cahill, L.; McGaugh, J.L. Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci. 1998, 21, 294–299. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dancey, E.; Yielder, P.; Murphy, B. Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration? Brain Sci. 2018, 8, 179. https://doi.org/10.3390/brainsci8100179
Dancey E, Yielder P, Murphy B. Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration? Brain Sciences. 2018; 8(10):179. https://doi.org/10.3390/brainsci8100179
Chicago/Turabian StyleDancey, Erin, Paul Yielder, and Bernadette Murphy. 2018. "Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration?" Brain Sciences 8, no. 10: 179. https://doi.org/10.3390/brainsci8100179
APA StyleDancey, E., Yielder, P., & Murphy, B. (2018). Does Location of Tonic Pain Differentially Impact Motor Learning and Sensorimotor Integration? Brain Sciences, 8(10), 179. https://doi.org/10.3390/brainsci8100179