A Review of Functional and Structural Neurobiology of the Action Observation Network in Autism Spectrum Disorder and Developmental Coordination Disorder


Abstract
:1. Introduction
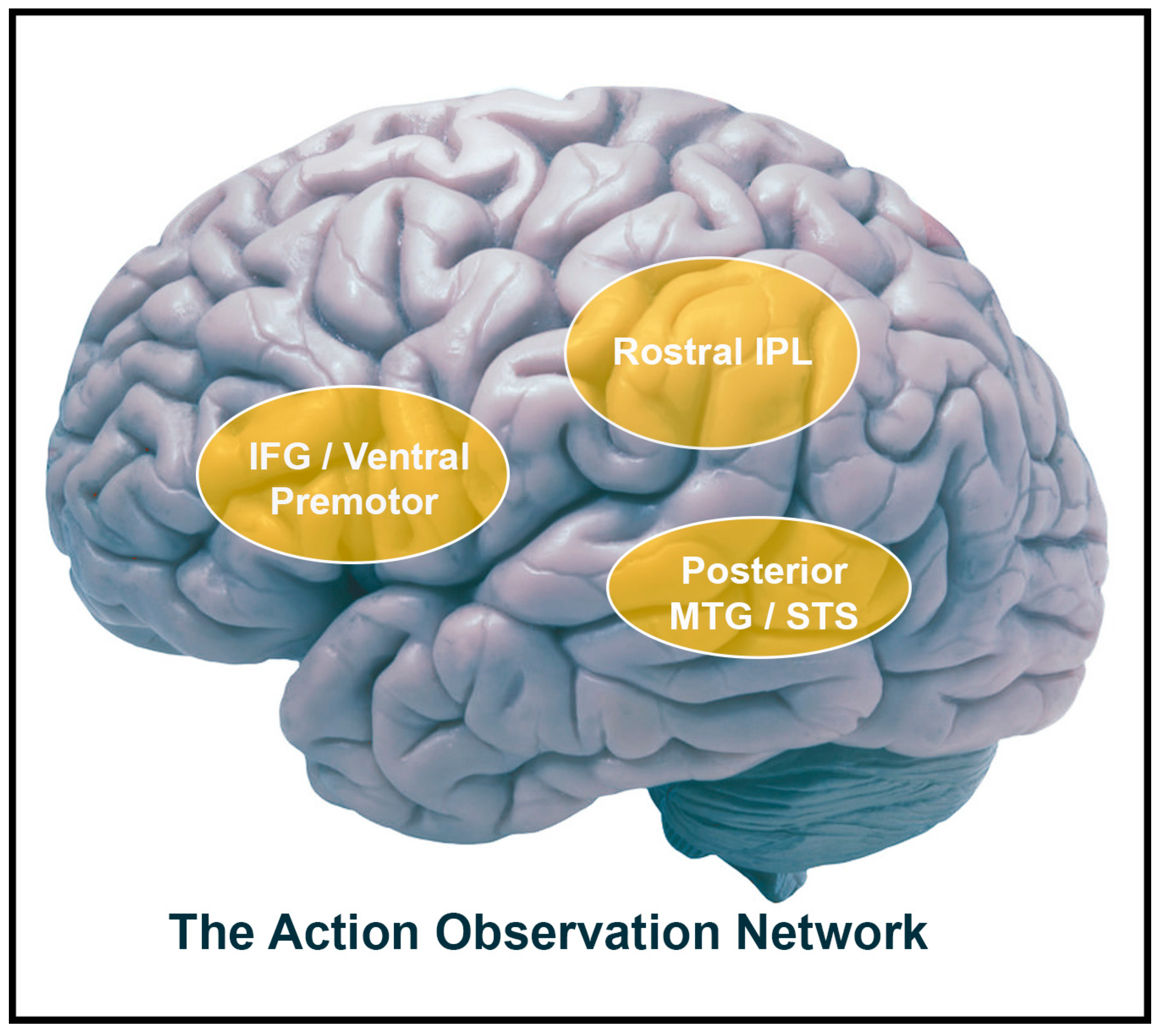
1.1. The Action Observation Network (AON)
1.2. Developmental Coordination Disorder (DCD) and Imitation Impairments
1.3. Motor and Imitation Impairments in ASD
1.4. Imitation Impairments in ASD and DCD
2. Task-Based AON Research
2.1. AON Activity in ASD
Neural Theories of Aberrant Findings in ASD
2.2. AON Activity in DCD
2.3. AON Activity in ASD and DCD
AON Connectivity in ASD and DCD During Tasks and at Rest
3. Resting State Research
3.1. ASD AON Task and Resting State Connectivity
3.2. DCD Task Based and Resting State Connectivity
3.3. ASD and DCD AON Resting State and Grey Matter Differences
4. AON White Matter Organization
4.1. ASD AON White Matter Organization
4.2. DCD AON White Matter Organization
4.3. AON White Matter Organization in DCD and ASD
5. Conclusions and Future Directions
6. Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; DSM-5; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 9780890425558. [Google Scholar]
- Hilton, C.L.; Zhang, Y.; Whilte, M.R.; Klohr, C.L.; Constantino, J.N. Motor Impairment in Sibling Pairs Concordant and Discordant for Autism Spectrum Disorders. Autism 2012, 16, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.N.; Landa, R.J.; Galloway, J.C. (Cole) Current Perspectives on Motor Functioning in Infants, Children, and Adults with Autism Spectrum Disorders. Phys. Ther. 2011, 91, 1116–1129. [Google Scholar] [CrossRef] [PubMed]
- Holloway, J.M.; Long, T.M.; Biasini, F. Relationships between Gross Motor Skills and Social Function in Young Boys with Autism Spectrum Disorder. Pediatr. Phys. Ther. 2018, 30, 1. [Google Scholar] [CrossRef] [PubMed]
- Kanner, L. Autistic Disturbances of Affective Contact. Nerv. Child 1943, 2. [Google Scholar] [CrossRef]
- Sinani, C.; Sugden, D.A.; Hill, E.L. Gesture Production in School vs. Clinical Samples of Children with Developmental Coordination Disorder (DCD) and Typically Developing Children. Res. Dev. Disabil. 2011, 32, 1270–1282. [Google Scholar] [CrossRef]
- Ahonen, T.P. Abstract: Developmental Coordination Disorders in Children: A Developmental Neuropsychological Follow-Up Study. Jyraskyla Stud. Educ. Psychol. Soc. Res. 1990, 78, 188. [Google Scholar]
- Cantell, M.H. Developmental Coordination Disorder in Adolescence: Perceptual-Motor, Academic and Social Outcomes Of Early Motor Delay. Ph.D. Dissertation, University of Lancaster, Lancaster, UK, 1998. [Google Scholar]
- Cantell, M.H.; Smyth, M.M.; Ahonen, T.P. Clumsiness in Adolescence: Educational, Motor, and Social Outcomes of Motor Delay Detected at 5 Years. Adapt. Phys. Act. Q. 1994, 11, 115–129. [Google Scholar] [CrossRef]
- Dewey, D.; Cantell, M.H.; Crawford, S.G. Motor and Gestural Performance in Children with Autism Spectrum Disorders, Developmental Coordination Disorder, and/or Attention Deficit Hyperactivity Disorder. J. Int. Neuropsychol. Soc. 2007, 13, 246–256. [Google Scholar] [CrossRef]
- Dewey, D.; Kaplan, B.J.; Crawford, S.G.; Wilson, B.N. Developmental Coordination Disorder: Associated Problems in Attention, Learning, and Psychosocial Adjustment. Hum. Mov. Sci. 2002, 21, 905–918. [Google Scholar] [CrossRef]
- Missiuna, C.; Gaines, R.; Mclean, J.; Delaat, D.; Egan, M.; Soucie, H. Description of Children Identified by Physicians as Having Developmental Coordination Disorder. Dev. Med. Child Neurol. 2008, 50, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Alaerts, K.; Woolley, D.G.; Steyaert, J.; Di Martino, A.; Swinnen, S.P.; Wenderoth, N. Underconnectivity of the Superior Temporal Sulcus Predicts Emotion Recognition Deficits in Autism. Soc. Cogn. Affect. Neurosci. 2013, 9, 1589–1600. [Google Scholar] [CrossRef]
- Dapretto, M.; Davies, M.S.; Pfeifer, J.H.; Scott, A.A.; Sigman, M.; Bookheimer, S.Y.; Iacoboni, M. Understanding Emotions in Others: Mirror Neuron Dysfunction in Children with Autism Spectrum Disorders. Nat. Neurosci. 2006, 9, 28–30. [Google Scholar] [CrossRef]
- Oberman, L.M.; Hubbard, E.M.; McCleery, J.P.; Altschuler, E.L.; Ramachandran, V.S.; Pineda, J.A. EEG Evidence for Mirror Neuron Dysfunction in Autism Spectrum Disorders. Cogn. Brain Res. 2005, 24, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Delbruck, E.; Yang, M.; Yassine, A.; Grossman, E.D. Functional Connectivity in ASD: Atypical Pathways in Brain Networks Supporting Action Observation and Joint Attention. Brain Res. 2019, 1706, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zwicker, J.G.; Missiuna, C.; Harris, S.R.; Boyd, L.A. Brain Activation of Children with Developmental Coordination Disorder Is Different Than Peers. Pediatrics 2010, 126, e678–e686. [Google Scholar] [CrossRef] [PubMed]
- Caspers, S.; Zilles, K.; Laird, A.R.; Eickhoff, S.B. ALE Meta-Analysis of Action Observation and Imitation in the Human Brain. Neuroimage 2010, 50, 1148–1167. [Google Scholar] [CrossRef]
- Cross, E.S.; Hamilton, A.; Kraemer, D.J.M.; Kelley, W.M.; Grafton, S.T. Dissociable Substrates for Body Motion and Physical Experience in the Human Action Observation Network. Eur. J. 2009, 30, 1383–1392. [Google Scholar] [CrossRef]
- Gazzola, V.; Rizzolatti, G.; Wicker, B.; Keysers, C. the Anthropomorphic Brain: The Mirror Neuron System Responds to Human and Robotic Actions. Neuroimage 2007, 35, 1674–1684. [Google Scholar] [CrossRef]
- Iacoboni, M.; Mazziotta, J.C. Mirror Neuron System: Basic Findings and Clinical Applications. Ann. Neurol. 2007, 62, 213–218. [Google Scholar] [CrossRef]
- Chong, T.J.; Cunnington, R.; Williams, M.A.; Kanwisher, N.; Mattingley, J.B. Report FMRI Adaptation Reveals Mirror Neurons in Human Inferior Parietal Cortex. Curr. Biol. 2008, 18, 1576–1580. [Google Scholar] [CrossRef]
- Rizzolatti, G. the Mirror Neuron System and Its Function in Humans. Anat. Embryol. 2005, 210, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef]
- Biagi, L.; Cioni, G.; Fogassi, L.; Guzzetta, A.; Sgandurra, G.; Tosetti, M. Action Observation Network in Childhood: A Comparative FMRI Study with Adults. Dev. Sci. 2016, 19, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Quandt, L.C.; Chatterjee, A. Rethinking Actions: Implementation and Association. Wiley Interdiscip. Rev. Cogn. Sci. 2015, 6, 483–490. [Google Scholar] [CrossRef]
- Gazzola, V.; Aziz-Zadeh, L.; Keysers, C. Empathy and the Somatotopic Auditory Mirror System in Humans. Curr. Biol. 2006, 16, 1824–1829. [Google Scholar] [CrossRef]
- Ortigue, S.; Sinigaglia, C.; Rizzolatti, G.; Grafton, S.T. Understanding Actions of Others: The Electrodynamics of the Left and Right Hemispheres. A High-Density EEG Neuroimaging Study. PLoS ONE 2010, 5, e12160. [Google Scholar] [CrossRef]
- Kaplan, J.T.; Iacoboni, M. Getting a Grip on Other Minds: Mirror Neurons, Intention Understanding, and Cognitive Empathy. Soc. Neurosci. 2006, 1, 175–183. [Google Scholar] [CrossRef]
- Sobhani, M.; Fox, G.R.; Kaplan, J.T.; Aziz-Zadeh, L. Interpersonal Liking Modulates Motor-Related Neural Regions. PLoS ONE 2012, 7, e46809. [Google Scholar] [CrossRef]
- Fishman, I.; Datko, M.; Cabrera, Y.; Carper, R.A.; Müller, R.A. Reduced Integration and Differentiation of the Imitation Network in Autism: A Combined Functional Connectivity Magnetic Resonance Imaging and Diffusion-Weighted Imaging Study. Ann. Neurol. 2015, 78, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Kashuk, S.R.; Wilson, P.H.; Thorpe, G.; Egan, G.F. White Matter Alterations in Adults with Probable Developmental Coordination Disorder: An MRI Diffusion Tensor Imaging Study. Neuroreport 2017, 28, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Bastiaansen, J.A.; Thioux, M.; Nanetti, L.; Van Der Gaag, C.; Ketelaars, C.; Minderaa, R.; Keysers, C. Age-Related Increase in Inferior Frontal Gyrus Activity and Social Functioning in Autism Spectrum Disorder. Biol. Psychiatry 2011, 69, 832–838. [Google Scholar] [CrossRef]
- Oberman, L.M.; Ramachandran, V.S. the simulating social mind: The role of the mirror neuron system and simulation in the social and communicative deficits of autism spectrum disorders. Psychol. Bull. 2007, 133, 310–327. [Google Scholar] [CrossRef]
- Iacoboni, M.; Dapretto, M. The Mirror Neuron System and the Consequences of Its Dysfunction. Nat. Rev. Neurosci. 2006, 7, 942–951. [Google Scholar] [CrossRef]
- Steinhorst, A.; Funke, J. Mirror Neuron Activity Is No Proof for Action Understanding. Front. Hum. Neurosci. 2014, 8, 333. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.; Bird, G.; Catmur, C.; Press, C.; Heyes, C. Mirror Neurons: From Origin to Function. Behav. Brain Sci. 2014, 37, 177–192. [Google Scholar] [CrossRef]
- Hickok, G. Eight Problems for the Mirror Neuron Theory of Action Understanding in Monkeys and Humans. J. Cogn. Neurosci. 2009, 21, 1229–1243. [Google Scholar] [CrossRef] [PubMed]
- Schultz, W. Multiple Reward Signals in the Brain. Nat. Rev. Neurosci. 2000, 1, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.G.; Whiten, A.; Suddendorf, T.; Perrett, D.I. Imitation, Mirror Neurons and Autism. Neurosci. Biobehav. Rev. 2001, 25, 287–295. [Google Scholar] [CrossRef]
- Schulte-Rüther, M.; Otte, E.; Adigüzel, K.; Firk, C.; Herpertz-Dahlmann, B.; Koch, I.; Konrad, K. Intact Mirror Mechanisms for Automatic Facial Emotions in Children and Adolescents with Autism Spectrum Disorder. Autism Res. 2017, 10, 298–310. [Google Scholar] [CrossRef]
- Alaerts, K.; Geerlings, F.; Herremans, L.; Swinnen, S.P.; Verhoeven, J.; Sunaert, S.; Wenderoth, N. Functional Organization of the Action Observation Network in Autism: A Graph Theory Approach. PLoS ONE 2015, 10, e0137020. [Google Scholar] [CrossRef] [PubMed]
- Dinstein, I.; Thomas, C.; Humphreys, K.; Minshew, N.J.; Behrmann, M.; Heeger, D.J. Normal Movement Selectivity in Autism. Neuron 2010, 66, 461–469. [Google Scholar] [CrossRef]
- Marsh, L.E.; Hamilton, A. Dissociation of Mirroring and Mentalising Systems in Autism. Neuroimage 2011, 56, 1511–1519. [Google Scholar] [CrossRef]
- Craig, F.; Lorenzo, A.; Lucarelli, E.; Russo, L.; Fanizza, I.; Trabacca, A. Motor competency and social communication skills in preschool children with autism spectrum disorder. Autism Res. 2018, 11, 893–902. [Google Scholar] [CrossRef]
- Leary, M.R. Moving on: Autism and Movement Disturbance. Ment. Retard. 1996, 34, 39–53. [Google Scholar]
- Cairney, J. Developmental Coordination Disorder and Its Consequences; University of Toronto Press: Toronto, ON, Canada, 2015. [Google Scholar]
- Sumner, E.; Leonard, H.C.; Hill, E.L. Overlapping Phenotypes in Autism Spectrum Disorder and Developmental Coordination Disorder: A Cross-Syndrome Comparison of Motor and Social Skills. J. Autism Dev. Disord. 2016, 46, 2609–2620. [Google Scholar] [CrossRef]
- Gomez, A.; Sirigu, A. Developmental Coordination Disorder: Core Sensori-Motor Deficits, Neurobiology and Etiology. Neuropsychologia 2015, 79, 272–287. [Google Scholar] [CrossRef]
- Piek, J.P.; Dyck, M.J. Sensory-Motor Deficits in Children with Developmental Coordination Disorder, Attention Deficit Hyperactivity Disorder and Autistic Disorder. Hum. Mov. Sci. 2004, 23, 475–488. [Google Scholar] [CrossRef]
- Zoia, S.; Pelamatti, G.; Cuttini, M.; Casotto, V.; Scabar, A. Performance of Gesture in Children with and Without DCD: Effects of Sensory Input Modalities. Dev. Med. Child Neurol. 2002, 44, 699–705. [Google Scholar] [CrossRef]
- Reynolds, J.E.; Kerrigan, S.; Elliott, C.; Lay, B.S.; Licari, M.K. Poor Imitative Performance of Unlearned Gestures in Children with Probable Developmental Coordination Disorder. J. Mot. Behav. 2017, 49, 378–387. [Google Scholar] [CrossRef]
- Dewey, D. Error Analysis of Limb and Orofacial Praxis in Children with Developmental Motor Deficits. Brain Cogn. 1993, 23, 203–221. [Google Scholar] [CrossRef]
- Dewey, D.; Kaplan, B.J. Analysis of Praxis Task Demands in the Assessment of Children with Developmental Motor Deficits. Dev. Neuropsychol. 1992, 8, 367–379. [Google Scholar] [CrossRef]
- Filipčič, T.; Ozbič, M. Prediction of Learning Difficulties with the Test of Complex Imitation of Movement. Acta Univ. Palacki. Olomuc. Gymnica 2008, 38, 25–30. [Google Scholar]
- Wilson, P.H.; Adams, I.L.J.; Caeyenberghs, K.; Thomas, P.; Smits-Engelsman, B.; Steenbergen, B. Motor Imagery Training Enhances Motor Skill in Children with DCD: A Replication Study. Res. Dev. Disabil. 2016, 57, 54–62. [Google Scholar] [CrossRef]
- Barhoun, P.; Fuelscher, I.; Kothe, E.J.; He, J.L.; Youssef, G.J.; Enticott, P.G.; Williams, J.; Hyde, C. Motor Imagery in Children with DCD: A Systematic and Meta-Analytic Review of Hand-Rotation Task Performance. Neurosci. Biobehav. Rev. 2019, 99, 282–297. [Google Scholar] [CrossRef]
- Ayres, A.J. Sensory Integration and Praxis Tests (SIPT); École des Sciences de la réadaptation, Sciences de la santé, Université d. Ottawa: Ottawa, ON, Canada, 1988. [Google Scholar]
- Elbasan, B.; Kayıhan, H.; Duzgun, I. Sensory Integration and Activities of Daily Living in Children with Developmental Coordination Disorder. Ital. J. Pediatr. 2012, 38, 14. [Google Scholar] [CrossRef]
- Ozbič, M.; Filipčič, T. Complex Imitation of Gestures in School-Aged Children with Learning Difficulties. Kinesiology 2010, 42, 44–55. [Google Scholar]
- Goyen, T.-A.; Lui, K.; Hummell, J. Sensorimotor Skills Associated with Motor Dysfunction in Children Born Extremely Preterm. Early Hum. Dev. 2011, 87, 489–493. [Google Scholar] [CrossRef]
- Rothi, L.J.G.; Raymer, A.M.; Heilman, K.M. Limb Praxis Assessment. In Apraxia: The Neuropsychology of Action; Psychology Press: London, UK, 1997; pp. 61–73. [Google Scholar]
- Dowell, L.; Mahone, E.M.; Mostofsky, S.H. Associations of Postural Knowledge and Basic Motor Skill with Dyspraxia in Autism: Implication for Abnormalities in Distributed Connectivity and Motor Learning. Neuropsychology 2009, 23, 563–570. [Google Scholar] [CrossRef]
- Mostofsky, S.H.; Dubey, P.; Jerath, V.K.; Jansiewicz, E.M.; Goldburg, M.C.; Denckla, M.B. Developmental Dyspraxia Is Not Limited to Imitation in Children with Autism Spectrum Disorders. J. Int. Neuropsychol. Soc. 2006, 12, 314–326. [Google Scholar] [CrossRef]
- Visser, J. Developmental Coordination Disorder: A Review of Research On Subtypes and Comorbidities. Hum. Mov. Sci. 2003, 22, 479–493. [Google Scholar] [CrossRef]
- Dewey, D.; Kaplan, B.J. Subtyping of Developmental Motor Deficits. Dev. Neuropsychol. 1994, 10, 265–284. [Google Scholar] [CrossRef]
- Green, D.; Chambers, M.E.; Sugden, D.A. Does Subtype of Developmental Coordination Disorder Count: Is There a Differential Effect on Outcome Following Intervention? Hum. Mov. Sci. 2008, 27, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Macnab, J.J.; Miller, L.T.; Polatajko, H.J. The Search for Subtypes of DCD: Is Cluster Analysis the Answer? Hum. Mov. Sci. 2001, 20, 49–72. [Google Scholar] [CrossRef]
- Wright, H.C.; Sugden, D.A. the Nature of Developmental Coordination Disorder: Inter- and Intragroup Differences. Adapt. Phys. Act. Q. 1996, 13, 357–371. [Google Scholar] [CrossRef]
- Hoare, D. Subtypes of Developmental Coordination Disorder. Adapt. Phys. Act. Q. 1994, 11, 158–169. [Google Scholar] [CrossRef]
- Koutsouki, D.; Asonitou, K. Cognitive Processes in Children with Developmental Coordination Disorder. In Cognition, Intelligence, and Achievement; Elsevier: Amsterdam, The Netherlands, 2015; pp. 267–289. [Google Scholar]
- Vaivre-Douret, L.; Lalanne, C.; Ingster-Moati, I.; Boddaert, N.; Cabrol, D.; Dufier, J.-L.; Golse, B.; Falissard, B. Subtypes of Developmental Coordination Disorder: Research on Their Nature and Etiology. Dev. Neuropsychol. 2011, 36, 614–643. [Google Scholar] [CrossRef] [PubMed]
- Lalanne, C.; Falissard, B.; Golse, B.; Vaivre-Douret, L. Refining Developmental Coordination Disorder Subtyping with Multivariate Statistical Methods. BMC Med. Res. Methodol. 2012, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Asperger, H. Die “Autistischen Psychopathen” Im Kindesalter. Arch. Psychiatr. Nervenkr. 1944, 117, 76–136. [Google Scholar] [CrossRef]
- Asperger, H. Problems of Infantile Autism. Communication 1979, 13, 45–52. [Google Scholar]
- Kanner, L.; Lesser, L.I. Early Infantile Autism. Pediatr. Clin. N. Am. 1958, 5, 711–730. [Google Scholar] [CrossRef]
- Dziuk, M.A.; Larson, J.C.G.; Apostu, A.; Mahone, E.M.; Denckla, M.B.; Mostofsky, S.H. Dyspraxia in Autism: Association with Motor, Social, and Communicative Deficits. Dev. Med. Child Neurol. 2007, 49, 734–739. [Google Scholar] [CrossRef]
- Mosconi, M.W.; Sweeney, J.A. Sensorimotor Dysfunctions As Primary Features of Autism Spectrum Disorders. Sci. China Life Sci. 2015, 58, 1016–1023. [Google Scholar] [CrossRef]
- Mostofsky, S.; Goldberg, M.; Landa, R.; Denckla, M. Evidence for A Deficit in Procedural Learning in Children and Adolescents with Autism: Implications for Cerebellar Contribution. J. Int. Neuropsychol. Soc. 2000, 6. [Google Scholar] [CrossRef]
- Takarae, Y.; Minshew, N.J.; Luna, B.; Sweeney, J.A. Oculomotor Abnormalities Parallel Cerebellar Histopathology in Autism. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1359–1361. [Google Scholar] [CrossRef]
- Tripi, G.; Roux, S.; Carotenuto, M.; Bonnet-Brilhault, F.; Roccella, M. Minor Neurological Dysfunctions (MNDs) in Autistic Children without Intellectual Disability. J. Clin. Med. 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Conson, M.; Mazzarella, E.; Frolli, A.; Esposito, D.; Marino, N.; Trojano, L.; Grossi, D. Motor imagery in Asperger syndrome: testing action simulation by the hand laterality task. PLoS ONE 2013, 8, e70734. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, N.; Charman, T. the Prodrome of Autism: Early Behavioral and Biological Signs, Regression, Peri- and Post-Natal Development and Genetics. J. Child Psychol. Psychiatry 2010, 51, 432–458. [Google Scholar] [CrossRef] [PubMed]
- Roley, S.S.; Mailloux, Z.; Parham, L.D.; Schaaf, R.C.; Lane, C.J.; Cermak, S.A. Sensory Integration and Praxis Patterns in Children with Autism. Am. J. Occup. Ther. 2015, 69, 6901220010p1. [Google Scholar] [CrossRef]
- Setoh, P.; Marschik, P.B.; Einspieler, C.; Esposito, G. Autism Spectrum Disorder and Early Motor Abnormalities: Connected or Coincidental Companions? Res. Dev. Disabil. 2017, 60, 13–15. [Google Scholar] [CrossRef]
- Harris, A.; Williams, J.M. the Impact of A Horse Riding Intervention on the Social Functioning of Children with Autism Spectrum Disorder. Int. J. Environ. Res. Public Health 2017, 14, 776. [Google Scholar] [CrossRef]
- Paquet, A.; Olliac, B.; Bouvard, M.-P.; Golse, B.; Vaivre-Douret, L. the Semiology of Motor Disorders in Autism Spectrum Disorders as Highlighted from a Standardized Neuro-Psychomotor Assessment. Front. Psychol. 2016, 7, 1292. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.H.; Smits-Engelsman, B.; Caeyenberghs, K.; Steenbergen, B.; Sugden, D.A.; Clark, J.E.; Mumford, N.; Blank, R. Cognitive and Neuroimaging Findings in Developmental Coordination Disorder: New Insights from a Systematic Review of Recent Research. Dev. Med. Child Neurol. 2017, 59, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Fournier, K.A.; Hass, C.J.; Naik, S.K.; Lodha, N.; Cauraugh, J.H. Motor Coordination in Autism Spectrum Disorders: A Synthesis and Meta-Analysis. J. Autism Dev. Disord. 2010, 40, 1227–1240. [Google Scholar] [CrossRef]
- Ming, X.; Brimacombe, M.; Wagner, G.C. Prevalence of Motor Impairment in Autism Spectrum Disorders. Brain Dev. 2007, 29, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Minshew, N.J.; Sung, K.; Jones, B.L.; Furman, J.M. Underdevelopment of the Postural Control System in Autism. Neurology 2004, 63, 2056–2061. [Google Scholar] [CrossRef] [PubMed]
- Van Waelvelde, H.; Oostra, A.; Dewitte, G.; Van Den Broeck, C.; Jongmans, M. Stability of Motor Problems in Young Children with or at Risk of Autism Spectrum Disorders, ADHD, and or Developmental Coordination Disorder. Dev. Med. Child Neurol. 2010, 52, e174–e178. [Google Scholar] [CrossRef]
- Wilson, R.B.; McCracken, J.T.; Rinehart, N.J.; Jeste, S.S. What’s Missing in Autism Spectrum Disorder Motor Assessments? J. Neurodev. Disord. 2018, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Theoret, H.; Halligan, E.; Kobayashi, M.; Fregni, F.; Tager-Flusberg, H.; Pascual-Leone, A. Supplemental Data: Impaired Motor Facilitation During Action Observation in Individuals with Autism Spectrum Disorder. Curr. Biol. 2005, 15, R1–R2. [Google Scholar] [CrossRef] [PubMed]
- Enticott, P.G.; Kennedy, H.A.; Rinehart, N.J.; Tonge, B.J.; Bradshaw, J.L.; Taffe, J.R.; Daskalakis, Z.J.; Fitzgerald, P.B. Mirror Neuron Activity Associated with Social Impairments but Not Age in Autism Spectrum Disorder. Biol. Psychiatry 2012, 71, 427–433. [Google Scholar] [CrossRef]
- Williams, J.H.G.; Whiten, A.; Singh, T. A systematic review of action imitation in autistic spectrum disorder. J. Autism Dev. Disord. 2004, 34, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Brindley, R.; Frith, U. Imitation and Action Understanding in Autistic Spectrum Disorders: How Valid Is the Hypothesis of A Deficit in the Mirror Neuron System? Neuropsychologia 2007, 45, 1859–1868. [Google Scholar] [CrossRef]
- Press, C.; Richardson, D.; Bird, G. Intact Imitation of Emotional Facial Actions in Autism Spectrum Conditions. Neuropsychologia 2010, 48, 3291–3297. [Google Scholar] [CrossRef] [PubMed]
- Schunke, O.; Schöttle, D.; Vettorazzi, E.; Brandt, V.; Kahl, U.; Bäumer, T.; Ganos, C.; David, N.; Peiker, I.; Engel, A.K.; et al. Mirror Me: Imitative Responses in Adults with Autism. Autism 2016, 20, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Bird, G.; Leighton, J.; Press, C.; Heyes, C. Intact Automatic Imitation of Human and Robot Actions in Autism Spectrum Disorders. Proc. R. Soc. B Biol. Sci. 2007, 274, 3027–3031. [Google Scholar] [CrossRef]
- Spengler, S.; Bird, G.; Brass, M. Hyperimitation of Actions Is Related to Reduced Understanding of Others’ Minds in Autism Spectrum Conditions. Biol. Psychiatry 2010, 68, 1148–1155. [Google Scholar] [CrossRef]
- Ingersoll, B. The Social Role of Imitation in Autism: Implications for the Treatment of Imitation Deficits. Infants Young Child. 2008, 21, 107–119. [Google Scholar] [CrossRef]
- Hamilton, A. Emulation and mimicry for social interaction: A theoretical approach to imitation in autism. Proc. Q. J. Exp. Psychol. 2008, 61, 101–115. [Google Scholar] [CrossRef]
- Kana, R.K.; Wadsworth, H.M.; Travers, B.G. A systems level analysis of the mirror neuron hypothesis and imitation impairments in autism spectrum disorders. Neurosci. Biobehav. Rev. 2011, 35, 894–902. [Google Scholar] [CrossRef]
- Rogers, S.J.; Bennetto, L.; McEvoy, R.; Pennington, B.F. Imitation and Pantomime in High-Functioning Adolescents with Autism Spectrum Disorders. Child Dev. 1996, 67, 2060–2073. [Google Scholar] [CrossRef]
- Bryson, S.E.; Smith, I.M. Epidemiology of autism: Prevalence, associated characteristics, and implications for research and service delivery. Ment. Retard. Dev. Disabil. Res. Rev. 1998, 4, 97–103. [Google Scholar] [CrossRef]
- Aldridge, M.A.; Stone, K.R.; Sweeney, M.H.; Bower, T.G.R. Preverbal Children with Autism Understand the Intentions of Others. Dev. Sci. 2000, 3, 294–301. [Google Scholar] [CrossRef]
- Meltzoff, A.N. Origins of Theory of Mind, Cognition and Communication. J. Commun. Disord. 1999, 32, 251–269. [Google Scholar] [CrossRef]
- Paquet, A.; Olliac, B.; Golse, B.; Vaivre-Douret, L. Nature of Motor Impairments in Autism Spectrum Disorder: A Comparison with Developmental Coordination Disorder. J. Clin. Exp. Neuropsychol. 2019, 41, 1–14. [Google Scholar] [CrossRef]
- Green, D.; Baird, G.; Barnett, A.L.; Henderson, L.; Huber, J.; Henderson, S.E. The Severity and Nature of Motor Impairment in Asperger’s Syndrome: A Comparison with Specific Developmental Disorder of Motor Function. J. Child Psychol. Psychiatry 2002, 43, 655–668. [Google Scholar] [CrossRef]
- Cermak, S.A.; Quintero, E.J.; Cohen, P.M. Developmental Age Trends in Crossing the Body Midline in Normal Children. Am. J. Occup. Ther. 1980, 34, 313–319. [Google Scholar] [CrossRef]
- Henderson, S.; Sugden, D. Movement Assessment Battery for Children; The Psychological Corporation, Harcourt Brace Jovanovich: London, UK, 1992. [Google Scholar]
- Dane, S.; Balci, N. Handedness, Eyedness and Nasal Cycle in Children with Autism. Int. J. Dev. Neurosci. 2007, 25, 223–226. [Google Scholar] [CrossRef]
- Hill, E.L. Executive Dysfunction in Autism. Trends Cogn. Sci. 2004, 8, 26–32. [Google Scholar] [CrossRef]
- Schmitt, L.M.; Cook, E.H.; Sweeney, J.A.; Mosconi, M.W. Saccadic Eye Movement Abnormalities in Autism Spectrum Disorder Indicate Dysfunctions in Cerebellum and Brainstem. Mol. Autism 2014, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Heiser, M.; Iacoboni, M.; Maeda, F.; Marcus, J.; Mazziotta, J.C. the Essential Role of Broca’s Area in Imitation. Eur. J. Neurosci. 2003, 17, 1123–1128. [Google Scholar] [CrossRef]
- Koski, L. Modulation of Motor and Premotor Activity During Imitation of Target-Directed Actions. Cereb. Cortex 2002, 12, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Debrabant, J.; Gheysen, F.; Caeyenberghs, K.; Van Waelvelde, H.; Vingerhoets, G. Neural Underpinnings of Impaired Predictive Motor Timing in Children with Developmental Coordination Disorder. Res. Dev. Disabil. 2013, 34, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, M.; Iwaki, S.; Narumi, Y.; Tamai, H.; Suzuki, S. Parietal Dysfunction in Developmental Coordination Disorder: A Functional MRI Study. Neuroreport 2009, 20, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.E.; Licari, M.K.; Billington, J.; Chen, Y.; Aziz-Zadeh, L.; Werner, J.; Winsor, A.M.; Bynevelt, M. Mirror Neuron Activation in Children with Developmental Coordination Disorder: A Functional MRI Study. Int. J. Dev. Neurosci. 2015, 47, 309–319. [Google Scholar] [CrossRef]
- Fuelscher, I.; Caeyenberghs, K.; Enticott, P.G.; Williams, J.; Lum, J.; Hyde, C. Differential Activation of Brain Areas in Children with Developmental Coordination Disorder During Tasks of Manual Dexterity: An ALE Meta-Analysis. Neurosci. Biobehav. Rev. 2018, 86, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hofmann, J. Action Observation and Imitation in Autism Spectrum Disorders: An ALE Meta-Analysis of FMRI Studies. Brain Imaging Behav. 2016, 10, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.G.; Waiter, G.D.; Gilchrist, A.; Perrett, D.I.; Murray, A.D.; Whiten, A. Neural Mechanisms of Imitation and “mirror Neuron” Functioning in Autistic Spectrum Disorder. Neuropsychologia 2006, 44, 610–621. [Google Scholar] [CrossRef]
- Ramachandran, V.S.; Oberman, L.M. Broken Mirrors: A Theory of Autism. Sci. Am. 2006, 295, 62–69. [Google Scholar] [CrossRef]
- Iacoboni, M. Imitation, Empathy, and Mirror Neurons. Annu. Rev. Psychol. 2009, 60, 653–670. [Google Scholar] [CrossRef]
- Chartrand, T.L.; Bargh, J.A. the Chameleon Effect: The Perception-Behavior Link and Social Interaction. J. Pers. Soc. Psychol. 1999, 76, 893–910. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fabbri-destro, M.; Cattaneo, L. Mirror Neurons and Their Clinical Relevance. Neurology 2009, 5, 24–34. [Google Scholar] [CrossRef]
- Grèzes, J.; Wicker, B.; Berthoz, S.; de Gelder, B. A Failure to Grasp the Affective Meaning of Actions in Autism Spectrum Disorder Subjects. Neuropsychologia 2009, 47, 1816–1825. [Google Scholar] [CrossRef]
- Schulte-Rüther, M.; Greimel, E.; Markowitsch, H.J.; Kamp-Becker, I.; Remschmidt, H.; Fink, G.R.; Piefke, M. Dysfunctions in Brain Networks Supporting Empathy: An FMRI Study in Adults with Autism Spectrum Disorders. Soc. Neurosci. 2011, 6, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-T.; Decety, J.; Yang, C.-Y.; Liu, J.-L.; Cheng, Y. Unbroken Mirror Neurons in Autism Spectrum Disorders. J. Child Psychol. Psychiatry 2010, 51, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Pokorny, J.J.; Hatt, N.V.; Colombi, C.; Vivanti, G.; Rogers, S.J.; Rivera, S.M. The Action Observation System When Observing Hand Actions in Autism and Typical Development. Autism Res. 2015, 8, 284–296. [Google Scholar] [CrossRef]
- Raymaekers, R.; Wiersema, J.R.; Roeyers, H. EEG Study of the Mirror Neuron System in Children with High Functioning Autism. Brain Res. 2009, 1304, 113–121. [Google Scholar] [CrossRef]
- Mostofsky, S.H.; Powell, S.K.; Simmonds, D.J.; Goldberg, M.C.; Caffo, B.; Pekar, J.J. Decreased Connectivity and Cerebellar Activity in Autism during Motor Task Performance. Brain 2009, 132, 2413–2425. [Google Scholar] [CrossRef] [PubMed]
- Floris, D.L.; Barber, A.D.; Nebel, M.B.; Martinelli, M.; Lai, M.-C.; Crocetti, D.; Baron-Cohen, S.; Suckling, J.; Pekar, J.J.; Mostofsky, S.H. Atypical Lateralization of Motor Circuit Functional Connectivity in Children with Autism Is Associated with Motor Deficits. Mol. Autism 2016, 7, 35. [Google Scholar] [CrossRef]
- Denckla, M.B.; Rudel, R. Rapid “Automatized” Naming of Pictured Objects, Colors, Letters and Numbers by Normal Children. Cortex 1974, 10, 186–202. [Google Scholar] [CrossRef]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E.H.J.; Leventhal, B.L.; DiLavore, P.C.; Pickles, A.; Rutter, M. the Autism Diagnostic Schedule—Generic: A Standard Measures of Social and Communication Deficits Associated with the Spectrum of Autism. J. Autism Dev. Disord. 2000, 30, 205–223. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Supekar, K.; Menon, V. Typical and Atypical Development of Functional Human Brain Networks: Insights From Resting-State FMRI. Front. Syst. Neurosci. 2010, 4, 21. [Google Scholar] [CrossRef]
- Hamilton, A.; Grafton, S.T. Action Outcomes Are Represented in Human Inferior Frontoparietal Cortex. Cereb. Cortex 2008, 18, 1160–1168. [Google Scholar] [CrossRef]
- Hamilton, A.F. de C. EPS Prize Lecture: Cognitive Underpinnings of Social Interaction. Q. J. Exp. Psychol. 2015, 68, 417–432. [Google Scholar] [CrossRef]
- Wang, Y.; Hamilton, A. Social Top-Down Response Modulation (STORM): A Model of the Control of Mimicry in Social Interaction. Front. Hum. Neurosci. 2012, 6, 153. [Google Scholar] [CrossRef]
- Hamilton, A.F. Gazing At Me: The Importance of Social Meaning in Understanding Direct-Gaze Cues. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2016, 371, 20150080. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, L.; Ortiz-Tudela, J.; Méndez, C.; Lorda, M.J. Mimicry Deficits in Autism Are Not Just Storm Effects. Res. Autism Spectr. Disord. 2015, 17, 64–69. [Google Scholar] [CrossRef]
- Oberman, L.M.; Winkielman, P.; Ramachandran, V.S. Slow Echo: Facial EMG Evidence for the Delay of Spontaneous, But Not Voluntary, Emotional Mimicry in Children with Autism Spectrum Disorders. Dev. Sci. 2009, 12, 510–520. [Google Scholar] [CrossRef]
- Ubaldi, S.; Barchiesi, G.; Cattaneo, L. Bottom-Up and Top-Down Visuomotor Responses to Action Observation. Cereb. Cortex 2015, 25, 1032–1041. [Google Scholar] [CrossRef]
- Miller, B.L.; Cummings, J.L. The Human Frontal Lobes; Guilford Press: New York, NY, USA, 1999; ISBN 1572303905. [Google Scholar]
- Elliott, R. Executive Functions and Their Disorders. Br. Med. Bull. 2003, 65, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Caggiano, V.; Fogassi, L.; Rizzolatti, G.; Casile, A.; Giese, M.A.; Thier, P. Mirror Neurons Encode the Subjective Value of An Observed Action. Proc. Natl. Acad. Sci. USA 2012, 109, 11848–11853. [Google Scholar] [CrossRef] [PubMed]
- Ávila, C.; Garbin, G.; Sanjuán, A.; Forn, C.; Barrós-Loscertales, A.; Bustamante, J.C.; Rodríguez-Pujadas, A.; Belloch, V.; Parcet, M.A. Frontostriatal Response to Set Switching Is Moderated by Reward Sensitivity. Soc. Cogn. Affect. Neurosci. 2012, 7, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Claramonte, P.; Ávila, C.; Rodríguez-Pujadas, A.; Costumero, V.; Ventura-Campos, N.; Bustamante, J.C.; Rosell-Negre, P.; Barrós-Loscertales, A. Inferior Frontal Cortex Activity Is Modulated by Reward Sensitivity and Performance Variability. Biol. Psychol. 2016, 114, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Scott-Van Zeeland, A.A.; Dapretto, M.; Ghahremani, D.G.; Poldrack, R.A.; Bookheimer, S.Y. Reward Processing in Autism. Autism Res. 2010, 3, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Sims, T.B.; Neufeld, J.; Johnstone, T.; Chakrabarti, B. Autistic Traits Modulate Frontostriatal Connectivity during Processing of Rewarding Faces. Soc. Cogn. Affect. Neurosci. 2013, 9, 2010–2016. [Google Scholar] [CrossRef]
- Aziz-Zadeh, L.; Kilroy, E.; Corcelli, G. Understanding Activation Patterns in Shared Circuits: Toward a Value Driven Model. Front. Hum. Neurosci. 2018, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lestou, V.; Pollick, F.E.; Kourtzi, Z. Neural Substrates for Action Understanding at Different Description Levels in the Human Brain. J. Cogn. Neurosci. 2008, 20, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Poulin-Lord, M.P.; Barbeau, E.B.; Soulières, I.; Monchi, O.; Doyon, J.; Benali, H.; Mottron, L. Increased Topographical Variability Of Task-Related Activation In Perceptive And Motor Associative Regions In Adult Autistics. NeuroImage Clin. 2014, 4, 444–453. [Google Scholar] [CrossRef]
- Hannant, P.; Cassidy, S.; Van de Weyer, R.; Mooncey, S. Sensory and Motor Differences in Autism Spectrum Conditions and Developmental Coordination Disorder in Children: A Cross-Syndrome Study. Hum. Mov. Sci. 2018, 58, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Bogdashina, O. Sensory perceptual issues in autism and asperger syndrome: different sensory experiences-different perceptual worlds; Jessica Kingsley Publishers: London, UK, 2016. [Google Scholar]
- Licari, M.K.; Billington, J.; Reid, S.L.; Wann, J.P.; Elliott, C.M.; Winsor, A.M.; Robins, E.; Thornton, A.L.; Jones, R.; Bynevelt, M. Cortical Functioning In Children With Developmental Coordination Disorder: A Motor Overflow Study. Exp. Brain Res. 2015, 233, 1703–1710. [Google Scholar] [CrossRef]
- Pangelinan, M.M.; Hatfield, B.D.; Clark, J.E. Differences in Movement-Related Cortical Activation Patterns Underlying Motor Performance in Children with and Without Developmental Coordination Disorder. J. Neurophysiol. 2013, 109, 3041–3050. [Google Scholar] [CrossRef]
- Reynolds, J.E.; Thornton, A.L.; Elliott, C.; Williams, J.; Lay, B.S.; Licari, M.K. A Systematic Review of Mirror Neuron System Function in Developmental Coordination Disorder: Imitation, Motor Imagery, and Neuroimaging Evidence. Res. Dev. Disabil. 2015, 47, 234–283. [Google Scholar] [CrossRef]
- Hyde, C.; Fuelscher, I.; Williams, J.; Lum, J.A.G.; He, J.; Barhoun, P.; Enticott, P.G. Corticospinal Excitability During Motor Imagery Is Reduced in Young Adults with Developmental Coordination Disorder. Res. Dev. Disabil. 2018, 72, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.E.; Billington, J.; Kerrigan, S.; Williams, J.; Elliott, C.; Winsor, A.M.; Codd, L.; Bynevelt, M.; Licari, M.K. Mirror neuron system activation in children with developmental coordination disorder: A replication functional MRI study. Res. Dev. Disabil. 2017. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.; Sugden, D.; Barnett, A. Movement Assessment Battery for Children [Examiner’s Manual], 2nd ed.; Pearson Assessment: London, UK, 2007. [Google Scholar]
- Kashuk, S. Investigation of MRI Brain Changes in Developmental Coordination Disorder and Friedreich’s Ataxia. Ph.D. Dissertation, Victoria University, Melbourne, Australia, 2017. [Google Scholar]
- Biotteau, M.; Chaix, Y.; Blais, M.; Tallet, J.; Péran, P.; Albaret, J.M. Neural signature of DCD: A critical review of MRI neuroimaging studies. Front. Neurol. 2016, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- David, S.P.; Ware, J.J.; Chu, I.M.; Loftus, P.D.; Fusar-Poli, P.; Radua, J.; Munafò, M.R.; Ioannidis, J.P.A. Potential Reporting Bias in FMRI Studies of the Brain. PLoS ONE 2013, 8, e70104. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.; Miller, M.; Wolford, G. Neural Correlates of Interspecies Perspective Taking in the Post-Mortem Atlantic Salmon: An Argument for Multiple Comparisons Correction. Neuroimage 2009. [Google Scholar] [CrossRef]
- Werner, J.; Cermak, S.A.; Aziz-Zadeh, L. Neural Correlates of Developmental Coordination Disorder: The Mirror Neuron System Hypothesis. J. Behav. Brain Sci. 2012, 2, 258–268. [Google Scholar] [CrossRef]
- Mesulam, M.-M. Large-Scale Neurocognitive Networks and Distributed Processing for Attention, Language, and Memory. Ann. Neurol. 1990, 28, 597–613. [Google Scholar] [CrossRef]
- Lynch, C.J.; Uddin, L.Q.; Supekar, K.; Khouzam, A.; Phillips, J.; Menon, V. Default Mode Network in Childhood Autism: Posteromedial Cortex Heterogeneity and Relationship with Social Deficits. Biol. Psychiatry 2013, 74, 212–219. [Google Scholar] [CrossRef]
- Mennes, M.; Kelly, C.; Zuo, X.N.; Di Martino, A.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Inter-Individual Differences in Resting-State Functional Connectivity Predict Task-Induced BOLD Activity. Neuroimage 2010, 50, 1690–1701. [Google Scholar] [CrossRef]
- Raichle, M.E. the Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef]
- Rudie, J.D.; Shehzad, Z.; Hernandez, L.M.; Colich, N.L.; Bookheimer, S.Y.; Iacoboni, M.; Dapretto, M. Reduced Functional Integration and Segregation of Distributed Neural Systems Underlying Social and Emotional Information Processing in Autism Spectrum Disorders. Cereb. Cortex 2012, 22, 1025–1037. [Google Scholar] [CrossRef]
- Kana, R.K.; Libero, L.E.; Hu, C.P.; Deshpande, H.D.; Colburn, J.S. Functional Brain Networks and White Matter Underlying Theory-Of-Mind in Autism. Soc. Cogn. Affect. Neurosci. 2014, 9, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Alaerts, K.; Nayar, K.; Kelly, C.; Raithel, J.; Milham, M.P.; Di martino, A. Age-Related Changes in Intrinsic Function of the Superior Temporal Sulcus in Autism Spectrum Disorders. Soc. Cogn. Affect. Neurosci. 2015, 10, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Fishman, I.; Keown, C.L.; Lincoln, A.J.; Pineda, J.A.; Müller, R.A. Atypical Cross Talk Between Mentalizing and Mirror Neuron Networks in Autism Spectrum Disorder. JAMA Psychiatry 2014, 71, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.; Shen, M.; Öttl, B.; Keehn, B.; Gaffrey, M.S.; Müller, R.A. Atypical Network Connectivity for Imitation in Autism Spectrum Disorder. Neuropsychologia 2010, 48, 2931–2939. [Google Scholar] [CrossRef]
- Nebel, M.B.; Eloyan, A.; Nettels, C.A.; Sweeney, K.L. Intrinsic Visual-Motor Synchrony Correlates with Social Deficits in Autism. Biol. Psychiatry 2016, 79, 633–641. [Google Scholar] [CrossRef]
- Green, S.A.; Hernandez, L.; Bookheimer, S.Y.; Dapretto, M. Salience Network Connectivity in Autism Is Related to Brain and Behavioral Markers of Sensory Overresponsivity. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 618–626. [Google Scholar] [CrossRef]
- Cerliani, L.; Mennes, M.; Thomas, R.M.; Di Martino, A.; Thioux, M.; Keysers, C. Increased Functional Connectivity Between Subcortical and Cortical Resting-State Networks in Autism Spectrum Disorder. JAMA Psychiatry 2015, 72, 767. [Google Scholar] [CrossRef]
- Hoffmann, E.; Brück, C.; Kreifelts, B.; Ethofer, T.; Wildgruber, D. Reduced Functional Connectivity to the Frontal Cortex During Processing of Social Cues in Autism Spectrum Disorder. J. Neural Transm. 2016, 126, e91–e92. [Google Scholar] [CrossRef]
- von dem Hagen, E.A.H.; Stoyanova, R.S.; Baron-Cohen, S.; Calder, A.J. Reduced Functional Connectivity within and between “Social” Resting State Networks in Autism Spectrum Conditions. Soc. Cogn. Affect. Neurosci. 2013, 8, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Querne, L.; Berquin, P.; Vernier-Hauvette, M.-P.; Fall, S.; Deltour, L.; Meyer, M.-E.; de Marco, G. Dysfunction Of The Attentional Brain Network In Children With Developmental Coordination Disorder: A FMRI Study. Brain Res. 2008, 1244, 89–102. [Google Scholar] [CrossRef] [PubMed]
- McLeod, K.R.; Langevin, L.M.; Goodyear, B.G.; Dewey, D.; Marie, L.; Goodyear, B.G.; Dewey, D.; Langevin, L.M.; Goodyear, B.G.; Dewey, D. Functional Connectivity of Neural Motor Networks Is Disrupted in Children with Developmental Coordination Disorder and Attention-Deficit/Hyperactivity Disorder. NeuroImage Clin. 2014, 4, 566–575. [Google Scholar] [CrossRef] [PubMed]
- McLeod, K.R.; Langevin, L.M.; Dewey, D.; Goodyear, B.G. Atypical within- and between-Hemisphere Motor Network Functional Connections in Children with Developmental Coordination Disorder and Attention-Deficit/Hyperactivity Disorder. NeuroImage Clin. 2016, 12, 157–164. [Google Scholar] [CrossRef]
- Nebel, M.B.; Joel, S.E.; Muschelli, J.; Barber, A.D.; Caffo, B.S.; Pekar, J.J.; Mostofsky, S.H. Disruption of Functional Organization Within the Primary Motor Cortex in Children with Autism. Hum. Brain Mapp. 2014, 35, 567–580. [Google Scholar] [CrossRef]
- Mahajan, R.; Dirlikov, B.; Crocetti, D.; Mostofsky, S.H. Motor Circuit Anatomy in Children with Autism Spectrum Disorder with or without Attention Deficit Hyperactivity Disorder. Autism Res. 2016, 9, 67–81. [Google Scholar] [CrossRef]
- Greicius, M.D.; Kaustubh, S.; Menon, V.; Dougherty, R.F. Resting-State Functional Connectivity Reflects Structural Connectivity in the Default Mode Network. Cereb. Cortex 2009, 19, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Thiebaut de Schotten, M. A Diffusion Tensor Imaging Tractography Atlas for Virtual in Vivo Dissection. Cortex 2008, 44, 1105–1132. [Google Scholar] [CrossRef] [PubMed]
- Ameis, S.H.; Catani, M. Altered white matter connectivity as a neural substrate for social impairment in Autism Spectrum Disorder. Cortex 2015, 62, 158–181. [Google Scholar] [CrossRef]
- Chien, H.Y.; Gau, S.S.F.; Hsu, Y.C.; Chen, Y.J.; Lo, Y.C.; Shih, Y.C.; Tseng, W.Y.I. Altered Cortical Thickness and Tract Integrity of the Mirror Neuron System and Associated Social Communication in Autism Spectrum Disorder. Autism Res. 2015, 8, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Abe, O.; Nippashi, Y.; Yamasue, H. Comparison of White Matter Integrity Between Autism Spectrum Disorder Subjects and Typically Developing Individuals: A Meta-Analysis of Diffusion Tensor Imaging Tractography Studies. Mol. Autism 2013, 4, 25. [Google Scholar] [CrossRef]
- Koldewyn, K.; Yendiki, A.; Weigelt, S.; Gweon, H.; Julian, J.; Richardson, H.; Malloy, C.; Saxe, R.; Fischl, B.; Kanwisher, N. Differences in the Right Inferior Longitudinal Fasciculus but No General Disruption of White Matter Tracts in Children with Autism Spectrum Disorder. Proc. Natl. Acad. Sci. USA 2014, 111, 1981–1986. [Google Scholar] [CrossRef]
- Hofer, S.; Frahm, J. Topography of the Human Corpus Callosum Revisited-Comprehensive Fiber Tractography Using Diffusion Tensor Magnetic Resonance Imaging. Neuroimage 2006, 32, 989–994. [Google Scholar] [CrossRef]
- De Lacoste, C.; Kirkpatrick, J.; Ross, E. Topography of the Human Corpus Callosum. J. Neuropathol. Exp. Neurol. 1985, 44, 578–591. [Google Scholar] [CrossRef]
- Wahl, M.; Lauterbach-Soon, B.; Hattingen, E.; Jung, P.; Singer, O.; Volz, S.; Klein, J.C.; Steinmetz, H.; Ziemann, U. Human Motor Corpus Callosum: Topography, Somatotopy, and Link between Microstructure and Function. J. Neurosci. 2007, 27, 12132–12138. [Google Scholar] [CrossRef] [PubMed]
- Paul, L.K.; Corsello, C.; Kennedy, D.P.; Adolphs, R. Agenesis of the Corpus Callosum and Autism: A Comprehensive Comparison. Brain 2014, 137, 1813–1829. [Google Scholar] [CrossRef] [PubMed]
- Egaas, B.; Courchesne, E.; Saitoh, O. Reduced Size of Corpus Callosum in Autism. Archives of neurology 1995, 52, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Frazier, T.W.; Hardan, A.Y. A Meta-Analysis of the Corpus Callosum in Autism. Biol. Psychiatry 2009, 66, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Hardan, A.Y.; Pabalan, M.; Gupta, N.; Bansal, R.; Melhem, N.M.; Fedorov, S.; Keshavan, M.S.; Minshew, N.J. Corpus Callosum Volume in Children with Autism. Psychiatry Res. Neuroimaging 2009, 174, 57–61. [Google Scholar] [CrossRef]
- Keary, C.J.; Minshew, N.J.; Bansal, R.; Goradia, D.; Fedorov, S.; Keshavan, M.S.; Hardan, A.Y. Corpus Callosum Volume and Neurocognition in Autism. J. Autism Dev. Disord. 2009, 39, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Piven, J.; Bailey, J.; Ranson, B.J.; Arndt, S. An MRI Study of the Corpus Callosum in Autism. Am. J. Psychiatry 1997, 154, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.N.; Nicolson, R.; DeVito, T.J. Mapping Corpus Callosum Deficits in Autism: An Index of Aberrant Cortical Connectivity. Biol. Psychiatry 2006, 60, 218–225. [Google Scholar] [CrossRef]
- Tepest, R.; Jacobi, E.; Gawronski, A.; Krug, B.; Möller-Hartmann, W.; Lehnhardt, F.G.; Vogeley, K. Corpus Callosum Size in Adults with High-Functioning Autism and the Relevance of Gender. Psychiatry Res. Neuroimaging 2010, 183, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Debrabant, J.; Vingerhoets, G.; Van Waelvelde, H.; Leemans, A.; Taymans, T.; Caeyenberghs, K. Brain Connectomics of Visual-Motor Deficits in Children with Developmental Coordination Disorder. J. Pediatr. 2016, 169, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Langevin, L.M.; Macmaster, F.P.; Crawford, S.G.; Lebel, C.; Dewey, D. Common White Matter Microstructure Alterations in Pediatric Motor and Attention Disorders. J. Pediatr. 2014, 164. [Google Scholar] [CrossRef] [PubMed]
- Zwicker, J.G.; Missiuna, C.; Harris, S.R.; Boyd, L.A. Developmental Coordination Disorder: A Pilot Diffusion Tensor Imaging Study. Pediatr. Neurol. 2012, 46, 162–167. [Google Scholar] [CrossRef]
- Hyde, C.; Fuelscher, I.; Enticott, P.G.; Jones, D.K.; Farquharson, S.; Silk, T.J.; Williams, J.; Caeyenberghs, K. White Matter Organization in Developmental Coordination Disorder: A Pilot Study Exploring the Added Value of Constrained Spherical Deconvolution. NeuroImage Clin. 2019, 21, 101625. [Google Scholar] [CrossRef]
- Caeyenberghs, K.; Taymans, T.; Wilson, P.H.; Vanderstraeten, G.; Hosseini, H.; van Waelvelde, H. Neural Signature of Developmental Coordination Disorder in the Structural Connectome Independent of Comorbid Autism. Dev. Sci. 2016, 19, 599–612. [Google Scholar] [CrossRef]
- Lim, C.Y.; Tan, P.C.; Koh, C.; Koh, E.; Guo, H.; Yusoff, N.D.; See, C.Q.; Tan, T. Beery-Buktenica Developmental Test of Visual-Motor Integration (Beery-VMI): Lessons From Exploration of Cultural Variations in Visual-Motor Integration Performance of Preschoolers. Child. Care. Health Dev. 2015, 41, 213–221. [Google Scholar] [CrossRef]
- Rudie, J.D.; Brown, J.A.; Beck-Pancer, D.; Hernandez, L.M.; Dennis, E.L.; Thompson, P.M.; Bookheimer, S.Y.; Dapretto, M. Altered Functional and Structural Brain Network Organization in Autism. NeuroImage Clin. 2012, 2, 79–94. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Aldinger, K.A.; Ashwood, P.; Bauman, M.L.; Blaha, C.D.; Blatt, G.J.; Chauhan, A.; Chauhan, V.; Dager, S.R.; Dickson, P.E.; et al. Consensus paper: Pathological role of the cerebellum in Autism. Cerebellum 2012, 11, 777–807. [Google Scholar] [CrossRef] [PubMed]
- Brown-Lum, M.; Zwicker, J.G. Brain Imaging Increases Our Understanding of Developmental Coordination Disorder: A Review of Literature and Future Directions. Curr. Dev. Disord. Rep. 2015, 2, 131–140. [Google Scholar] [CrossRef]


{kind=link}
| Author | Population | Sample Size | Age (years) | DCD Inclusion | Motor measures | Task | AON Results (DCD vs. TD) |
|---|---|---|---|---|---|---|---|
| Querne et al. 2008 | DCD, TD | n = 19 DCD = 9 TD = 10 | 8–13 | DSM-IV criteria for DCD | Sensori-motor functions scales (NEPSY) | Go/no-go | none |
| Kashiwagi et al. 2009 | DCD, TD | n = 24 DCD = 12 TD = 12 | 9–12 | Below 15th percentile > 2 positive SNS Parent interview | MABC | Visuomotor task (visually guided tracking task) | Decreased activation: left posterior parietal cortex and left postcentral gyrus |
| Zwicker et al. 2010 | DCD, TD | n = 14 DCD = 7 TD = 7 | 8–12 | Below 15th percentile | MABC-2 | Trail-tracing task | Decreased activation: IFG Increased activation: left IPL |
| Zwicker et al. 2011 | DCD, TD | n = 14 DCD = 7 TD = 7 | 8–12 | Below 15th percentile | MABC-2; DCDQ | Trail-tracing task | Decreased activation: bilateral IPL |
| Debrabant et al 2013 | DCD, TD | n = 34 DCD = 17 TD = 17 | 7–10 | Below 5th percentile | MABC-2 | Visuomotor reaction time task | none |
| Licari et al. 2015 | DCD, TD | n = 26 DCD = 13 TD = 13 | 8–10 | Below 5th percentile | MABC-2 | *Finger sequencing task Hand clenching task | Decreased activation: left IFG |
| Reynolds et al. 2015 | DCD, TD | n = 26 DCD = 14 TD = 12 | 8–12 | Below 16th percentile | MABC-2 | *Imitative finger-sequencing task (1) observation, (2) action execution, and (3) action imitation | None Across decreased IFG during imitation compared to other tasks. |
| Reynolds et al. 2017 | DCD, TD | n = 19 DCD = 10 TD = 9 | 8–12 | Below 16th percentile | The Postural Praxis and Sequencing Praxis (SIPT), MABC-2, hand rotation task | *Finger abduction/adduction (1) action observation; (2) motor imagery; (3) action execution; and (4) action imitation. | none |
| Biotteau et al. 2017 | DD, DCD, DD+DCD | n = 48 DCD = 16 DD =16 DD + DCD = 16 | 8–12 | Below 5th percentile, and outside diagnosis | MABC-2 | Procedural learning finger-tapping task | DCD vs. DD: Increased activation: bilateral premotor cortex |
| Kashuk et al. 2017 | pDCD, TD | n = 23 pDCD = 12 TD = 11 | 18-40 | Below 15th percentile on total, fine motor, or gross motor scores and functional deficit on ADC | MAND ADC | Hand rotation task (no execution/imitation) | none |
| Thornton et al., 2018 | DCD, TD, ADHD, DCD+ADHD | n = 48 DCD = 9 TD = 20 ADHD = 20 DCD + ADHD = 18 | 8–17 | At or below 16th percentile and functional deficit on DCDQ | MABC-2, DCDQ | Go/no go task | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilroy, E.; Cermak, S.A.; Aziz-Zadeh, L. A Review of Functional and Structural Neurobiology of the Action Observation Network in Autism Spectrum Disorder and Developmental Coordination Disorder. Brain Sci. 2019, 9, 75. https://doi.org/10.3390/brainsci9040075
Kilroy E, Cermak SA, Aziz-Zadeh L. A Review of Functional and Structural Neurobiology of the Action Observation Network in Autism Spectrum Disorder and Developmental Coordination Disorder. Brain Sciences. 2019; 9(4):75. https://doi.org/10.3390/brainsci9040075
Chicago/Turabian StyleKilroy, Emily, Sharon A. Cermak, and Lisa Aziz-Zadeh. 2019. "A Review of Functional and Structural Neurobiology of the Action Observation Network in Autism Spectrum Disorder and Developmental Coordination Disorder" Brain Sciences 9, no. 4: 75. https://doi.org/10.3390/brainsci9040075