DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu rubripes, Following an Intramuscular Administration of Tetrodotoxin

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. TTX Administration and Sample Preparation
2.3. TTX Extraction and Quantification
2.4. RNA Extraction for Microarray Analysis
2.5. Preparation of Fluorescently-Labeled cRNA and Microarray Analysis
2.6. Quantitative Real-Time PCR
2.7. Data Analysis and Statistics
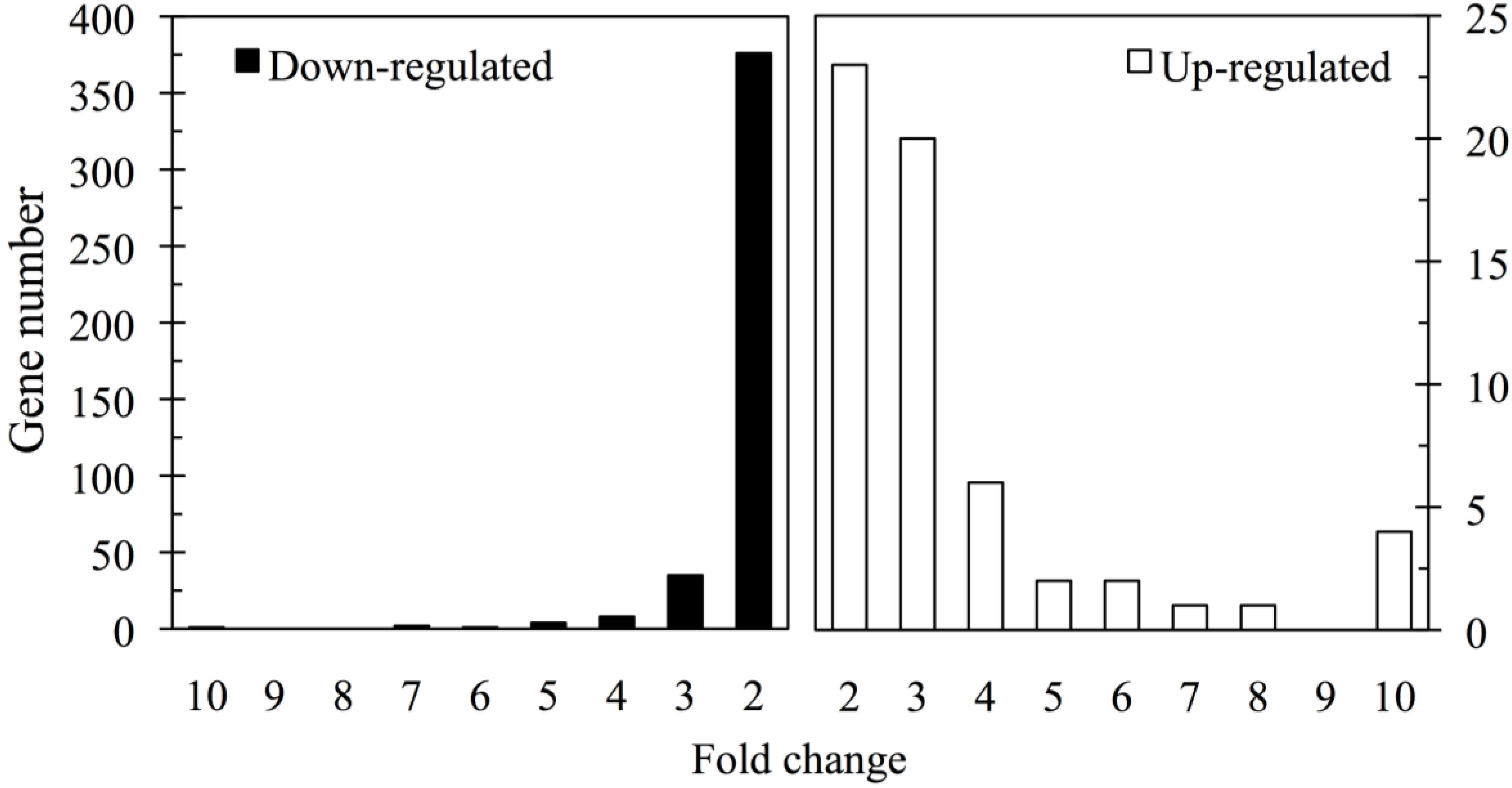
3. Results
3.1. TTX Determination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sex | Body weight (kg) | Dose (µg) | TTX concentration (µg/g) | Total amount (µg) | Accumulation (% of dose) | |||
|---|---|---|---|---|---|---|---|---|---|
| Liver | Skin | Ovary | Muscle | ||||||
| 1 | F | 1.14 | 285 | 0.48 | 0.30 | 5.43 | N.D.1 | 145 | 51 |
| 2 | F | 0.84 | 210 | 0.68 | 0.97 | 5.30 | N.D. | 162 | 77 |
| 3 | F | 1.00 | 250 | 1.12 | 0.62 | 4.97 | N.D. | 206 | 82 |
| 4 | F | 1.10 | 275 | 0.54 | 0.33 | 4.51 | N.D. | 124 | 45 |
| 5 | M | 0.86 | 215 | 1.35 | 0.79 | N.D. 1 | N.D. | 176 | 82 |
| Mean ± SE | 0.99 ± 0.15 | 247 ± 15 | 0.84 ± 0.17 | 0.60 ± 0.13 | 5.05 ± 0.21 | N.D. | 163 ± 14 | 67 ± 8 | |
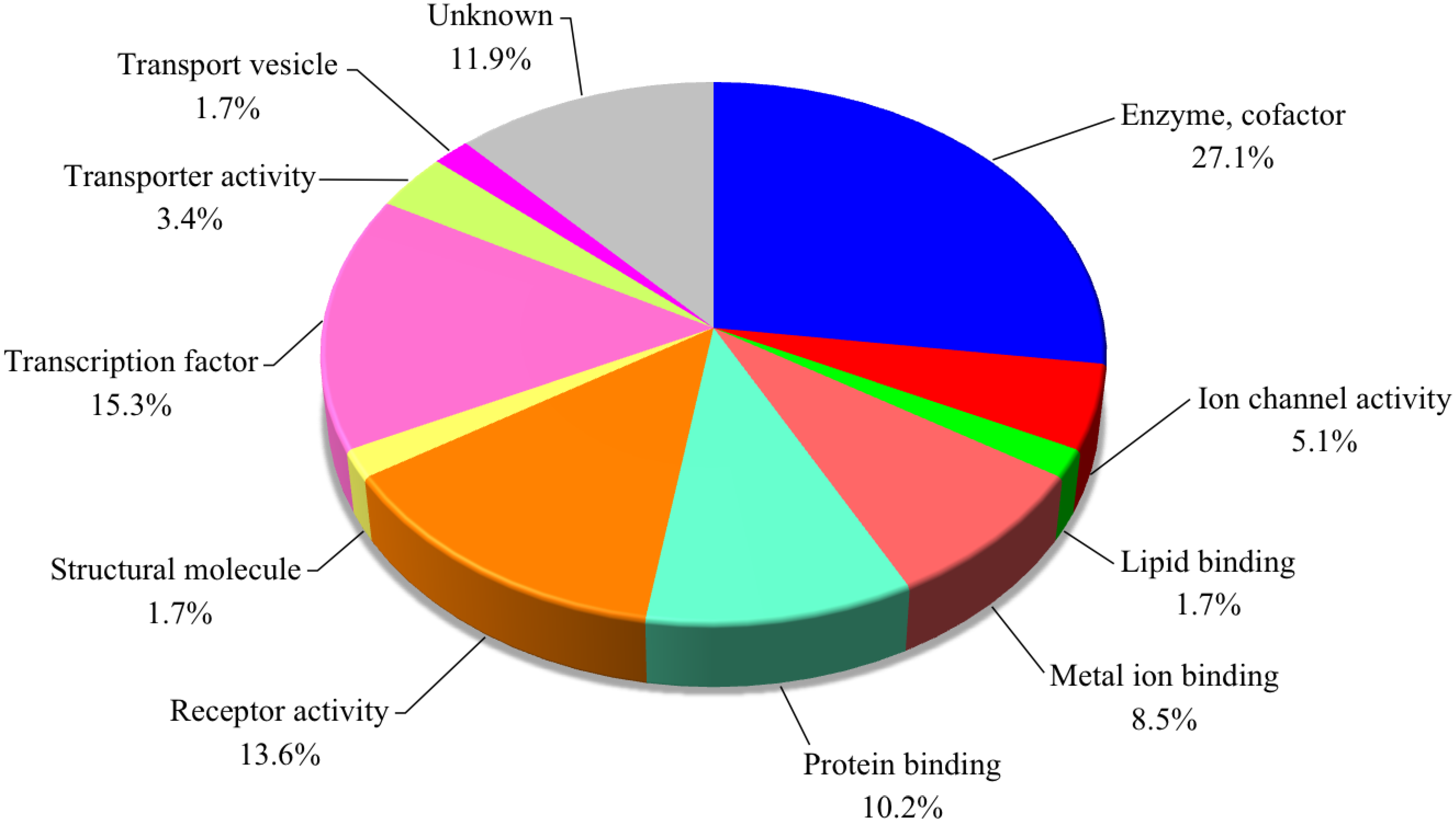
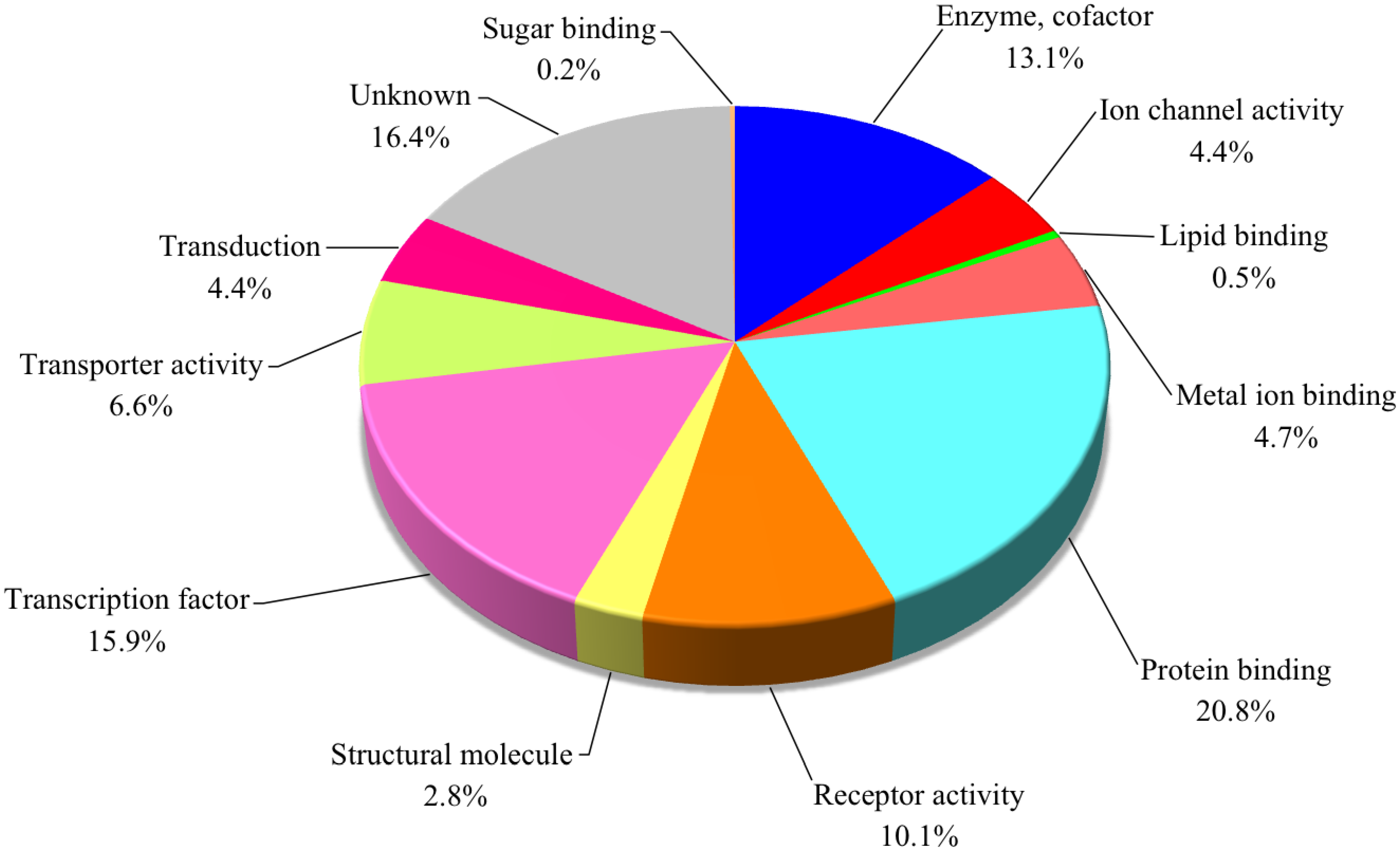
3.2. Gene Expression Analysis


| Ensemble ID | Gene name | Predicted description | Functional classification | FC |
|---|---|---|---|---|
| ENSTRUT00000045544 | Cela2a | Chymotrypsin-like elastase family member 2A | Enzyme, cofactor | 37.6 |
| ENSTRUT00000041722 | Tmem168 | Transmembrane protein 168 | Transport vesicle | 20.0 |
| ENSTRUT00000006023 | Arhgap29 | Rho GTPase-activating protein 29 | Enzyme, cofactor | 12.1 |
| ENSTRUT00000034782 | Kcnma1 | T. rubripes calcium channel alpha-1 subunit homolog (AF026198.1) | Ion channel activity | 10.4 |
| ENSTRUT00000004724 | Chrna9 | T. rubripes nicotinic acetylcholine receptor alpha 9d subunit (AY299471.1) | Ion channel activity | 8.1 |
| ENSTRUT00000038610 | Csmd1 | CUB and sushi domain-containing protein 1 | Protein binding | 7.0 |
| ENSTRUT00000036792 | Serinc4 | Serine incorporator 4 | Transporter activity | 6.8 |
| ENSTRUT00000026094 | C14orf135 | Pecanex-like protein C14orf135 | Unknown | 6.6 |
| ENSTRUT00000046072 | Lrp2 | Low density lipoprotein receptor-related protein 2 | Receptor activity | 5.4 |
| ENSTRUT00000046718 | Col27a1 | Collagen alpha-1(XXVII) chain A-like | Structural molecule | 5.2 |
| ENSTRUT00000000397 | Klhl32 | Kelch-like protein 32 | Protein binding | 4.5 |
| ENSTRUT00000000173 | Elk4 | ELK4, ETS-domain protein | Transcription factor | 4.3 |
| ENSTRUT00000025902 | Zbtb45 | Zinc finger and BTB domain-containing protein 45 | Transcription factor | 4.3 |
| ENSTRUT00000020252 | Spsb1 | SPRY domain-containing SOCS box protein 1 | Receptor activity | 4.3 |
| ENSTRUT00000032342 | Dysf | Dysferlin | Lipid binding | 4.1 |
| ENSTRUT00000003310 | Scn2b | Sodium channel beta-2 subunit | Ion channel activity | 4.0 |
| ENSTRUT00000011167 | Loxl2 | Lysyl oxidase homolog 2 | Enzyme, cofactor | 3.9 |
| ENSTRUT00000017057 | Dmbx1 | Diencephalon/mesencephalon homeobox protein 1 | Transcription factor | 3.9 |
| ENSTRUT00000023088 | Clk2 | Dual specificity protein kinase CLK2 | Enzyme, cofactor | 3.9 |
| ENSTRUT00000045827 | Myo3b | Myosin-IIIb | Enzyme, cofactor | 3.8 |
| ENSTRUT00000025143 | Megf11 | Multiple epidermal growth factor-like domains protein 11 | Metal ion binding | 3.7 |
| ENSTRUT00000003539 | Slc44a1 | Choline transporter-like protein 1 | Transporter activity | 3.7 |
| ENSTRUT00000017798 | Cc4 | Carbonic anhydrase 4 | Enzyme, cofactor | 3.5 |
| ENSTRUT00000006962 | OR4563-2 | T. rubripes odorant receptor (DQ306241.1) | Receptor activity | 3.5 |
| ENSTRUT00000024316 | Pitpnm2 | Membrane-associated phosphatidylinositol transfer protein 2 | Metal ion binding | 3.4 |
| ENSTRUT00000030952 | Ptafr | Platelet-activating factor receptor | Receptor activity | 3.4 |
| ENSTRUT00000038903 | Hepacam2 | HEPACAM family member 2 | Unknown | 3.4 |
| ENSTRUT00000043658 | Gpr22 | Probable G-protein coupled receptor 22 | Receptor activity | 3.3 |
| ENSTRUT00000031306 | Ptpn2 | Tyrosine-protein phosphatase non-receptor type 2 | Enzyme, cofactor | 3.3 |
| ENSTRUT00000019629 | Eps15l1 | Epidermal growth factor receptor substrate 15-like 1 | Receptor activity | 3.2 |
| ENSTRUT00000036590 | Zdhhc8 | Membrane-associated DHHC8 zinc finger protein (NM_001078596.1) | Enzyme, cofactor | 3.2 |
| ENSTRUT00000031045 | Rheb | GTP-binding protein Rheb | Enzyme, cofactor | 3.2 |
| ENSTRUT00000047583 | Agap2 | Arf GAP with GTPase domain, ankyrin repeat and PH domain 2 | Enzyme, cofactor | 3.0 |
| ENSTRUT00000017824 | Ppargc1a | Peroxisome proliferator activated receptor gamma coactivator 1 alpha (DQ157766.1) | Receptor activity | 3.0 |
| ENSTRUT00000027961 | Pfkp | 6-phosphofructokinase type C | Enzyme, cofactor | 3.0 |
| ENSTRUT00000029743 | E2f2 | Transcription factor E2F2 | Unknown | 3.0 |

| Ensemble ID | Gene name | Predicted description | Functional classification | FC | |||
|---|---|---|---|---|---|---|---|
| ENSTRUT00000047556 | Fusa2 | Elongation factor G 2 | Transcription factor | −3.0 | |||
| ENSTRUT00000009260 | Rspo3 | R-spondin-3 | Transduction | −7.8 | |||
| ENSTRUT00000029878 | Ncoa2 | Nuclear receptor coactivator 2 | Transcription factor | −7.3 | |||
| ENSTRUT00000005082 | Sasb | Fatty acyl-CoA hydrolase precursor, medium chain | Enzyme, cofactor | −6.2 | |||
| ENSTRUT00000042623 | Kcnj3 | G protein-activated inward rectifier potassium channel 1 | Ion channel activity | −5.9 | |||
| ENSTRUT00000029012 | Galnt1 | Polypeptide N-acetylgalactosaminyltransferase 1 | Enzyme, cofactor | −5.7 | |||
| ENSTRUT00000034755 | Stk11ip | Serine/threonine kinase 11-interacting protein | Protein binding | −5.5 | |||
| ENSTRUT00000007314 | Unc5d | Netrin receptor UNC5D | Receptor activity | −5.3 | |||
| ENSTRUT00000024559 | Dnmt3a | DNA (cytosine-5)-methyltransferase 3A | Transcription factor | −4.7 | |||
| ENSTRUT00000028271 | finTRIM | Fish virus induced TRIM protein | Metal ion binding | −4.7 | |||
| ENSTRUT00000021561 | Synpo2 | Synaptopodin-2 | Protein binding | −4.4 | |||
| ENSTRUT00000035155 | Klhl8 | Kelch-like protein 8 | Protein binding | −4.4 | |||
| ENSTRUT00000013283 | Lnx2 | Ligand of Numb protein X2 | Metal ion binding | −4.3 | |||
| ENSTRUT00000005282 | Sox5 | T. rubripes transcription factor SOX5 (AY277973.1) | Transcription factor | −4.1 | |||
| ENSTRUT00000004950 | Egr3 | Early growth response protein 3 | Transcription factor | −4.0 | |||
| ENSTRUT00000001876 | Nme1 | Nucleoside diphosphate kinase A | Transcription factor | −4.0 | |||
| ENSTRUT00000037748 | Cadm2 | Cell adhesion molecule 2 | Protein binding | −3.9 | |||
| ENSTRUT00000029148 | Cln3 | Battenin | Enzyme, cofactor | −3.8 | |||
| ENSTRUT00000018412 | Hecd3 | Probable E3 ubiquitin-protein ligase HECTD3 | Protein binding | −3.7 | |||
| ENSTRUT00000010231 | Wipi2 | WD repeat domain phosphoinositide-interacting protein 2 | Enzyme, cofactor | −3.7 | |||
| ENSTRUT00000038056 | Angpt2 | Angiopoietin-2 | Receptor activity | −3.7 | |||
| ENSTRUT00000011781 | - | Putative F-type lectin | Sugar binding | −3.6 | |||
| ENSTRUT00000025271 | Tyro3 | Tyrosine-protein kinase receptor TYRO3 | Transduction | −3.6 | |||
| ENSTRUT00000036834 | Pcolce | T. rubripes procollagen C-endopeptidase enhancer 1 (AF016494.1) | Protein binding | −3.4 | |||
| ENSTRUT00000009581 | Rims1 | Regulating synaptic membrane exocytosis protein 1 | Protein binding | −3.4 | |||
| ENSTRUT00000013940 | Pdlim5 | PDZ and LIM domain protein 5 | Protein binding | −3.4 | |||
| ENSTRUT00000041778 | Foxa3 | T. rubripes folkhead transcription factor FoxA3 (AB604763.1) | Transcription factor | −3.3 | |||
| ENSTRUT00000026385 | Fam70a | Protein FAM70A | Unknown | −3.3 | |||
| ENSTRUT00000003229 | Arhgef26 | Rho guanine nucleotide exchange factor 26 | Transcription factor | −3.2 | |||
| ENSTRUT00000007982 | Kif2c | Kinesin-like protein KIF2C | Protein binding | −3.2 | |||
| ENSTRUT00000013282 | Lnx2 | Ligand of Numb protein X2 | Protein binding | −3.2 | |||
| ENSTRUT00000005850 | Crybb2 | Beta-crystallin A2 | Protein binding | −3.2 | |||
| ENSTRUT00000003860 | Edaradd | Ectodysplasin-A receptor-associated adapter protein | Protein binding | −3.1 | |||
| ENSTRUT00000015819 | Pbxip1 | Pre-B-cell leukemia transcription factor-interacting protein 1 | Transcription factor | −3.1 | |||
| ENSTRUT00000011222 | Serpinh1 | Serpin H1 | Protein binding | −3.1 | |||
| ENSTRUT00000020096 | Atp2b3 | Plasma membrane calcium-transporting ATPase 3 | Transporter activity | −3.1 | |||
| ENSTRUT00000009874 | Plxdc1 | Plexin domain-containing protein 1 | Unknown | −3.1 | |||
| ENSTRUT00000047512 | Suox | Sulfite oxidase, mitochondrial | Enzyme, cofactor | −3.1 | |||
| ENSTRUT00000020028 | finTRIM | Fish virus induced TRIM protein | Metal ion binding | −3.0 | |||
| ENSTRUT00000041535 | Pxn | Paxillin | Protein binding | −3.0 | |||
| ENSTRUT00000033313 | Nox5 | T. rubripes NADPH oxidase 5 (BR000279.1) | Enzyme, cofactor | −3.0 | |||
| ENSTRUT00000027204 | Pacs2 | Phosphofurin acidic cluster sorting protein 2 | Unknown | −3.0 | |||
| ENSTRUT00000044222 | Tgfbrap1 | Transforming growth factor-beta receptor-associated protein 1 | Protein binding | −3.0 | |||
| ENSTRUT00000027017 | Hnrnpk | Heterogeneous nuclear ribonucleoprotein K | Transcription factor | −3.0 | |||
| ENSTRUT00000043644 | Hsp90b1 | Heat shock protein 90kDa beta member 1 | Protein binding | −3.0 | |||
| ENSTRUT00000043157 | Tle3 | T. rubripes transducin-like enhancer protein 3 (AB236415.1) | Transcription factor | −3.0 | |||
| ENSTRUT00000022207 | Vwa1 | Von Willebrand factor A domain-containing protein 1 | Unknown | −3.0 | |||
| ENSTRUT00000026592 | Tacr3 | Neuromedin-K receptor | Receptor activity | −3.0 | |||
| ENSTRUT00000028610 | Dach1 | Dachshund homolog 1 | Protein binding | −3.0 | |||
| ENSTRUT00000011629 | Gas2l3 | GAS2-like protein 3 | Protein binding | −3.0 | |||
| ENSTRUT00000018505 | Epha2 | Ephrin type-A receptor 4 precursor | Receptor activity | −3.0 | |||

4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Halstead, B.W. Poisonous and Venomous Marine Animals of the World, 2nd ed.; The Darwin Press Inc.: Princeton, NJ, USA, 1988; pp. 525–644. [Google Scholar]
- Lee, C.H.; Ruben, P.C. Interaction between voltage-gated sodium channels and the neurotoxin, tetrodotoxin. Channels 2008, 2, 407–412. [Google Scholar] [CrossRef]
- Matsui, T.; Hamada, S.; Konosu, S. Difference in accumulation of puffer fish toxin and crystalline tetrodotoxin in the puffer fish, Fugu rubripes rubripes. Nippon Suisan Gakk. 1981, 47, 535–537. [Google Scholar] [CrossRef]
- Saito, T.; Maruyama, J.; Kanoh, S.; Jeon, J.K.; Noguchi, T.; Harada, T.; Murata, O.; Hashimoto, K. Toxicity of the cultured pufferfish Fugu rubripes rubripes along with their resistibility against tetrodotoxin. Nippon Suisan Gakk. 1984, 50, 1573–1575. [Google Scholar] [CrossRef]
- Honda, S.; Arakawa, O.; Takatani, T.; Tachibana, K.; Yagi, M.; Tanigawa, A.; Noguchi, T. Toxification of cultured puffer fish Takifugu rubripes by feeding on tetrodotoxin-containing diet. Nippon Suisan Gakk. 2005, 71, 815–820. [Google Scholar]
- Noguchi, T.; Arakawa, O.; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp. Biochem. Physiol. Part D: Genom. Proteom. 2006, 1, 153–157. [Google Scholar]
- Kono, M.; Matsui, T.; Furukawa, K.; Yotsu-Yamashita, M.; Yamamori, K. Accumulation of tetrodotoxin and 4,9-anhydrotetrodotoxin in cultured juvenile kusafugu Fugu niphobles by dietary administration of natural toxic komonfugu Fugu poecilonotus liver. Toxicon 2008, 51, 1269–1273. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin-distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O.; Daigo, K.; Hashimoto, K. Local differences in toxin composition of a xanthid crab Atergatis floridus inhabiting Ishigaki Island, Okinawa. Toxicon 1986, 24, 705–711. [Google Scholar] [CrossRef]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Simidu, U.; Noguchi, T.; Hwang, D.F.; Shida, Y.; Hashimoto, K. Marine bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1987, 53, 1714–1715. [Google Scholar]
- Tamplin, M.L. A bacterial source of tetrodotoxins and saxitoxins. ACS Symp. Ser. 1990, 418, 78–106. [Google Scholar]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar]
- Yu, V.C.; Yu, P.H.; Ho, K.C.; Lee, F.W. Isolation and identification of a new tetrodotoxin-producing bacterial species, Raoultella terrigena, from Hong Kong marine puffer fish Takifugu niphobles. Mar. Drugs 2011, 9, 2384–2396. [Google Scholar]
- Yasumoto, T.; Yotsu-Yamashita, M. Chemical and etiological studies on tetrodotoxin and its analogs. J. Toxicol. Toxin Rev. 1996, 15, 81–90. [Google Scholar]
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Evaluation of hepatic uptake clearance of tetrodotoxin in the puffer fish Takifugu rubripes. Toxicon 2008, 52, 369–374. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Pharmacokinetics of tetrodotoxin in puffer fish Takifugu rubripes by a single administration technique. Toxicon 2008, 51, 1051–1059. [Google Scholar] [CrossRef]
- Nagashima, Y.; Toyoda, M.; Hasobe, M.; Shimakura, K.; Shiomi, K. In vitro accumulation of tetrodotoxin in pufferfish liver tissue slices. Toxicon 2003, 41, 569–574. [Google Scholar]
- Matsumoto, T.; Nagashima, Y.; Takayama, K.; Shimakura, K.; Shiomi, K. Difference between tetrodotoxin and saxitoxins in accumulation in puffer fish Takifugu rubripes liver tissue slices. Fish Physiol. Biochem. 2005, 31, 95–100. [Google Scholar]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Sugiyama, Y.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Involvement of carrier-mediated transport system in uptake of tetrodotoxin into liver tissue slices of puffer fish Takifugu rubripes. Toxicon 2007, 50, 173–179. [Google Scholar] [CrossRef]
- Lee, J.H.; Kondo, H.; Sato, S.; Akimoto, S.; Saito, T.; Kodama, M.; Watabe, S. Identification of novel genes related to tetrodotoxin intoxication in pufferfish. Toxicon 2007, 49, 939–953. [Google Scholar] [CrossRef]
- Matsumoto, T.; Ishizaki, S.; Nagashima, Y. Differential gene expression profile in the liver of the marine puffer fish Takifugu rubripes induced by intramuscular administration of tetrodotoxin. Toxicon 2011, 57, 304–310. [Google Scholar] [CrossRef]
- Feroudj, H.; Matsumoto, T.; Kurosu, Y.; Kaneko, G.; Ushio, H.; Suzuki, K.; Kondo, H.; Hirono, I.; Nagashima, Y.; Akimoto, S.; et al. DNA microarray analysis on gene candidates possibly related to tetrodotoxin accumulation in pufferfish. Toxicon 2014, 77, 68–72. [Google Scholar]
- Kodama, K.; Sato, S. Puffer fish toxin. In Standard Methods of Analysis in Food Safety Regulation, Chemistry, 3rd ed.; Ministry of Health Labour and Welfare, Ed.; Japan Food Hygiene Association: Tokyo, Japan, 2005; Volume 1, pp. 661–666. [Google Scholar]
- Nagashima, Y.; Maruyama, J.; Noguchi, T.; Hashimoto, K. Analysis of paralytic shellfish poison and tetrodotoxin by ion-pairing high-performance liquid-chromatography. Nippon Suisan Gakk. 1987, 53, 819–823. [Google Scholar] [CrossRef]
- Shoji, Y.; Yotsu-Yamashita, M.; Miyazawa, T.; Yasumoto, T. Electrospray ionization mass spectrometry of tetrodotoxin and its analogs: Liquid chromatography/mass spectrometry, tandem mass spectrometry, and liquid chromatography/tandem mass spectrometry. Anal. Biochem. 2001, 290, 10–17. [Google Scholar] [CrossRef]
- Agilent eArray Application—eArray User Login Site in Agilent Technologies Homepage. Available online: https://earray.chem.agilent.com/earray/ (accessed on 22 October 2014).
- Fugu Genome Project. Available online: http://www.fugu-sg.org/ (accessed on 22 October 2014).
- Feature Extraction ver. 10.7.3—download software site in Agilent Technologies Homepage. Available online: http://www.genomics.agilent.com/article.jsp?pageId=2059&_requestid=928092 (accessed on 22 October 2014).
- GeneSpring Support—user guides site in Agilent Technologies Homepage. Available online: http://genespring-support.com/support/documentation (accessed on 22 October 2014).
- Primer Express Version 3.0—Getting Started Guide in Applied Biosystems Homepage. Available online: https://www3.appliedbiosystems.com/cms/groups/mcb_support/documents/generaldocuments/cms_041902.pdf (accessed on 22 October 2014).
- Gene Ontology Consortium. Available online: http://geneontology.org/ (accessed on 22 October 2014).
- Watabe, S.; Sato, Y.; Nakaya, M.; Nogawa, N.; Oohashi, K.; Noguchi, T.; Morikawa, N.; Hashimoto, K. Distribution of tritiated tetrodotoxin administered intraperitoneally to pufferfish. Toxicon 1987, 25, 1283–1289. [Google Scholar] [CrossRef]
- Ikeda, K.; Murakami, Y.; Emoto, Y.; Ngy, L.; Taniyama, S.; Yagi, M.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to non-toxic cultured specimens of the pufferfish Takifugu rubripes. Toxicon 2009, 53, 99–103. [Google Scholar] [CrossRef]
- Kodama, M.; Ogata, T.; Sato, S. External secretion of tetrodotoxin from puffer fishes stimulated by electric shock. Mar. Biol. 1985, 87, 199–202. [Google Scholar] [CrossRef]
- Kodama, M.; Sato, S.; Ogata, T.; Suzuki, Y.; Kaneko, T.; Aida, K. Tetrodotoxin secreting glands in the skin of puffer fishes. Toxicon 1986, 24, 819–829. [Google Scholar] [CrossRef]
- Saito, T.; Noguchi, T.; Harada, T.; Murata, O.; Hashimoto, K. Tetrodotoxin as a biological defense agent for puffers. Nippon Suisan Gakk. 1985, 51, 1175–1180. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef]
- Tatsuno, R.; Yamaguchi, K.; Takatani, T.; Arakawa, O. RT-PCR- and MALDI-TOF mass spectrometry-based identification and distribution of isoforms homologous to pufferfish saxitoxin- and tetrodotoxin-binding protein in the plasma of non-toxic cultured pufferfish (Takifugu rubripes). Biosci. Biotechnol. Biochem. 2013, 77, 208–212. [Google Scholar]
- Kawashima, I.; Tani, T.; Shimoda, K.; Takiguchi, Y. Characterization of pancreatic elastase II cDNAs: Two elastase II mRNAs are expressed in human pancreas. DNA 1987, 6, 163–172. [Google Scholar]
- Lausen, J.; Liu, S.; Fliegauf, M.; Lübbert, M.; Werner, M.H. ELA2 is regulated by hematopoietic transcription factors, but not repressed by AML1-ETO. Oncogene 2006, 25, 1349–1357. [Google Scholar] [CrossRef]
- Friedman, A.D. Transcriptional regulation of granulocyte and monocyte development. Oncogene 2002, 21, 3377–3390. [Google Scholar] [CrossRef]
- Whitcomb, D.C.; Lowe, M.E. Human pancreatic digestive enzymes. Dig. Dis. Sci. 2007, 52, 1–17. [Google Scholar] [CrossRef]
- Isom, L.L.; Ragsdale, D.S.; De-Jongh, K.S.; Westenbroek, R.E.; Reber, B.F.; Scheuer, T.; Catterall, W.A. Structure and function of the beta 2 subunit of brain sodium channels, a transmembrane glycoprotein with a CAM motif. Cell 1995, 83, 433–442. [Google Scholar] [CrossRef]
- Srinivasan, J.; Schachner, M.; Catterall, W.A. Interaction of voltage-gated sodium channels with the extracellular matrix molecules tenascin-C and tenascin-R. Proc. Natl. Acad. Sci. USA 1998, 95, 15753–15757. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kazen-Gillespie, K.; Hortsch, M.; Isom, L.L. Sodium channel beta subunits mediate homophilic cell adhesion and recruit ankyrin to points of cell-cell contact. J. Biol. Chem. 2000, 275, 11383–11388. [Google Scholar] [CrossRef]
- Pertin, M.; Ji, R.R.; Berta, T.; Powell, A.J.; Karchewski, L.; Tate, S.N.; Isom, L.L.; Woolf, C.J.; Gilliard, N.; Spahn, D.R.; et al. Upregulation of the voltage-gated sodium channel beta2 subunit in neuropathic pain models: Characterization of expression in injured and non-injured primary sensory neurons. J. Neurosci. 2005, 25, 10970–10980. [Google Scholar] [CrossRef]
- Lopez Santiago, L.F.; Pertin, M.; Morisod, X.; Chen, C.; Hong, S.; Wiley, J.; Decosterd, I.; Isom, L.L. Sodium channel beta2 subunits regulate tetrodotoxin-sensitive sodium channels in small dorsal root ganglion neurons and modulate the response to pain. J. Neurosci. 2006, 26, 7984–7994. [Google Scholar] [CrossRef]
- Lek, A.; Evesson, F.J.; Sutton, R.B.; North, K.N.; Cooper, S.T. Ferlins: Regulators of vesicle fusion for auditory neurotransmission, receptor trafficking and membrane repair. Traffic 2012, 13, 185–194. [Google Scholar] [CrossRef]
- De Morrée, A.; Flix, B.; Bagaric, I.; Wang, J.; van den Boogaard, M.; Grand Moursel, L.; Frants, R.R.; Illa, I.; Gallardo, E.; Toes, R.; van der Maarel, S.M. Dysferlin regulates cell adhesion in human monocytes. J. Biol. Chem. 2013, 288, 14147–14157. [Google Scholar]
- Kerr, J.P.; Ward, C.W.; Bloch, R.J. Dysferlin at transverse tubules regulates Ca2+ homeostasis in skeletal muscle. Front. Physiol. 2014. [Google Scholar] [CrossRef]
- Oulhen, N.; Onorato, T.M.; Ramos, I.; Wessel, G.M. Dysferlin is essential for endocytosis in the sea star oocyte. Dev. Biol. 2014, 388, 94–102. [Google Scholar] [CrossRef]
- Kitamura, C.; Takahashi, M.; Kondoh, Y.; Tashiro, H.; Tashiro, T. Identification of synaptic activity-dependent genes by exposure of cultured cortical neurons to tetrodotoxin followed by its withdrawal. J. Neurosci. Res. 2007, 85, 2385–2399. [Google Scholar] [CrossRef]
- Saras, J.; Franzen, P.; Aspenstrom, P.; Hellman, U.; Gonez, L.J.; Heldin, C.H. A novel GTPase-activating protein for Rho interacts with a PDZ domain of the protein-tyrosine phosphatase PTPL1. J. Biol. Chem. 1997, 272, 24333–24338. [Google Scholar]
- Hakoshima, T. Structural basis of the rho GTPase signaling. J. Biochem. 2003, 134, 327–331. [Google Scholar] [CrossRef]
- Myagmar, B.E.; Umikawa, M.; Asato, T.; Taira, K.; Oshiro, M.; Hino, A.; Takei, K.; Uezato, H.; Kariya, K. PARG1, a protein-tyrosine phosphatase-associated RhoGAP, as a putative Rap2 effector. Biochem. Biophys. Res. Commun. 2005, 329, 1046–1052. [Google Scholar]
- Adams, J.W.; Wang, J.; Davis, J.R.; Liaw, C.; Gaidarov, I.; Gatlin, J.; Dalton, N.D.; Gu, Y.; Ross, J., Jr.; Behan, D.; Chien, K.; Connolly, D. Myocardial expression, signaling, and function of GPR22: A protective role for an orphan G protein-coupled receptor. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H509–H521. [Google Scholar] [CrossRef]
- Karassek, S.; Berghaus, C.; Schwarten, M.; Goemans, C.G.; Ohse, N.; Kock, G.; Jockers, K.; Neumann, S.; Gottfried, S.; Herrmann, C.; Heumann, R.; Stoll, R. Ras homolog enriched in brain (Rheb) enhances apoptotic signaling. J. Biol. Chem. 2010, 285, 33979–33991. [Google Scholar]
- Sciarretta, S.; Zhai, P.; Shao, D.; Maejima, Y.; Robbins, J.; Volpe, M.; Condorelli, G.; Sadoshima, J. Rheb is critical regulator of autophagy during myocardial ischemia: pathophysiological implications in obesity and metabolic syndrome. Circulation 2013, 125, 1134–1146. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, T.; Feroudj, H.; Kikuchi, R.; Kawana, Y.; Kondo, H.; Hirono, I.; Mochizuki, T.; Nagashima, Y.; Kaneko, G.; Ushio, H.; et al. DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu rubripes, Following an Intramuscular Administration of Tetrodotoxin. Microarrays 2014, 3, 226-244. https://doi.org/10.3390/microarrays3040226
Matsumoto T, Feroudj H, Kikuchi R, Kawana Y, Kondo H, Hirono I, Mochizuki T, Nagashima Y, Kaneko G, Ushio H, et al. DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu rubripes, Following an Intramuscular Administration of Tetrodotoxin. Microarrays. 2014; 3(4):226-244. https://doi.org/10.3390/microarrays3040226
Chicago/Turabian StyleMatsumoto, Takuya, Holger Feroudj, Ryosuke Kikuchi, Yuriko Kawana, Hidehiro Kondo, Ikuo Hirono, Toshiaki Mochizuki, Yuji Nagashima, Gen Kaneko, Hideki Ushio, and et al. 2014. "DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu rubripes, Following an Intramuscular Administration of Tetrodotoxin" Microarrays 3, no. 4: 226-244. https://doi.org/10.3390/microarrays3040226
APA StyleMatsumoto, T., Feroudj, H., Kikuchi, R., Kawana, Y., Kondo, H., Hirono, I., Mochizuki, T., Nagashima, Y., Kaneko, G., Ushio, H., Kodama, M., & Watabe, S. (2014). DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu rubripes, Following an Intramuscular Administration of Tetrodotoxin. Microarrays, 3(4), 226-244. https://doi.org/10.3390/microarrays3040226