
Neurophysiological Effects of Whole Coffee Cherry Extract in Older Adults with Subjective Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled, Cross-Over Pilot Study

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics and Reproducibility
2.2. Participants and Recruitment
2.3. Procedures
2.3.1. Interventions
2.3.2. MRI Sessions
2.3.3. Exosomal BDNF
2.3.4. Behavioral Tasks
2.4. Chemical Analysis
2.4.1. Chlorogenic Acid
2.4.2. Trigonelline by HPLC
2.4.3. Caffeine
2.4.4. Sample Preparation for Antioxidant Measurements
Peroxyl Radicals Scavenging Capacity (ORAC Assay)
Hydroxyl Radical Scavenging Capacity (HORAC Assay)
Peroxynitrite Scavenging Capacity (NORAC Assay)
Superoxide Anion Scavenging Assay (SORAC Assay)
Singlet Oxygen Scavenging Assay (SOAC Assay)
2.5. Statistical Analysis Plan
2.5.1. Functional Magnetic Resonance Imaging (fMRI)
2.5.2. Magnetic Resonance Spectroscopy (MRS)
2.5.3. Exosomal BDNF and Behavioral Data
3. Results
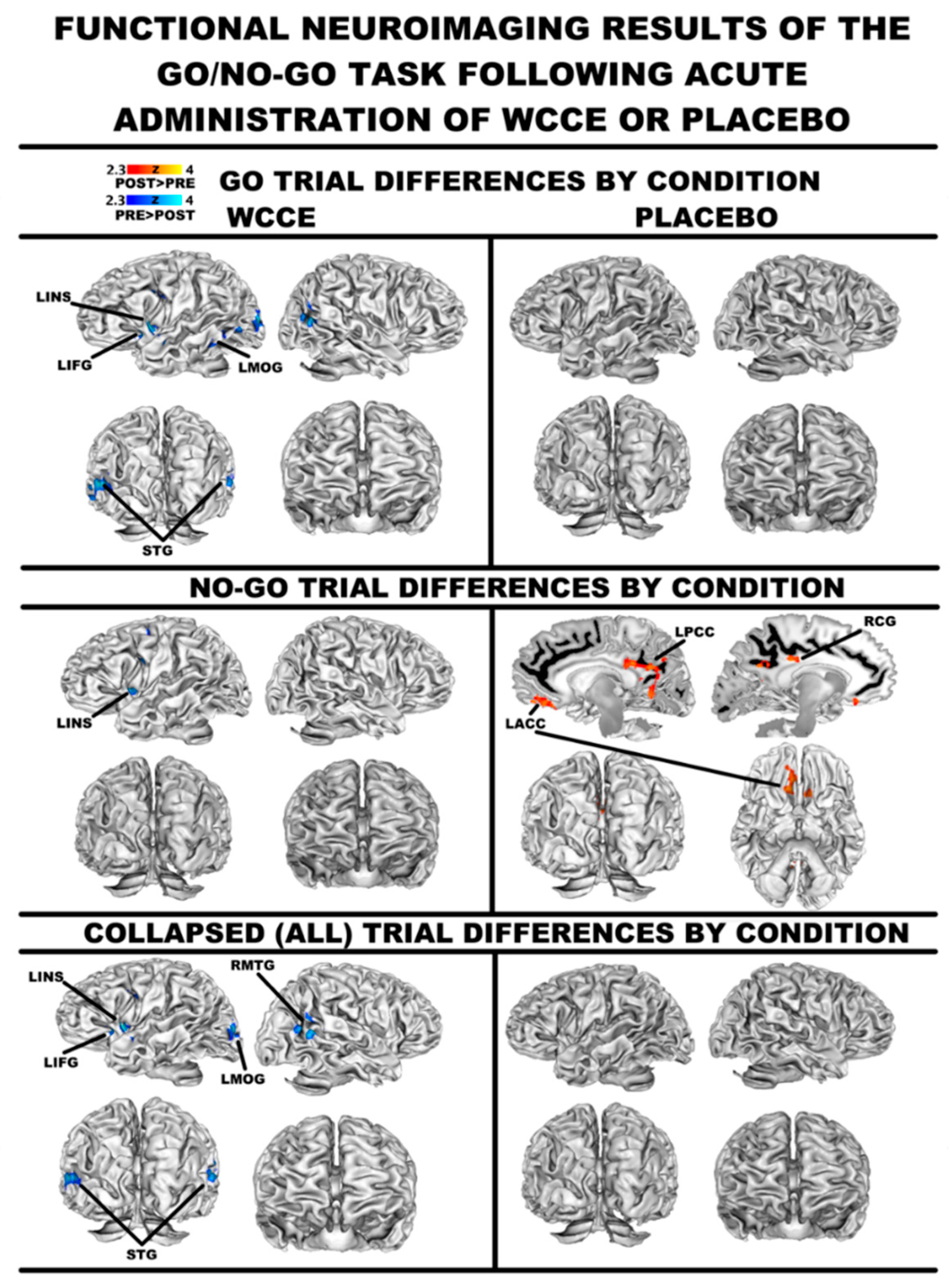
3.1. FMRI: General Linear Modeling
3.2. FMRI: Connectivity
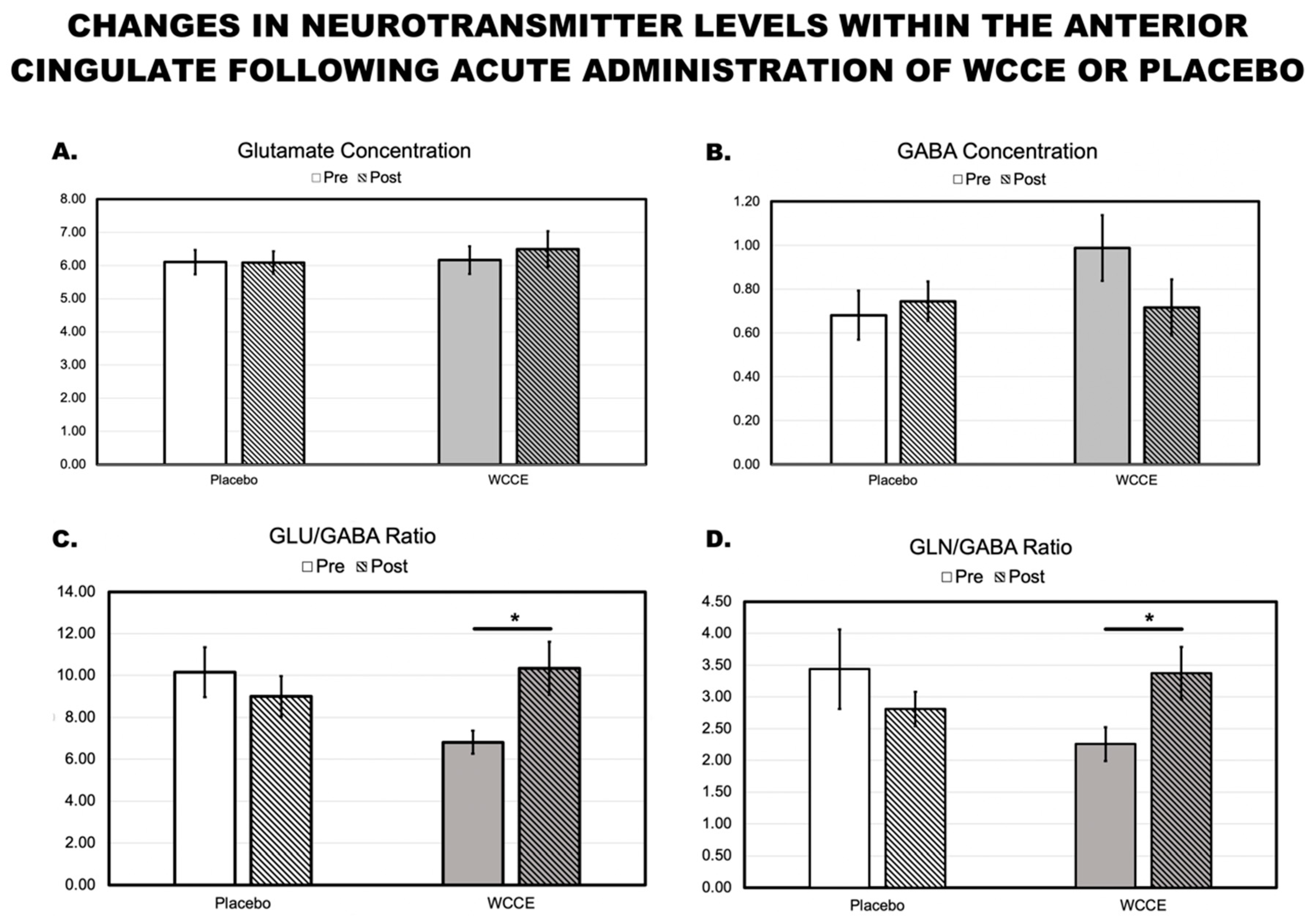
3.3. MRS
3.4. BDNF
3.5. Behavioral Results
3.6. HPLC Analyses
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, H.; Ou, J.; Chen, L.; Zhang, Y.; Szkudelski, T.; Delmas, D.; Daglia, M.; Xiao, J. Dietary polyphenols and type 2 diabetes: Human Study and Clinical Trial. Crit. Rev. Food Sci. Nutr. 2019, 59, 3371–3379. [Google Scholar] [CrossRef]
- Hurtado-Barroso, S.; Quifer-Rada, P.; Rinaldi de Alvarenga, J.; Pérez-Fernández, S.; Tresserra-Rimbau, A.; Lamuela-Raventos, R. Changing to a Low-Polyphenol Diet Alters Vascular Biomarkers in Healthy Men after Only Two Weeks. Nutrients 2018, 10, 1766. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, R.D.; Carvalho, N.C.; Martin-Moreno, J.M.; Pimenta, A.M.; Lopes, A.C.S.; Gea, A.; Martinez-Gonzalez, M.A.; Bes-Rastrollo, M. Total polyphenol intake, polyphenol subtypes and incidence of cardiovascular disease: The SUN cohort study. Nutr. Metab. Cardiovasc. Dis. 2018, 29, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.; Mitchell, E.S. More than just caffeine: Psychopharmacology of methylxanthine interactions with plant-derived phytochemicals. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Sarubbo, F.; Moranta, D.; Pani, G. Dietary polyphenols and neurogenesis: Molecular interactions and implication for brain ageing and cognition. Neurosci. Biobehav. Rev. 2018, 90, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Miller, M.G.; Chu, Y.-F.; Lyle, B.J.; Joseph, J.A. Coffee, but not caffeine, has positive effects on cognition and psychomotor behavior in aging. AGE 2013, 35, 2183–2192. [Google Scholar] [CrossRef]
- Whyte, R.A.; Cheng, N.; Fromentin, E.; Williams, M.C. A Randomized, Double-Blinded, Placebo-Controlled Study to Compare the Safety and Efficacy of Low Dose Enhanced Wild Blueberry Powder and Wild Blueberry Extract (ThinkBlue™) in Maintenance of Episodic and Working Memory in Older Adults. Nutrients 2018, 10, 660. [Google Scholar] [CrossRef]
- Heimbach, J.T.; Marone, P.A.; Hunter, J.M.; Nemzer, B.V.; Stanley, S.M.; Kennepohl, E. Safety studies on products from whole coffee fruit. Food Chem. Toxicol. 2010, 48, 2517–2525. [Google Scholar] [CrossRef]
- Reyes-Izquierdo, T.; Nemzer, B.; Shu, C.; Huynh, L.; Argumedo, R.; Keller, R.; Pietrzkowski, Z. Modulatory effect of coffee fruit extract on plasma levels of brain-derived neurotrophic factor in healthy subjects. Br. J. Nutr. 2013, 110, 420–425. [Google Scholar] [CrossRef]
- Reed, R.A.; Mitchell, E.S.; Saunders, C.; O’Connor, P.J. Acute Low and Moderate Doses of a Caffeine-Free Polyphenol-Rich Coffeeberry Extract Improve Feelings of Alertness and Fatigue Resulting from the Performance of Fatiguing Cognitive Tasks. J. Cogn. Enhanc. 2019, 3, 193–206. [Google Scholar] [CrossRef]
- Reyes-Izquierdo, T.; Argumedo, R.; Shu, C.; Nemzer, B.; Pietrzkowski, Z. Stimulatory effect of whole coffee fruit concentrate powder on plasma levels of total and exosomal brain-derived neurotrophic factor in healthy subjects: An acute within-subject clinical study. Food Nutr. Sci. 2013, 4, 984–990. [Google Scholar] [CrossRef]
- Abdel-Aziz, K.; Larner, A.J. Six-item cognitive impairment test (6CIT): Pragmatic diagnostic accuracy study for dementia and MCI. Int. Psychogeriatr. 2015, 27, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Glorioso, C.; Sabatini, M.; Unger, T.; Hashimoto, T.; Monteggia, L.M.; Lewis, D.A.; Mirnics, K. Specificity and timing of neocortical transcriptome changes in response to BDNF gene ablation during embryogenesis or adulthood. Mol. Psychiatry 2006, 11, 633. Available online: https://www.nature.com/articles/4001835#supplementary-information (accessed on 3 January 2019). [CrossRef] [PubMed]
- Kohara, K.; Yasuda, H.; Huang, Y.; Adachi, N.; Sohya, K.; Tsumoto, T. A Local Reduction in Cortical GABAergic Synapses after a Loss of Endogenous Brain-Derived Neurotrophic Factor, as Revealed by Single-Cell Gene Knock-Out Method. J. Neurosci. 2007, 27, 7234. [Google Scholar] [CrossRef]
- Palizvan, M.R.; Sohya, K.; Kohara, K.; Maruyama, A.; Yasuda, H.; Kimura, F.; Tsumoto, T. Brain-derived neurotrophic factor increases inhibitory synapses, revealed in solitary neurons cultured from rat visual cortex. Neuroscience 2004, 126, 955–966. [Google Scholar] [CrossRef]
- Wang, L.; Chang, X.; She, L.; Xu, D.; Huang, W.; Poo, M.M. Autocrine Action of BDNF on Dendrite Development of Adult-Born Hippocampal Neurons. J. Neurosci. 2015, 35, 8384. [Google Scholar] [CrossRef]
- Oh, H.; Piantadosi, S.C.; Rocco, B.R.; Lewis, D.A.; Watkins, S.C.; Sibille, E. The Role of Dendritic Brain-Derived Neurotrophic Factor Transcripts on Altered Inhibitory Circuitry in Depression. Biol. Psychiatry 2019, 85, 517–526. [Google Scholar] [CrossRef]
- Sun, Z.; Yu, J.; Liu, Y.L.; Hong, Z.; Ling, L.; Li, G.; Zhuo, Y.; Wang, W.; Zhang, Y. Reduced Serum Levels of Brain-Derived Neurotrophic Factor Are Related to Mild Cognitive Impairment in Chinese Patients with Type 2 Diabetes Mellitus. Ann. Nutr. Metab. 2018, 73, 271–281. [Google Scholar] [CrossRef]
- Barha, C.K.; Liu-Ambrose, T.; Best, J.R.; Yaffe, K.; Rosano, C. Sex-dependent effect of the BDNF Val66Met polymorphism on executive functioning and processing speed in older adults: Evidence from the health ABC study. Neurobiol. Aging 2019, 74, 161–170. [Google Scholar] [CrossRef]
- Kumar, A.; Dogra, S.; Sona, C.; Umrao, D.; Rashid, M.; Singh, S.K.; Wahajuddin, M.; Yadav, P.N. Chronic histamine 3 receptor antagonism alleviates depression like conditions in mice via modulation of brain-derived neurotrophic factor and hypothalamus-pituitary adrenal axis. Psychoneuroendocrinology 2019, 101, 128–137. [Google Scholar] [CrossRef]
- Poulose, S.M.; Miller, M.G.; Scott, T.; Shukitt-Hale, B. Nutritional Factors Affecting Adult Neurogenesis and Cognitive Function. Adv. Nutr. 2017, 8, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Barfoot, K.L.; May, G.; Lamport, D.J.; Ricketts, J.; Riddell, P.M.; Williams, C.M. The effects of acute wild blueberry supplementation on the cognition of 7–10-year-old schoolchildren. Eur. J. Nutr. 2019, 58, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Sakakibara, E.; Lin, W.-J.; Wang, J.; Pasinetti, G.M.; Salton, S.R. Grape-derived polyphenols produce antidepressant effects via VGF- and BDNF-dependent mechanisms. Ann. N. Y. Acad. Sci. 2019, 1455, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Caracci, F.; Harary, J.; Simkovic, S.; Pasinetti, G.M. Grape-Derived Polyphenols Ameliorate Stress-Induced Depression by Regulating Synaptic Plasticity. J. Agric. Food Chem. 2020, 68, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Mi, Y.; Wang, Y.; Li, R.; Huang, S.; Li, X.; Liu, X. Neuroprotective action of tea polyphenols on oxidative stress-induced apoptosis through the activation of the TrkB/CREB/BDNF pathway and Keap1/Nrf2 signaling pathway in SH-SY5Y cells and mice brain. Food Funct. 2017, 8, 4421–4432. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Dev. Ther. 2015, 10, 23–42. [Google Scholar] [CrossRef]
- Nemzer, B.; Abshiru, N.; Al-Taher, F. Identification of phytochemical compounds in coffea arabica whole coffee cherries and their extracts by LC-MS/MS. J. Agric. Food Chem. under review.
- Mullen, W.; Nemzer, B.; Stalmach, A.; Ali, S.; Combet, E. Polyphenolic and Hydroxycinnamate Contents of Whole Coffee Fruits from China, India, and Mexico. J. Agric. Food Chem. 2013, 61, 5298–5309. [Google Scholar] [CrossRef]
- Li, L.; Su, C.; Chen, X.; Wang, Q.; Jiao, W.; Luo, H.; Tang, J.; Wang, W.; Li, S.; Guo, S. Chlorogenic Acids in Cardiovascular Disease: A Review of Dietary Consumption, Pharmacology, and Pharmacokinetics. J. Agric. Food Chem. 2020, 68, 6464–6484. [Google Scholar] [CrossRef]
- Mullen, W.; Nemzer, B.; Ou, B.; Stalmach, A.; Hunter, J.; Clifford, M.N.; Combet, E. The Antioxidant and Chlorogenic Acid Profiles of Whole Coffee Fruits Are Influenced by the Extraction Procedures. J. Agric. Food Chem. 2011, 59, 3754–3762. [Google Scholar] [CrossRef]
- Zheng, T.; Liu, H.; Qin, L.; Chen, B.; Zhang, X.; Hu, X.; Xiao, L.; Qin, S. Oxidative stress-mediated influence of plasma DPP4 activity to BDNF ratio on mild cognitive impairment in elderly type 2 diabetic patients: Results from the GDMD study in China. Metab. Clin. Exp. 2018, 87, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Kent, B.A.; Morici, J.F.; Gallo, F.; Saksida, L.M.; Bussey, T.J.; Weisstaub, N.; Bekinschtein, P. NMDA receptors and BDNF are necessary for discrimination of overlapping spatial and non-spatial memories in perirhinal cortex and hippocampus. Neurobiol. Learn. Mem. 2018, 155, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Niculescu, D.; Michaelsen-Preusse, K.; Güner, Ü.; van Dorland, R.; Wierenga, C.J.; Lohmann, C. A BDNF-Mediated Push-Pull Plasticity Mechanism for Synaptic Clustering. Cell Rep. 2018, 24, 2063–2074. [Google Scholar] [CrossRef]
- Zhen, L.; Shao, T.; Luria, V.; Li, G.; Li, Z.; Xu, Y.; Zhao, X. EphB2 Deficiency Induces Depression-Like Behaviors and Memory Impairment: Involvement of NMDA 2B Receptor Dependent Signaling. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.L.; Hunter, J.M.; Reyes-Izquierdo, T.; Argumedo, R.; Brizuela-Bastien, J.; Keller, R.; Pietrzkowski, Z. Cognitive short- and long-term effects of coffee cherry extract in older adults with mild cognitive decline. Aging Neuropsychol. Cogn. 2019, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zafra, F.; Castrén, E.; Thoenen, H.; Lindholm, D. Interplay between glutamate and gamma-aminobutyric acid transmitter systems in the physiological regulation of brain-derived neurotrophic factor and nerve growth factor synthesis in hippocampal neurons. Proc. Natl. Acad. Sci. USA 1991, 88, 10037. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Holmes, M.J.; Newton, A.T.; Morgan, V.L.; Landman, B.A. A Comparison of Distributional Considerations with Statistical Analysis of Resting State fMRI at 3T and 7T. Proc. SPIE-Intern. Soc. Opt. Eng. 2012, 8314, 831416. [Google Scholar] [CrossRef]
- Trattnig, S.; Springer, E.; Bogner, W.; Hangel, G.; Strasser, B.; Dymerska, B.; Cardoso, P.L.; Robinson, S.D. Key clinical benefits of neuroimaging at 7T. NeuroImage 2018, 168, 477–489. [Google Scholar] [CrossRef]
- Pohmann, R.; Speck, O.; Scheffler, K. Signal-to-noise ratio and MR tissue parameters in human brain imaging at 3, 7, and 9.4 tesla using current receive coil arrays. Magn. Reson. Med. 2016, 75, 801–809. [Google Scholar] [CrossRef]
- Reid, M.A.; Salibi, N.; White, D.M.; Gawne, T.J.; Denney, T.S.; Lahti, A.C. 7T Proton Magnetic Resonance Spectroscopy of the Anterior Cingulate Cortex in First-Episode Schizophrenia. Schizophr. Bull. 2018, 45, 180–189. [Google Scholar] [CrossRef]
- Strasser, A.; Xin, L.; Gruetter, R.; Sandi, C. Nucleus accumbens neurochemistry in human anxiety: A 7 T 1H-MRS study. Eur. Neuropsychopharmacol. 2019, 29, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Gordon, E.M.; Laumann, T.O.; Adeyemo, B.; Gilmore, A.W.; Nelson, S.M.; Dosenbach, N.U.F.; Petersen, S.E. Individual-specific features of brain systems identified with resting state functional correlations. NeuroImage 2017, 146, 918–939. [Google Scholar] [CrossRef] [PubMed]
- Gordon, E.M.; Laumann, T.O.; Adeyemo, B.; Petersen, S.E. Individual Variability of the System-Level Organization of the Human Brain. Cereb. Cortex 2015, 27, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Gordon, E.M.; Laumann, T.O.; Gilmore, A.W.; Newbold, D.J.; Greene, D.J.; Berg, J.J.; Ortega, M.; Hoyt-Drazen, C.; Gratton, C.; Sun, H.; et al. Precision Functional Mapping of Individual Human Brains. Neuron 2017, 95, 791–807.e797. [Google Scholar] [CrossRef] [PubMed]
- Laumann, T.O.; Gordon, E.M.; Adeyemo, B.; Snyder, A.Z.; Joo, S.J.; Chen, M.-Y.; Gilmore, A.W.; McDermott, K.B.; Nelson, S.M.; Dosenbach, N.U.F.; et al. Functional System and Areal Organization of a Highly Sampled Individual Human Brain. Neuron 2015, 87, 657–670. [Google Scholar] [CrossRef]
- Poldrack, R.A. Precision Neuroscience: Dense Sampling of Individual Brains. Neuron 2017, 95, 727–729. [Google Scholar] [CrossRef]
- Braga, R.M.; Buckner, R.L. Parallel Interdigitated Distributed Networks within the Individual Estimated by Intrinsic Functional Connectivity. Neuron 2017, 95, 457–471.e455. [Google Scholar] [CrossRef]
- Feilong, M.; Nastase, S.A.; Guntupalli, J.S.; Haxby, J.V. Reliable individual differences in fine-grained cortical functional architecture. NeuroImage 2018, 183, 375–386. [Google Scholar] [CrossRef]
- Gagnier, J.J.; Boon, H.; Rochon, P.; Moher, D.; Barnes, J.; Bombardier, C. Reporting Randomized, Controlled Trials of Herbal Interventions: An Elaborated CONSORT Statement. Ann. Int. Med. 2006, 144, 364–367. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Memory Scale, 4th ed.; Pearson Assessment: San Antonio, TX, USA, 2009. [Google Scholar]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Rovner, B.W.; Folstein, M.F. Mini-mental state exam in clinical practice. Hosp. Pract. 1987, 22, 140–141. [Google Scholar]
- Hughes, C.P.; Berg, L.; Danziger, W.L.; Coben, L.A.; Martin, R.L. A new clinical scale for the staging of dementia. Br. J. Psychiatry 1982, 140, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.A.; Walsh, C.; Blanco, A.; Moran, M.; Coen, R.F.; Walsh, J.B.; Lawlor, B.A. The clinical dementia rating sum of box score in mild dementia. Dement. Geriatr. Cogn. Disord. 2006, 21, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Santos-Buelga, C.; González-Paramás, A.M.; Oludemi, T.; Ayuda-Durán, B.; González-Manzano, S. Plant phenolics as functional food ingredients. Adv. Food Nutr. Res. 2019, 90, 183–257. [Google Scholar] [PubMed]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.B.; Hassan, S.; Waheed, M.; Javed, A.; Farooq, M.A.; Tahir, A. Bioavailability and Metabolic Pathway of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; Soto-Hernández, M., García-Mateos, R., Palma-Tenango, M., Eds.; InTechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Rubia, K.; Smith, A.B.; Brammer, M.J.; Taylor, E. Right inferior prefrontal cortex mediates response inhibition while mesial prefrontal cortex is responsible for error detection. Neuroimage 2003, 20, 351–358. [Google Scholar] [CrossRef]
- Braver, T.S.; Barch, D.M.; Gray, J.R.; Molfese, D.L.; Snyder, A. Anterior cingulate cortex and response conflict: Effects of frequency, inhibition and errors. Cereb Cortex 2001, 11, 825–836. [Google Scholar] [CrossRef]
- Rubia, K.; Russell, T.; Bullmore, E.T.; Soni, W.; Brammer, M.J.; Simmons, A.; Taylor, E.; Andrew, C.; Giampietro, V.; Sharma, T. An fMRI study of reduced left prefrontal activation in schizophrenia during normal inhibitory function. Schizophr. Res. 2001, 52, 47–55. [Google Scholar] [CrossRef]
- Shibata, T.; Shimoyama, I.; Ito, T.; Abla, D.; Iwasa, H.; Koseki, K.; Yamanouchi, N.; Sato, T.; Nakajima, Y. The time course of interhemispheric EEG coherence during a GO/NO-GO task in humans. Neurosci. Lett. 1997, 233, 117–120. [Google Scholar] [CrossRef]
- Roberts, L.E.; Rau, H.; Lutzenberger, W.; Birbaumer, N. Mapping P300 waves onto inhibition: Go/No-Go discrimination. Electroencephalogr. Clin. Neurophysiol. Evoked Potential. 1994, 92, 44–45. [Google Scholar] [CrossRef]
- Menon, V.; Adleman, N.E.; White, C.D.; Glover, G.H.; Reiss, A.L. Error-related brain activation during a Go/NoGo response inhibition task. Hum. Br. Mapp. 2001, 12, 131–143. [Google Scholar] [CrossRef]
- Swick, D.; Ashley, V.; Turken, U. Are the neural correlates of stopping and not going identical? Quantitative meta-analysis of two response inhibition tasks. NeuroImage 2011, 56, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, L.; Neri, F.; Momi, D.; Menardi, A.; Rossi, S.; Rossi, A.; Santarnecchi, E. Stimuli, presentation modality, and load-specific brain activity patterns during n-back task. Hum. Br. Mapp. 2019, 40, 3810–3831. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, W.; Wu, J.; Zhang, J.; Jin, Z.; Li, L. A coordinate-based meta-analysis of the n-back working memory paradigm using activation likelihood estimation. Br. Cogn. 2019, 132, 1–12. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.K.; Prior, R.L.; Huang, D. Novel Fluorometric Assay for Hydroxyl Radical Prevention Capacity Using Fluorescein as the Probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Chung, H.Y.; Choi, H.R.; Park, H.J.; Choi, J.S.; Choi, W.C. Peroxynitrite Scavenging and Cytoprotective Activity of 2,3,6-Tribromo-4,5-dihydroxybenzyl Methyl Ether from the Marine Alga Symphyocladia latiuscula. J. Agric. Food Chem. 2001, 49, 3614–3621. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, D.; Kondo, M.; Fan, E.; Ji, H.; Kou, Y.; Ou, B. Novel High-Throughput Assay for Antioxidant Capacity against Superoxide Anion. J. Agric. Food Chem. 2009, 57, 2661–2667. [Google Scholar] [CrossRef]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004, 23. [Google Scholar] [CrossRef]
- Jenkinson, M.; Bannister, P.R.; Brady, J.M.; Smith, S.M. Improved optimisation for the robust and accurate linear registration and motion correction of brain images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Smith, S.M. A global optimisation method for robust affine registration of brain images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Woolrich, M.W.; Ripley, B.D.; Ripley, J.M.; Smith, S.M. Temporal autocorrelation in univariate linear modeling of fMRI data. NeuroImage 2001, 14, 1370–1386. [Google Scholar] [CrossRef] [PubMed]
- Worsley, K.J. Statistical analysis of activation images. In Functional MRI: An Introduction to Methods; Jezzard, P., Matthews, P.M., Smith, S.M., Eds.; OUP: Cary, NC, USA, 2001. [Google Scholar]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Br. Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef]
- Statistical Parametric Mapping. Available online: http://www.fil.ion.ucl.ac.uk/spm (accessed on 3 January 2019).
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef]
- Kreis, R. The trouble with quality filtering based on relative Cramér-Rao lower bounds. Magn. Reson. Med. 2016, 75, 15–18. [Google Scholar] [CrossRef]
- Gasparovic, C.; Song, T.; Devier, D.; Bockholt, H.J.; Caprihan, A.; Mullins, P.G.; Posse, S.; Jung, R.E.; Morrison, L.A. Use of tissue water as a concentration reference for proton spectroscopic imaging. Magn. Reson. Med. 2006, 55, 1219–1226. [Google Scholar] [CrossRef]
- Han, X.; Shen, T.; Lou, H. Dietary Polyphenols and Their Biological Significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef]
- Henneberger, C.; Jüttner, R.; Rothe, T.; Grantyn, R. Postsynaptic Action of BDNF on GABAergic Synaptic Transmission in the Superficial Layers of the Mouse Superior Colliculus. J. Neurophysiol. 2002, 88, 595–603. [Google Scholar] [CrossRef]
- Newsholme, P.; Procopio, J.; Lima, M.M.; Pithon-Curi, T.C.; Curi, R. Glutamine and glutamate—Their central role in cell metabolism and function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [CrossRef]
- Watford, M. Glutamine and glutamate: Nonessential or essential amino acids? Anim. Nutr. 2015, 1, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Shen, J. Modeling the glutamate–glutamine neurotransmitter cycle. Front. Neuroenerg. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Strużyńska, L.; Sulkowski, G. Relationships between glutamine, glutamate, and GABA in nerve endings under Pb-toxicity conditions. J. Inorg. Biochem. 2004, 98, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Al-Otaish, H.; Al-Ayadhi, L.; Bjørklund, G.; Chirumbolo, S.; Urbina, M.A.; El-Ansary, A. Relationship between absolute and relative ratios of glutamate, glutamine and GABA and severity of autism spectrum disorder. Metab. Br. Dis. 2018, 33, 843–854. [Google Scholar] [CrossRef]
- Huang, D.; Liu, D.; Yin, J.; Qian, T.; Shrestha, S.; Ni, H. Glutamate-glutamine and GABA in brain of normal aged and patients with cognitive impairment. Eur. Radiol. 2017, 27, 2698–2705. [Google Scholar] [CrossRef]
- Liguz-Lecznar, M.; Lehner, M.; Kaliszewska, A.; Zakrzewska, R.; Sobolewska, A.; Kossut, M. Altered glutamate/GABA equilibrium in aged mice cortex influences cortical plasticity. Br. Struct. Funct. 2015, 220, 1681–1693. [Google Scholar] [CrossRef]
- Niu, X.; Zheng, S.; Liu, H.; Li, S. Protective effects of taurine against inflammation, apoptosis, and oxidative stress in brain injury. Mol. Med. Rep. 2018, 18, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Seidel, U.; Huebbe, P.; Rimbach, G. Taurine: A Regulator of Cellular Redox Homeostasis and Skeletal Muscle Function. Mol. Nutr. Food Res. 2018, 63, 1800569. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenol | mg/g |
|---|---|
| 3-O-Caffeoylquinic acid | 41.3 ± 8.4 |
| 5-p-coumaroylquinic acid | 0.4 ± 0.1 |
| 5-O-Caffeoylquinic acid | 134.6 ± 16.7 |
| 3-Feruloylquinic acid | 7 ± 1.2 |
| 4-O-Caffeoylquinic acid | 74.9 ± 11.1 |
| p-coumaroylquinic acid | 0.6 ± 0.1 |
| p-coumaroylquinic acid | 2.5 ± 0.5 |
| 4-Feruloylquinic acid | 8.4 ± 1.7 |
| 5-Feruloylquinic acid | 38.1 ± 6.3 |
| 3-O-Caffeoylquinic lactone | 7.8 ± 1.2 |
| 4-O-Caffeoylquinic lactone | 3.8 ± 0.6 |
| 3,4-O-Dicaffeoylquinic acid | 37.6 ± 7.3 |
| 3,5-O-Dicaffeoylquinic acid | 12.1 ± 2.2 |
| 4,5-O-Dicaffeoylquinic acid | 47.2 ± 7.8 |
| 3-O-Feruloyl-4-Caffeoylquinic acid | 0.9 ± 0.2 |
| 3-O-Caffeoy-4-Feruloylquinic acid | 3.3 ± 0.6 |
| 3-O-Feruloyl-5-Caffeoylquinic acid | 0.2 ± 0 |
| 3-O-Caffeoy-5-Feruloylquinic acid | 1.1 ± 0.3 |
| 4-O-Feruloyl-5-Caffeoylquinic acid | 0.7 ± 0.2 |
| 4-O-Caffeoy-5-Feruloylquinic acid | 3.4 ± 0.7 |
| Total CGA | 425.8 ± 63.9 |
| Trigonelline | 33.78 ± 5.2 |
| Caffeine | 18.2 ± 3.3 |
| Exosomal BDNF Results | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Descriptives | t-Statistics | |||||||||||||
| Pre | Post | 95% Confidence Interval of the Difference | ||||||||||||
| M | SD | SEM | M | SD | SEM | M | SD | SEM | Lower | Upper | t | df | Sig. (1-Tailed) | |
| Post WCCE> Pre WCCE | 17.92 | 6.17 | 2.18 | 25.31 | 9.26 | 3.27 | 7.40 | 10.45 | 3.69 | −1.34 | 16.13 | 2.00 | 7 | 0.04 |
| Post Placebo > Pre Placebo | 18.03 | 2.95 | 1.04 | 14.47 | 7.50 | 2.65 | −3.56 | 7.08 | 2.50 | −9.48 | 2.36 | −1.42 | 7 | 0.10 |
| N-back Behavioral Results | |||||||||||||||
| Descriptives | t-Statistics | ||||||||||||||
| Pre | Post | 95% Confidence Interval of the Difference | |||||||||||||
| M | SD | SEM | M | SD | SEM | M | SD | SEM | Lower | Upper | t | df | Sig. (1-Tailed) | ||
| Post WCCE > Pre WCCE | Number of Errors | 2.125 | 2.532 | 0.895 | 2.625 | 2.774 | 0.981 | 0.500 | 2.070 | 0.732 | −1.231 | 2.231 | 0.683 | 7 | 0.258 |
| Reaction Time for Correct Trials | 574.670 | 84.335 | 29.817 | 536.527 | 82.185 | 29.057 | −38.144 | 29.567 | 10.454 | −62.863 | −13.425 | −3.649 | 7 | 0.004 | |
| Post Placebo > Pre Placebo | Number of Errors | 3.250 | 3.770 | 1.333 | 3.000 | 2.828 | 1.000 | −0.250 | 1.389 | 0.491 | −1.411 | 0.911 | −0.509 | 7 | 0.313 |
| Reaction Time for Correct Trials | 568.838 | 111.814 | 39.532 | 559.269 | 86.193 | 30.474 | −9.569 | 66.678 | 23.574 | −65.314 | 46.175 | −0.406 | 7 | 0.349 | |
| Go/No-Go Behavioral Results | |||||||||||||||
| Descriptives | t-Statistics | ||||||||||||||
| Pre | Post | 95% Confidence Interval of the Difference | |||||||||||||
| M | SD | SEM | M | SD | SEM | M | SD | SEM | Lower | Upper | t | df | Sig. (1-Tailed) | ||
| Post WCCE > Pre WCCE | Reaction Time for Successful Trials | 387.953 | 38.729 | 15.811 | 381.654 | 44.676 | 18.239 | −6.299 | 23.166 | 9.457 | −30.610 | 18.011 | −0.666 | 5 | 0.2675 |
| Accuracy | 242.000 | 5.967 | 2.436 | 242.333 | 4.082 | 1.667 | 0.333 | 2.805 | 1.145 | −2.610 | 3.277 | 0.291 | 5 | 0.3915 | |
| Post Placebo > Pre Placebo | Reaction Time for Successful Trials | 397.013 | 26.611 | 9.408 | 395.563 | 41.291 | 14.599 | −1.450 | 25.287 | 8.940 | −22.591 | 19.691 | −0.162 | 7 | 0.438 |
| Accuracy | 242.125 | 5.463 | 1.931 | 239.625 | 7.170 | 2.535 | −2.500 | 2.563 | 0.906 | −4.643 | −0.357 | −2.758 | 7 | 0.014 | |
| Antioxidant Assay | µmol Trolox Equivalent/g |
|---|---|
| ORAC | 6097 ± 225 |
| HORAC | 18,709 ± 426 |
| NORAC | 527 ± 52 |
| SORAC | 860 ± 24 |
| SOAC | 2042 ± 185 |
| Total ORAC | 28,237 ± 782 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, J.L.; Yanes, J.A.; Reid, M.A.; Murphy, J.E.; Busler, J.N.; Mumford, P.W.; Young, K.C.; Pietrzkowski, Z.J.; Nemzer, B.V.; Hunter, J.M.; et al. Neurophysiological Effects of Whole Coffee Cherry Extract in Older Adults with Subjective Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled, Cross-Over Pilot Study. Antioxidants 2021, 10, 144. https://doi.org/10.3390/antiox10020144
Robinson JL, Yanes JA, Reid MA, Murphy JE, Busler JN, Mumford PW, Young KC, Pietrzkowski ZJ, Nemzer BV, Hunter JM, et al. Neurophysiological Effects of Whole Coffee Cherry Extract in Older Adults with Subjective Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled, Cross-Over Pilot Study. Antioxidants. 2021; 10(2):144. https://doi.org/10.3390/antiox10020144
Chicago/Turabian StyleRobinson, Jennifer L., Julio A. Yanes, Meredith A. Reid, Jerry E. Murphy, Jessica N. Busler, Petey W. Mumford, Kaelin C. Young, Zbigniew J. Pietrzkowski, Boris V. Nemzer, John M. Hunter, and et al. 2021. "Neurophysiological Effects of Whole Coffee Cherry Extract in Older Adults with Subjective Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled, Cross-Over Pilot Study" Antioxidants 10, no. 2: 144. https://doi.org/10.3390/antiox10020144
APA StyleRobinson, J. L., Yanes, J. A., Reid, M. A., Murphy, J. E., Busler, J. N., Mumford, P. W., Young, K. C., Pietrzkowski, Z. J., Nemzer, B. V., Hunter, J. M., & Beck, D. T. (2021). Neurophysiological Effects of Whole Coffee Cherry Extract in Older Adults with Subjective Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled, Cross-Over Pilot Study. Antioxidants, 10(2), 144. https://doi.org/10.3390/antiox10020144