
Hydrogen Peroxide Increases during Endodormancy and Decreases during Budbreak in Grapevine (Vitis vinifera L.) Buds


Laboratorio de Bioquimica Vegetal, Facultad de Ciencias, Universidad de Chile, Casilla 653, Las Palmeras, Ñuñoa 3425, Chile
*
Author to whom correspondence should be addressed.
Antioxidants 2021, 10(6), 873; https://doi.org/10.3390/antiox10060873
Submission received: 26 April 2021
/
Revised: 24 May 2021
/
Accepted: 25 May 2021
/
Published: 29 May 2021
(This article belongs to the Special Issue Antioxidant Networks in Seed and Bud Dormancy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Changes in the level of hydrogen peroxide (H2O2) is a good indicator to monitor fluctuations in cellular metabolism and in the stress responses. In this study, the changes in H2O2 content during bud endodormancy (ED) and budbreak were analysed in grapevine (Vitis vinifera L.). The results showed a gradual increase in the H2O2 content during the development of bud ED, which was mainly due to an increase in the activity of peroxidases (PODs). The maximum H2O2 content reached in the grapevine buds coincided with the maximum depth of bud ED. In contrast, during budbreak, the H2O2 content decreased. As the plant hormones cytokinin (CK) and auxin play an important role in budbreak and growth resumption in grapevine, the effect of exogenous applications of H2O2 on the expression of genes involved in CK and auxin metabolism was analysed. The results showed that H2O2 represses the expression of the CK biosynthesis genes VvIPT3a and VvLOG1 and induces the expression of the CK-inactivating gene VvCKX3, thus reducing potentially the CK content in the grapevine bud. On the other hand, H2O2 induced the expression of the auxin biosynthesis genes VvAMI1 and VvYUC3 and of the auxin transporter gene VvPIN3, thus increasing potentially the auxin content and auxin transport in grapevine buds. In general, the results suggest that H2O2 in grapevine buds is associated with the depth of ED and negatively regulates its budbreak.
1. Introduction
The buds of the grapevine (Vitis vinifera L.), similar to the buds of other deciduous fruit trees enter a state of winter recess or endodormancy (ED) to survive winter conditions. In the grapevine bud, ED is induced by the shortening of the photoperiod [1,2,3] and is regulated by endogenous factors that inhibit its growth and development [4]. The plant hormone abscisic acid (ABA) accumulates in the buds of grapevines throughout ED [5,6] and plays a key role in its maintenance and release [7,8,9]. On the other hand, hydrogen peroxide (H2O2), which is continuously generated from various sources during normal metabolism and is a signalling molecule that mediates responses to biotic and abiotic stresses [10], also accumulates in the buds of grapevine during ED [11,12,13]. Further, H2O2 has been suggested to play a pivotal role in the control of ED and budbreak [14]. H2O2 can also be generated by specific enzymes such as the respiratory burst oxidase homologs (RBOH), xanthine oxidase, amine oxidase and cell wall peroxidase [10]. Generally, the plant antioxidant system removes H2O2 efficiently, and thus the oxidative effects of many stimuli could be mediated via a reduction in the activities of antioxidant enzymes, rather than by increased H2O2 generation [10]. In grapevine buds, hypoxia, as well as mitochondrial respiration inhibitors, such as potassium cyanide (KCN) and sodium nitroprusside (SNP), increase H2O2 levels [15]. In Arabidopsis thaliana, hypoxia was found to activate Rop-GTPase signalling, which in turn activated RBOH [16]. In grapevines, seven VvRBOH genes have been identified and characterised [17], and three of them are expressed in the buds [18]. Peroxidase class III (POD) represents a class of ubiquitous enzymes widely distributed in plants whose primary function is to oxidize molecules at the expense of H2O2. POD is therefore generally considered an H2O2 detoxifying enzyme; however, POD can also produce H2O2. In grapevines and peach, it has been reported that POD can generate H2O2 through the oxidation of NADH, and this reaction is catalysed by p-coumaric acid [11,19]. Recently, a subfamily of 47 VvPOD genes was identified in the grapevine genome [20] and 30 subfamily members were expressed in the bud [18]. Oxidative stress and reactive oxygen species (ROS) have been suggested to play a central role in the release of buds from ED in grapevines [14]. Interestingly, most studies using chemical and physical dormancy breakdown stimuli such as hydrogen cyanamide (HC), sodium azide (AZ) and heat shock (HS) [11,12,13] show a transient increase in H2O2 levels within the grapevine bud, which has been interpreted as a positive signal for the release of buds from the ED [13,14].
In this study, the changes in H2O2 levels though ED and budbreak were analysed in grapevine buds. Additionally, the effects of exogenous applications of H2O2 on the expression of cytokinin (CK) and auxin-related genes in grapevine buds were examined. From the results, we found that the H2O2 level is associated with the degree of ED and is a negative regulator of budbreak in grapevine buds.
2. Materials and Methods
2.1. Plant Material
Plant material was collected from 8-year-old (Vitis vinifera L. cv. Thompson Seedless) vineyards growing at the experimental station of the Chilean National Institute of Agriculture Research (INIA) located in Santiago, Chile (33◦34′ S). The grapevine plants were watered by immersion and trained in an aerial training system. Six whole canes were randomly collected every 3 weeks starting at the beginning of January (early summer) and ending at the end of August (early spring). The collected canes were cut at both ends, leaving a central section of 10–12 buds for experimental use.
2.2. Dormancy Depth
The budbreak response of single-bud cuttings under forced conditions is widely used to describe the depth of dormancy in grapevine [21,22]. Thirty cuttings were mounted on polypropylene sheets and placed in a container with water and transferred to a growth chamber set at 23 ± 2 °C with 16 h (h) light. The budbreak was assessed every 5 days (d) during a period of 30 d, the appearance of a green tip was the signal that indicated the beginning of the budbreak. This procedure was repeated for the different collection dates. The time required for each sample to reach budbreak, including right-censored observations of the buds that did not break during the treatment, was adjusted to the survival distribution function by the nonparametric Kaplan–Meier method [23]. The first sample of the year collected in early January before the onset of ED [24,25] was used as a reference of the behaviour of budbreak, when growth was not restricted within the buds (paradormant buds). A log-rank test was performed to compare the estimated survival distributions of the reference sample with samples collected at other dates. The greater the difference in the chi-square between the analysed sample and the reference sample, the greater the degree of ED [23].
2.3. Chemical Treatments
Single-node cuttings collected on June 11 were painted with 2.5% (w/v) hydrogen cyanamide (HC) (Dormex, SKW, Trotsberg, Germany), 2% (w/v) 3-amino-1,2,4-triazole (Sigma, Burlington, VT, USA) or water, as a control. Cuttings were mounted on a polypropylene sheet and placed in a container with water and transferred to the growth chamber set at 23 ± 2 °C under 14 h light forced conditions (FC); samples were removed at each specific point. Experiments inducing the breakage of ED were carried out with buds collected on July 18 and treated with 2.5% (w/v) HC, 2% (w/v) 3-amino-1,2,4-triazole or water as a control. After treatments, cuttings were mounted as described above and settled in the growth chamber under FC and the breakage of buds was assessed every 2 d.
2.4. H2O2 Measurements
The H2O2 concentration was measured by chemiluminescence (CL) based on a cobalt-catalysed oxidation of luminol (5-amino-2,3-dihydro-1,4-phtalazinedione) [26]. Three different buds were analysed for each sampling date and values corresponded to the average of the three samples.
2.5. Peroxidase and Catalase Activity
Buds were ground in liquid nitrogen, and the resulting powder was extracted with a buffer containing 0.5 M Tris-HCl, 5 mM DTT, 1 mM MgCl2, 10 µM PMSF, 2% insoluble PVP and 12.5% glycerol (pH 7.5). Peroxidase (donor: hydrogen peroxide oxidoreductase EC1.11.1.7) activity was assayed according to [27] by measuring the H2O2-dependent oxidation of o-phenylendiamine (o-PDA) via spectrophotometry at 450 nanometers (nm) in a mixture with pH 4.5 containing 0. 1 M sodium citrate, 44 mM o-PDA, 1 mM H2O2 and 5–10 µL of the extract. Catalase activity was determined following O2 evolution using a Clark-type oxygen electrode (Hansatech, UK) as according to the methodology of [12].
2.6. RNA Purification, and cDNA Synthesis
Total RNA was isolated and purified from dormant buds (0.5 g FW) of Thompson seedless. Total RNA was extracted and purified using the method of Chang et al. [28] modified according to Noriega et al. [29]. The DNA was removed from the sample with RNAse-free DNase I (1 U/µg) (Thermo Scientific, Bedford, MA, USA) at 37 °C for 30 min. The first cDNA strand was synthesised from 1.0 µg of purified RNA with 1 µL of oligo (dT) 12–18 (0.5 µg × µL−1) as a primer, 1 µL of dNTP mixture (10 mM) and Superscript® II RT (Invitrogen, CA, USA).
2.7. Gene Expression Analysis
The expression analysis of the genes was performed by quantitative real-time PCR (RT-qPCR) using an Eco Real-Time PCR system (Illumina, Inc. San Diego, CA, USA) and KAPA SYBR FAST (KK 4602) qPCR Master Mix (2×). Sequences of grapevine genes were obtained from the grape genomic database (www.genoscope.fr, accessed on 24 May 2021). Primers for amplification were designed using PRIMER3 software [30]. The cDNA was amplified under the following conditions: denaturation at 94 °C for 2 min and 40 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 45 s. Relative changes in gene expression levels were determined using the 2−ΔΔCT method [31]. Each reaction was performed with at least two biological replicates, each with three technical replicates. VvUBIQUITIN (GSVIVT01038617001) and VvACTIN (GSVIVT01026580001) were used as reference genes for normalisation.
2.8. Data Analysis
Survival analysis, or the time to event analysis, is a category of statistical methods designed specifically to handle a response variable that measures the elapsed time until a specific event occurs (here, budbreak), which may be censored [32]. Kaplan–Meier (KM) survival curves are the simplest way to estimate survival over time when data are censored [23]. The percentage or probability of absence of budbreak is calculated as a function of time after sampling according to the KM method. However, in this study, we used the complement of the probability of the absence of budbreak, which corresponds to the probability of budbreak [23]. A log-rank test was carried out to compare the estimated survival distribution of the reference sample against the other samples. A significant difference indicates differences in the budbreak distribution curves, and the larger the value of the chi-square (χ2) is, the greater the difference between the reference and the other sample, thus the greater the ED depth of the sample.
3. Results
3.1. Depth of Endodormancy and H2O2 Content in Grapevine Buds
The survival analysis of the budbreak of grapevine buds under forced conditions was carried out on the different collection dates using the nonparametric Kaplan–Meier method [23]. The comparison of the KM probabilistic function between the buds collected before (reference) and after the onset of the ED was performed using a log-rank test. The greater the chi-square between the reference and the samples, the greater the degree of ED [23]. Using this methodology, two phases were distinguished during ED in Thompson seedless grapevines grown in Santiago, Chile. The first phase began in mid-January and lasted until mid-April, and the second phase peaked in late May and ended in late August. During the second phase of the ED, a sustained increase in the H2O2 content was observed, whose maximum coincided with the maximum depth of the ED (Figure 1). This dormancy pattern is consistent with the transcriptome changes observed during bud development in grapevines [18].
3.2. Increases in Peroxidase Activity and H2O2 Content Coincided through Endodormancy in the Grapevine Buds
In grapevine, it has been reported that peroxidases (PODs) can generate H2O2 by oxidation of NADH, and this reaction is catalysed by p-coumaric acid [11]. Here, we analysed the POD activity throughout the ED period in Thompson seedless grapevine buds, and this activity was compared with the evolution of the H2O2 content in the buds (Figure 2a). The results showed that the maximum POD activity coincided with the maximum H2O2 content, suggesting that the POD activity may be at least partially responsible for the increases in the H2O2 content in grapevine buds. The increase in POD activity is probably due to an increase in protein abundance, since the expression of VvPODs genes increased (see Section 3.3).
3.3. Expression Profile of VvPOD Genes throughout the Endodormancy in Grapevine Buds
A total of 47 VvPOD genes have been identified in the V. vinifera genome [20], and a phylogenetic tree revealed that VvPODs showed a relatively close genetic relationship with similar genes from Arabidopsis [20]. Of this large number of VvPODs, 30 are expressed in the bud [18], and nine change their expression level during the development of ED. The transcript levels of VvPODs taken from microarray data on the development of Tempranillo grape buds grown in the Northern Hemisphere [18] were plotted throughout the bud growth period. The results showed that when the data were adapted to the conditions of the Southern Hemisphere (Figure 2b), only the expression profile of VvPOD38 was consistent with the increase in POD activity during the development of ED, and VvPOD25 was expressed slightly during ED but increased before budbreak (Figure 2b).
3.4. Expression Profile of RBOH throughout the Endodormancy Period in Grapevine Buds
As one of the major sources of ROS in plants is the NADPH oxidase-catalysed conversion of oxygen to O2−, the expression of the RBOH genes in grapevine buds was analysed. Seven VvRBOH genes have been identified in the V. vinifera genome [17], and three of them are expressed in the bud [18]. The transcript levels of VvRBOHs taken from microarray data on the development of Tempranillo grape buds grown in the Northern Hemisphere [18] were plotted throughout the ED period. The results showed that when the data were adapted to the conditions of the Southern Hemisphere (Figure 3), the expression of VvRBOHE was the only family member to have its transcriptional activity significantly altered during the development of ED; however, its maximum expression level did not match the maximum level of H2O2.
3.5. Scavenging Activity of H2O2 during Endodormancy in Grapevine Buds
It is well known that the ROS scavenging system is involved in the control of H2O2 in many plant developmental processes [33]. Here, we analysed the expression profile of the catalase genes VvCAT1 and VvCAT2 and catalase activity throughout the ED period in grapevine buds. The transcript levels of VvCAT1 and VvCAT2 taken from microarray data on the development of Tempranillo grape buds grown in the Northern Hemisphere [18] were plotted throughout the ED period. The results showed that when the data were adapted to the conditions of the Southern Hemisphere (Figure 4b), no major variations in the expression of VvCAT1 and VvCAT2 in the buds during their ED period were observed. Catalase activity also did not vary significantly during the ED period in Thompson seedless grapevines, but a peak was detected at the end of July (Figure 4a). As there is a correspondence between catalase activity and expression of catalase genes, the changes in activity reflect the changes in protein abundance.
On the other hand, aminotriazole, an inhibitor of catalase activity [34], rapidly increased H2O2 content in grapevine buds collected at the end of the ED, and this high H2O2 level was maintained for longer than when buds were treated with HC (Figure 5a). Furthermore, aminotriazole delayed the sprouting of grapevine buds relative to control buds (Figure 5b).
3.6. Variations in the H2O2 Content during Budbreak in Grapevine Buds under Forced Conditions
To evaluate the changes in the H2O2 content during forcing budbreak, the H2O2 content was measured several days after the onset of the treatment in grapevine buds collected in early June, when they were endodormant [24] (Figure 6a). The H2O2 was also measured a few hours after the onset of the treatment in grapevine buds collected at the end of July, once the ED was finished and the buds were quiescent [24] (Figure 6b). The H2O2 content decreased rapidly in the quiescent buds (Figure 5b), while in the dormant buds the decrease in the H2O2 content was much slower (Figure 5a).
3.7. Cytokinin-Inactivating Genes and Auxin Biosynthesis Genes Are Induced by H2O2 in Grapevine Buds, While Cytokinin Biosynthesis Genes Are Repressed
Cytokinin has an antagonistic function to auxin in different developmental processes [35]. Here, we analysed the effect of exogenous applications of H2O2 on the expression of the CK biosynthesis genes, VvIPT3a and VvLOG1, and the CK inactivating gene VvCKX3 in grapevine buds. We selected these genes of CK metabolism because their expression varied greatly during the sprouting of grapevine buds under forced conditions [36]. Additionally, the effect of H2O2 on the expression of the auxin biosynthesis genes, VvAMI1 and VvYUC3, and the auxin transport gene VvPIN3 was also analysed for the same reason mentioned above. The results showed that after 48 h of H2O2 treatment, the expression levels of the CK biosynthesis genes, VvIPT3a and VvLOG1 decreased, while that of the CK-inactivating gene VvCKX3 increased (Figure 7a). These results suggest that H2O2 decreases the CK content in the grapevine buds. On the other hand, H2O2 increased the expression levels of the auxin biosynthesis genes VvAMI1 and VvYUC3 and of the auxin transport gene VvPIN3 (Figure 7b) in grapevine buds. These results suggest that the auxin content and auxin transport are increased in grapevine buds after H2O2 treatment.
4. Discussion
4.1. ABA and H2O2 Accumulate in Grapevine Buds during ED
Under stress conditions, abscisic acid (ABA) and hydrogen peroxide (H2O2) accumulate in many plant biological systems [37,38]. In grapevine buds, both molecules accumulate during ED [5,6]. The ABA peak occurred before that of H2O2 [6] and the peak of H2O2 coincided with the maximum depth of the ED (Figure 1), which suggests that ABA could control the level of H2O2, which in turn, would be related to the depth of the ED in grapevine buds. In guard cells, ABA induces H2O2 production presumably via the activity of NADPH-oxidase [39]. In rice roots, ABA induces increases in H2O2 via activation of cell wall peroxidases [40]. In rice leaves under water stress, ABA controls H2O2 accumulation through the induction of the catalase gene OsCATB [41], and grafting-induced ABA accumulation in cucumber leaves triggers H2O2 production and enhances the activities of antioxidant enzymes [42]. In grapevine, during bud ED, H2O2 levels correlated with increases in POD activity and VvPOD38 gene expression, but not with increases in VvRBOH and VvCAT gene expression, which suggested that during bud ED, the H2O2 content is controlled mainly by POD activity. Interestingly, ABA has been reported to regulate the expression of several VvPOD genes in grapevine berry skins (Table S1) including VvPOD38 [43,44], but its effect on H2O2 levels has not been described in grapevines. However, the role of ABA in bud ED and budbreak in grapevines has been extensively reported in the literature [7,8,9]. Thus, it has been shown that ABA represses the expression of cell cycle genes in the bud meristem of grapevine [8], and its removal is a key step for dormancy release [7,9]. Additionally, ABA promotes starch synthesis and storage metabolism in endodormant grapevine buds, thus contributing to the establishment of bud ED [6]. On the other hand, although H2O2 accumulates in grapevine buds during ED, its role in dormancy regulation has been associated mainly with the release of buds from ED rather than with their establishment and maintenance.
4.2. The Role of H2O2 during Bud ED Release and Budbreak
As the application of chemical and physical stimuli that break bud dormancy, such as hydrogen cyanamide (HC), sodium azide (AZ), heat shock (HS) and hypoxia, induces a transient increase in H2O2 levels [15,45,46], the release of buds from ED has been associated with oxidative and respiratory stress [14]. Recently, it has been demonstrated that HC transiently increases H2O2 levels and the expression of the VvPOD72 (GSVIVT01029771001) gene in paradormant grapevine buds [45], and in a proteomic study, it was demonstrated that HC induces the expression of VvPOD25 [19], a gene that is weakly expressed during ED and more highly expressed during budbreak. All these studies suggest that the transient increase in H2O2 levels observed in grapevine buds after being treated with HC is due either to an increase in the expression of VvPOD genes or to the inhibition of catalase activity [12,13]. The transient increase in H2O2 levels could act as a secondary messenger, triggering the expression of genes related to endodormancy release [13]. However, when H2O2 levels remained high for a long period of time, such as after aminotriazole treatment, the budbreak was delayed (Figure 7). In rose flower buds, the content of H2O2 decreases during the outgrowth process as a result of the activation of the ascorbic acid-glutathione (ASA-GSH) cycle [47]. Additionally, exogenous applications of H2O2 to rose buds delay their outgrowth and inhibit the expression of key genes involved in bud outgrowth, such as VACUOLAR INVERTASE (RhVI), EXPANSIN (RhEXP), PROLIFERATING CELL NUCLEAR ANTIGEN (RhPCNA) and CYCLIN D3 (RHCYCD3) [47]. These results indicate that H2O2 negatively controls the outgrowth of rose buds. Our results on the budbreak of quiescent grapevine buds under forced conditions showed a rapid drop in the H2O2 content with budbreak progress, while in dormant buds, this drop was slower, indicating that H2O2 should be removed from the bud by the antioxidative system before budbreak and the resumption of growth begins. In addition, the fact that exogenous applications of H2O2 reduce the CK content by inducing the expression of the CK degrading gene (VvCKX3) and repressing the expression of the CK biosynthesis genes (VvIPT3a and VvLOG1) suggests that H2O2 negatively affects budbreak, since CK positively modulates cell division and proliferation in meristematic tissue [48]. One of the first cellular events that occurs before the onset of budbreak in grapevine buds is the increase in the expression of CK biosynthesis genes as well as for genes related to the cell cycle [49,50]. In tomato plants, H2O2 promotes auxin biosynthesis in the apex, which, in turn, inhibits CK biosynthesis and subsequent bud outgrowth [51]. This result agrees with our results obtained in the grapevine buds which indicates an increase in the expression of auxin biosynthesis and transport genes after H2O2 treatments (Figure 6b).
5. Conclusions
During the development of ED in grapevine buds, the level of H2O2 increased as a result of a higher expression of the VvPODs genes, whose transcription could be regulated by ABA. On the contrary, during budbreak, the H2O2 level decreased. This difference, in changes in the level of H2O2 during bud development, is consistent with the transcriptional effect of H2O2 on the genes that regulate CK and auxin metabolism.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/antiox10060873/s1, Table S1: V. vinifera peroxidase genes (VvPODs) whose expression is regulated by ABA.
Author Contributions
Conceptualization F.J.P.; Methodology X.N., S.R., F.J.P.; Software S.R., X.N., F.J.P., Formal analysis, X.N., S.R., F.J.P.; Investigation X.N., S.R., F.J.P.; Resources F.J.P.; Writing F.J.P., Administration F.J.P.; Funding acquisition F.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funding by Fondecyt project #1190057.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kühn, N.; Ormeño-Nuñez, J.; Jaque-Zamora, G.; Pérez, F.J. Photoperiod modifies the diurnal expression profile of VvPHYA and VvPHYB transcripts in field-grown grapevine leaves. J. Plant Physiol. 2009, 66, 1172–1180. [Google Scholar] [CrossRef]
- Grant, T.N.L.; Gargrave, J.; Dami, I.E. Morphological physiological, and biochemical changes in Vitis genotype in responses to photoperiod regimes. Am. J. Enol. Vitic. 2013, 64, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Cragin, J.; Serpe, M.; Keller, M.; Shellie, K. Dormancy and cold hardiness transition in wine grape cultivars Chardonnay and Cabernet Sauvignon. Am. J. Enol. Vitic. 2017, 68, 195–202. [Google Scholar] [CrossRef]
- Lang, G.A. Dormancy: A new universal terminology. HortScience 1987, 22, 817–820. [Google Scholar]
- Or, E.; Belausov, E.; Popilevski, I.; Tal, Y.B. Changes in endogenous ABA level in relation to the dormancy cycle in grapevines grown in a hot climate. J. Hortic. Sci. Biotechnol. 2000, 75, 190–194. [Google Scholar] [CrossRef]
- Rubio, S.; Noriega, X.; Pérez, F.J. ABA promotes starch synthesis and storage metabolism in dormant grapevine buds. J. Plant Physiol. 2019, 234, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.H.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef] [PubMed]
- Vergara, R.; Noriega, X.; Pérez, F.J. ABA represses the expression of cell cycle genes an may modulate the development of endodormancy in grapevine buds. Front. Plant Sci. 2017, 8, 812. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Acheampog, A.K.; Shi, Z.; Mugzech, A.; Halaly-Basha, T.; Shaya, F.; Sun, Y.; Colova, V.; Mosquna, A.; Ophir, R.; et al. Abscisic acid catabolism enhances dormancy release of grapevine buds. Plant Cell Environ. 2018, 1, 14. [Google Scholar] [CrossRef]
- Neil, S.; Desikan, R.; Hancock, J. Hydrogen Peroxide signaling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Pérez, F.J.; Burgos, B. Alterations in the pattern of peroxidase isoenzymes and transient increases in its activity and in H2O2 levels take place during the dormancy cycle of grapevine buds: The effect of hydrogen cyanamide. Plant Growth Regul. 2004, 43, 213–220. [Google Scholar] [CrossRef]
- Pérez, F.J.; Lira, W. Possible role of catalase in post-dormancy bud break in grapevines. J. Plant Physiol. 2005, 162, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Pérez, F.J.; Vergara, R.; Rubio, S. H2O2 is involved in the dormancy-breaking effect of hydrogen cyanamide in grapevine buds. Plant Growth Regul. 2008, 55, 149–515. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud dormancy in perennial fruit tree species: A pivotal role for oxidative cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- Vergara, R.; Parada, F.; Rubio, S.; Pérez, F.J. Hypoxia induces H2O2 production and activates antioxidant defense system in grapevine buds through mediation of H2O2 and ethylene. J. Exp. Bot. 2012, 63, 4123–4131. [Google Scholar] [CrossRef] [Green Version]
- Baxler-Burrel, A.; Yang, Z.; Springer, P.S.; Bailey-Seres, J. RopGAP4-dependent Rop-GTPase rheostat control of Arabidopsis oxygen de privation tolerance. Science 2002, 296, 2026–2028. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Xu, X.; Gao, M.; Li, J.; Guo, C.; Song, J.; Wang, X. Genome-Wide analysis of respiratory burst oxidase homologs in grape (Vitis vinifera L.). Int. J. Mol. Sci. 2013, 14, 24169–24186. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Riquelme, J.; Grimplet, J.; Martínez-Zapater, J.M.; Carmona, M.J. Transcriptome variation along bud development in grapevine. BMC Plant Biol. 2012, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Khalil-Ur-Rehman, M.; Wang, W.; Dong, Y.; Faheem, M.; Xu, Y.; Gao, Z.; Tao, J. Comparative transcriptomic and proteomic analysis to deeply investigate the role of hydrogen cyanamide in grape bud dormancy. Int. J. Mol. Sci. 2019, 20, 3528. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Wang, C.; Khan, N.; Chen, M.; Fu, W.; Guan, L.; Leng, X. Genome-wide identification of class III POD gene family and their expression profiling in grapevine (Vitis vinifera L.). BMC Genom. 2020, 21, 444. [Google Scholar] [CrossRef]
- Koussa, T.; Broquedis, M.; Bouard, J. Changes of abscisic acid during the development of latent buds particularly in the phase of dormancy break. Vitis 1994, 33, 63–67. [Google Scholar]
- Dennis, G. Problems in standardizing methods for evaluating the chilling requirements for the breaking of dormancy in buds of woody plants. HortScience 2003, 38, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Camargo Alvarez, H.; Salazar-Gutierrez, M.; Zapata, D.; Keller, M.; Hoogenboom, G. Time to event analysis to evaluate dormancy status of single-bud cuttings: An example for grapevines. Plant Methods 2018, 14, 94. [Google Scholar] [CrossRef]
- Vergara, R.; Pérez, F.J. Similarities between natural and chemically induced bud-endodormancy release in grapevine Vitis vinifera L. Sci. Hortic. 2010, 125, 648–653. [Google Scholar] [CrossRef]
- Rubio, S.; Dantas, D.; Bressan-Smith, R.; Pérez, F.J. Relationship between endodormancy and cold hardiness in grapevine buds. J. Plant Growth Regul. 2016, 35, 266–275. [Google Scholar] [CrossRef]
- Pérez, F.J.; Rubio, S. An improved chemiluminescence method for hydrogen peroxide determinations in plant tissues. Plant Growth Regul. 2006, 48, 89–95. [Google Scholar] [CrossRef]
- Pérez, F.J.; Gómez, M. Gibberelic acid stimulation of isoperoxidase from pedicel of grape. Phytochemistry 1998, 48, 411–414. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Noriega, X.; Burgos, B.; Pérez, F.J. Short-day photoperiod triggers and low temperatures increase expression of peroxidase RNA transcripts and basic peroxidase isoenzyme activity in grapevine-buds. Phytochemitry 2007, 68, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the www for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Time to event survival data. BMJ 1998, 317, 468–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, P. The reaction between aminotriazole and catalase. Biochem. Biophys. Acta 1962, 59, 414–420. [Google Scholar] [CrossRef]
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin–auxin crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef]
- Noriega, X.; Pérez, F.J. ABA biosynthesis genes are down-regulated while auxin and cytokinin biosynthesis gene are up-regulated during the release of grapevine from endodormancy. J. Plant Growth Regul. 2017, 36, 814–823. [Google Scholar] [CrossRef]
- Assmann, S.M.; Shimazaki, K.I. The multisensory guard cell, stomatal responses to blue light and abscisic acid. Plant Physiol. 1999, 119, 809–815. [Google Scholar] [CrossRef] [Green Version]
- Potikha, T.S.; Collins, C.C.; Johnson, D.I.; Delmer, D.P.; Levine, A. The involvement of hydrogen peroxide in the differentiation of secondary walls in cotton fibers. Plant Physiol. 1999, 119, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, L.; Dong, F.; Gao, J.; Galbraith, W.; Song, C. Hydrogen Peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol. 2001, 126, 1428–1448. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Kao, C.H. Abscisic acid induced changes in cell wall peroxidase activity and hydrogen peroxide level in roots of rice seedlings. Plant Sci. 2001, 160, 323–329. [Google Scholar] [CrossRef]
- Ye, N.; Zhu, G.; Liu, Y.; Li, Y.; Zhang, J. ABA controls H2O2 accumulation through the induction of OsCATB in rice leaves under water stress. Plant Cell Physiol. 2011, 54, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Gao, P.; Li, L.; Yuan, Y.; Sun, J.; Guo, S. Abscicic acid-induced H2O2 accumulation enhances antioxidant capacity in pumpkin-grafted cucumber leaves under Ca(NO3)2 stress. Front. Plant Sci. 2016, 7, 1489. [Google Scholar] [CrossRef]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic acid is a major regulator of grape berry ripening onset. New insights into ABA signaling network. Front. Plant Sci. 2017, 8, 1093. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, P.; Lecourieux, D.; Kappel, C.; Cluzet, S.; Cramer, G.; Delrot, S.; Lecorieux, F. The basic Leucine-Zipper transcription factor ABSICISIC ACID RESPONSE ELEMENT-BINDING FACTOR2 is an important transcriptional regulator of absicisic-dependent grape berry ripening processes. Plant Physiol. 2014, 164, 365–383. [Google Scholar] [CrossRef] [Green Version]
- Sudawan, B.; Chang, C.; Chao, H.; Ku, M.S.; Yen, Y. Hydrogen cyanamide breaks grapevine buds dormancy in the summer through transient activation of gene expression and accumulatio of reactive oxygen and nitrogen species. BMC Plant Biol. 2016, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.; Zrig, A.; Geuns, J.M.C.; Khemira, H. Near-lethal heat treatment induced metabolic changes associated with endodormancy release of Superior Seedless grapevine cv. (Vitis vinifera L.) buds. Aust. J. Crop. Sci. 2014, 8, 486–494. [Google Scholar]
- Porcher, A.; Guérin, V.; Montrichard, F.; Lebrec, A.; Lothier, J.; Vian, A. Ascorbate glutathione-dependent H2O2 scavenging is an important process in axillary bud outgrowth in rosebush. Ann. Bot. 2020, 126, 1049–1062. [Google Scholar] [CrossRef]
- Azizi, P.; Rafi, M.Y.; Maziah, M.; Abdullah, S.N.A.; Hanafi, M.M.; Latif, M.A.; Rashid, A.A.; Sahebi, M. Understanding the shoot apical meristem regulation: Astudy of the phytohormones, auxin and cytokinin, in rice. Mech. Dev. 2015, 135, 1–15. [Google Scholar] [CrossRef]
- Noriega, X.; Pérez, F.J. Cell cycle genes are activated earlier than respiratory genes during release of grapevine buds from endodormancy. Plant Signal. Behav. 2017, 12, e1321189. [Google Scholar] [CrossRef]
- Pérez, F.J.; Noriega, X. Sprouting of paradormant and end dormant grapevine buds under conditions of forced growth: Similarities and differences. Planta 2018, 248, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Xia, X.J.; Guo, X.; Zhou, Y.H.; Shi, K.; Zhou, J.; Yu, J.Q. Apoplastic H2O2 plays a critical role in axillary bud outgrowth by altering auxin and cytokinin homeostasis in tomato plants. New Phytol. 2016, 211, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relationship between the degree of endodormancy and H2O2 levels in Thompson seedless grapevine buds grown in Santiago, Chile. The degree of dormancy was determined by means of a log-rank test between the probabilistic function KM of the buds collected before (reference) and after the onset of the ED [26]. H2O2 was determined by a chemiluminescence method [29].
Figure 1.
Relationship between the degree of endodormancy and H2O2 levels in Thompson seedless grapevine buds grown in Santiago, Chile. The degree of dormancy was determined by means of a log-rank test between the probabilistic function KM of the buds collected before (reference) and after the onset of the ED [26]. H2O2 was determined by a chemiluminescence method [29].

Figure 2.
Expression of peroxidase genes (VvPODs), peroxidase activity and H2O2 levels during grape bud endodormancy. (a) Relationship between H2O2 levels and peroxidase activity (VvPOD) during endodormancy in Thompson seedless grapevine buds grown in Santiago, Chile. (b) Changes in the expression of peroxidase genes (VvPOD) throughout bud growth in Tempranillo grapevines grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.
Figure 2.
Expression of peroxidase genes (VvPODs), peroxidase activity and H2O2 levels during grape bud endodormancy. (a) Relationship between H2O2 levels and peroxidase activity (VvPOD) during endodormancy in Thompson seedless grapevine buds grown in Santiago, Chile. (b) Changes in the expression of peroxidase genes (VvPOD) throughout bud growth in Tempranillo grapevines grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.
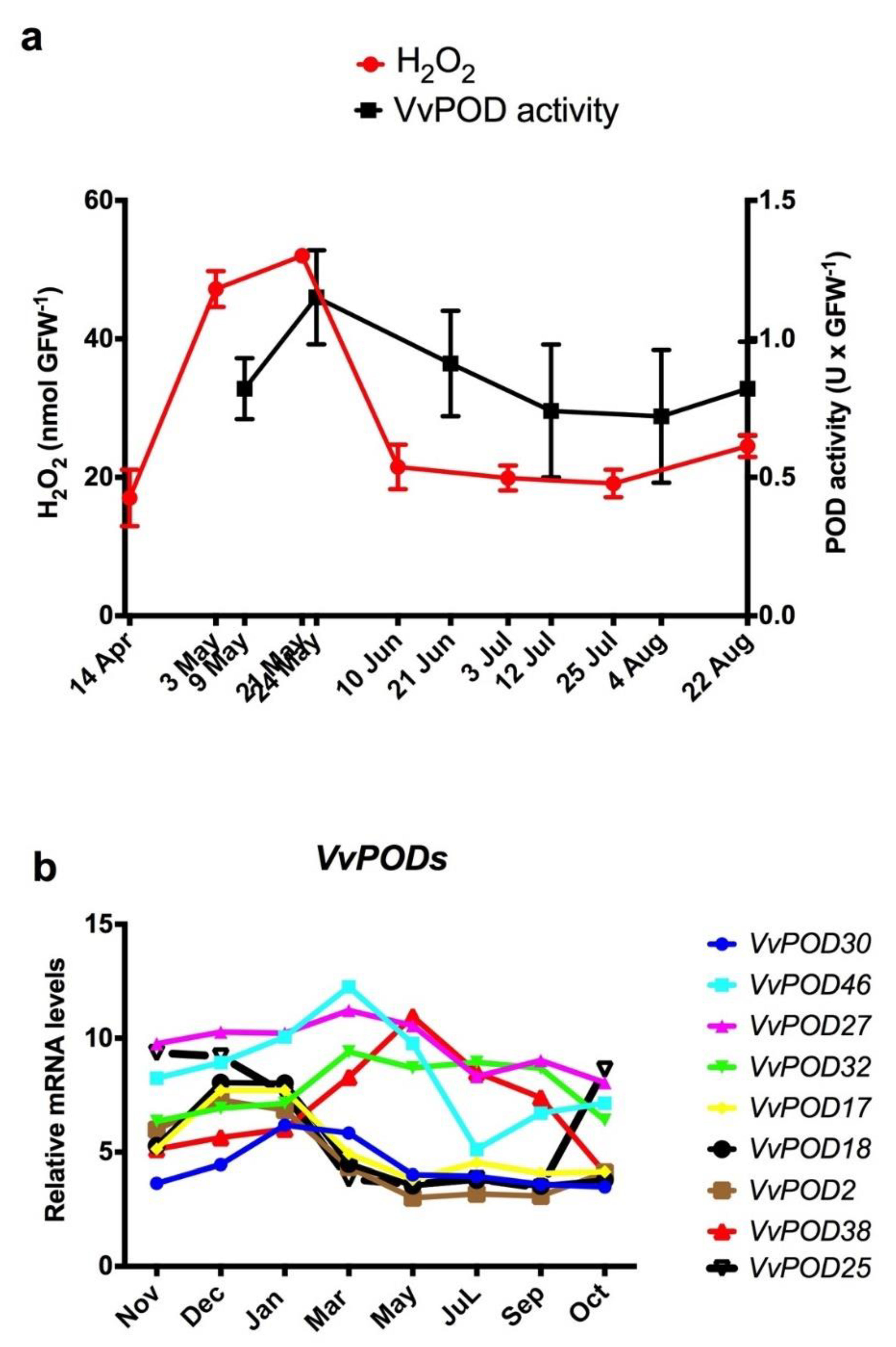
Figure 3.
Expression profile of RESPIRATORY BURST OXIDASE HOMOLOGS (VvRBOHs) genes in grapevine buds during endodormancy. The expression profile of VvRBOH genes in Tempranillo grapevine buds grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.
Figure 3.
Expression profile of RESPIRATORY BURST OXIDASE HOMOLOGS (VvRBOHs) genes in grapevine buds during endodormancy. The expression profile of VvRBOH genes in Tempranillo grapevine buds grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.
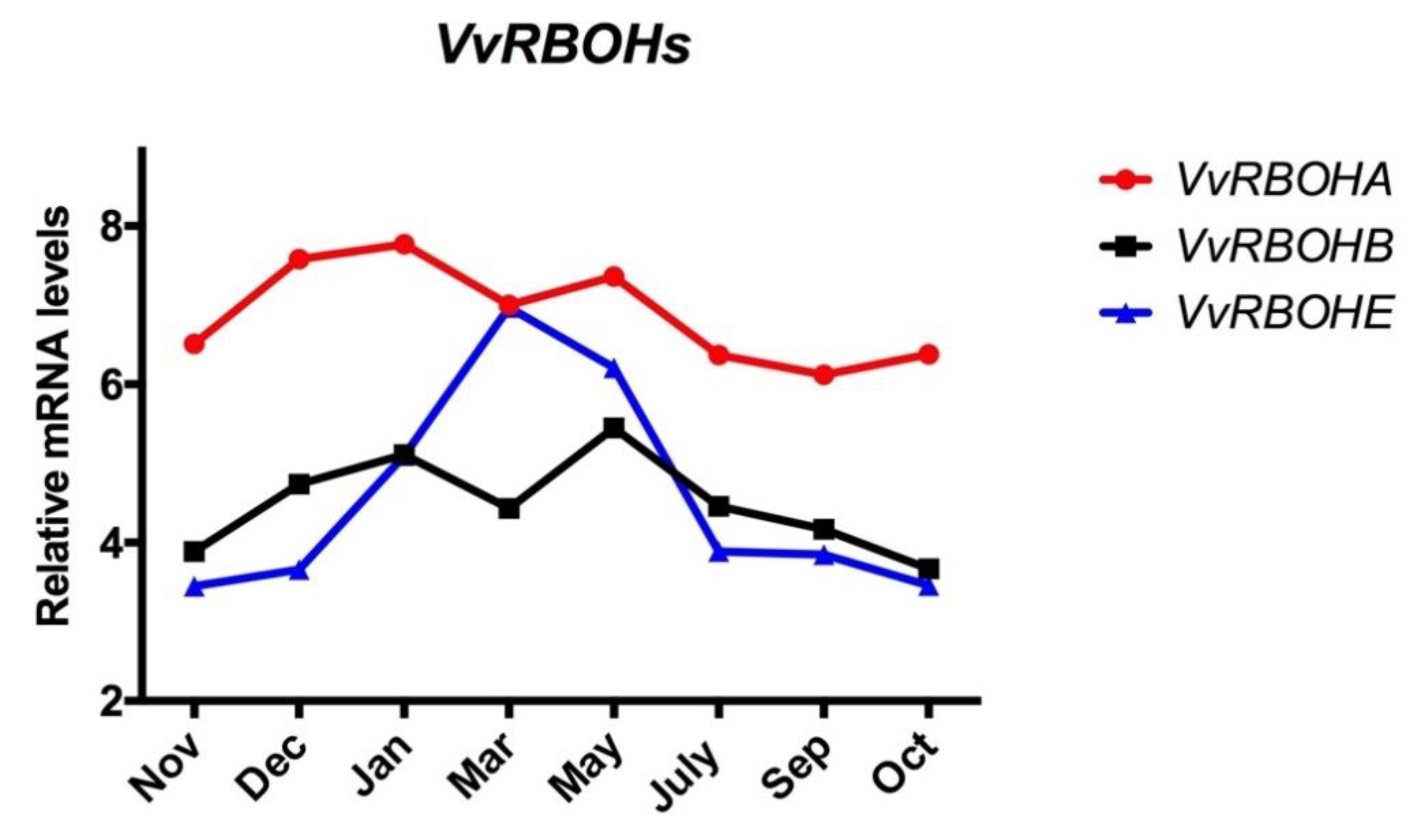
Figure 4.
Catalase activity and expression of catalase genes in grapevine buds during endodormancy. (a) Catalase activity during endodormancy in Thompson seedless grapevine buds grown in Santiago, Chile. (b) Expression of catalase genes (VvCAT1 and VvCAT2) in buds of Tempranillo grapevines grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.
Figure 4.
Catalase activity and expression of catalase genes in grapevine buds during endodormancy. (a) Catalase activity during endodormancy in Thompson seedless grapevine buds grown in Santiago, Chile. (b) Expression of catalase genes (VvCAT1 and VvCAT2) in buds of Tempranillo grapevines grown in Alcalá de Henares, Madrid [21] and adapted to the Southern Hemisphere conditions.

Figure 5.
Aminotriazole effect on H2O2 content and budbreak in grapevine buds. The effect of aminotriazole (a) on the level of H2O2 and (b) on the budbreak of Thompson seedless grapevine buds was analysed. Buds collected at June 11 were excised into single-bud cuttings and sprayed with 2.5% HC, 2% Aminotriazole and water as control. The treated single-bud cuttings were placed in the growth chamber and H2O2 was determined at the desired time by a chemiluminescence method [29]. Values are the average of three biological replicates ± SD; and (*) indicates statistically significance differences (Dunnett’s multiple comparison test α = 0.05). Buds collected at July 18 were excised into single-bud cuttings (30 per treatment) and sprayed with 2.5% HC, 2% Aminotriazole and water as control. The treated single-bud cuttings were placed in the growth chamber and budbreak was assessed by the presence of green tips.
Figure 5.
Aminotriazole effect on H2O2 content and budbreak in grapevine buds. The effect of aminotriazole (a) on the level of H2O2 and (b) on the budbreak of Thompson seedless grapevine buds was analysed. Buds collected at June 11 were excised into single-bud cuttings and sprayed with 2.5% HC, 2% Aminotriazole and water as control. The treated single-bud cuttings were placed in the growth chamber and H2O2 was determined at the desired time by a chemiluminescence method [29]. Values are the average of three biological replicates ± SD; and (*) indicates statistically significance differences (Dunnett’s multiple comparison test α = 0.05). Buds collected at July 18 were excised into single-bud cuttings (30 per treatment) and sprayed with 2.5% HC, 2% Aminotriazole and water as control. The treated single-bud cuttings were placed in the growth chamber and budbreak was assessed by the presence of green tips.
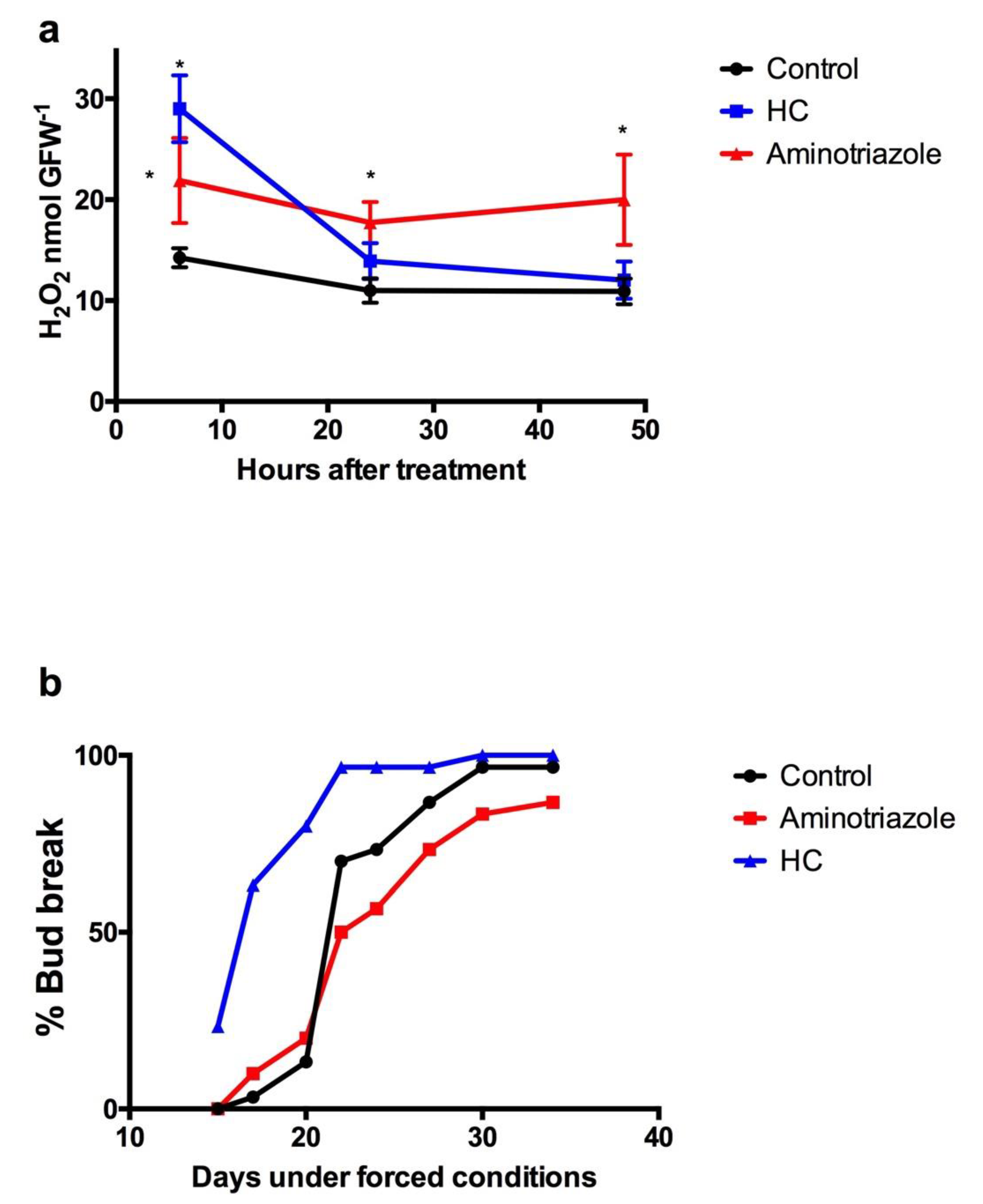
Figure 6.
Rate of H2O2 reduction in dormant and quiescent grapevine buds under forcing budbreak. Decreases in H2O2 content in (a) endodormant and (b) quiescent buds of Thompson seedless grapevines under forced growth conditions. H2O2 was determined by a chemiluminescence method [29]. Values are the average of three biological replicates ± SD; (*) indicates statistically significance differences. Student’s test (α = 0.05).
Figure 6.
Rate of H2O2 reduction in dormant and quiescent grapevine buds under forcing budbreak. Decreases in H2O2 content in (a) endodormant and (b) quiescent buds of Thompson seedless grapevines under forced growth conditions. H2O2 was determined by a chemiluminescence method [29]. Values are the average of three biological replicates ± SD; (*) indicates statistically significance differences. Student’s test (α = 0.05).
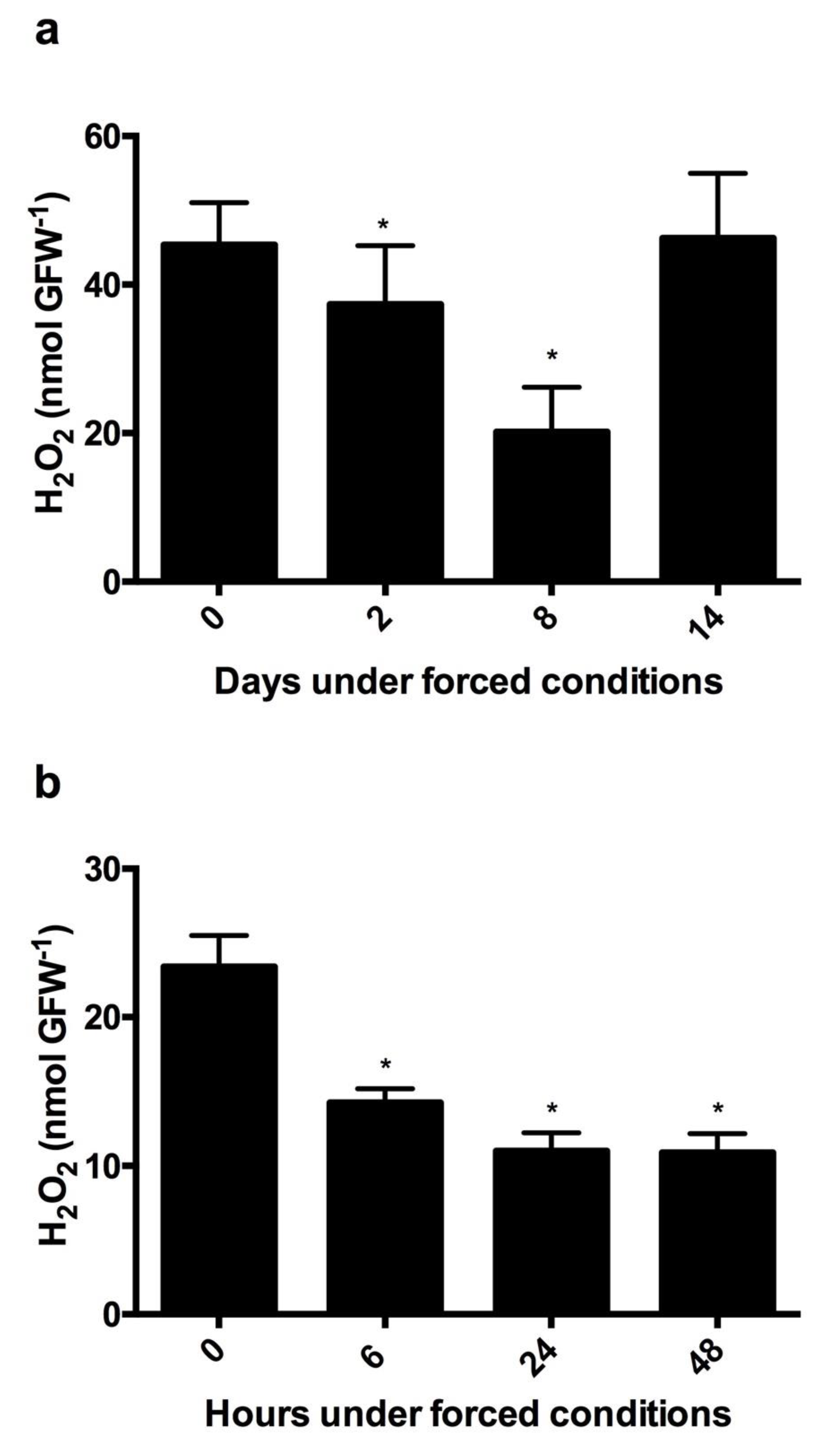
Figure 7.
Transcriptional effect of H2O2 on the homeostasis of CK and auxin related genes. Effect of H2O2 applications on the expression of (a) cytokinin (CK) VvIPT3a, VvLOG1, and VvCKX3 and (b) auxin VvAMI1, VvYUC3 and VvPIN3 genes. Gene expression analysis was performed by RT-qPCR and normalised against VvUBIQITIN. Values are the average of three biological replicates with three technical repetitions ± SD; (*) indicates statistically significance differences p ≤ 0.05; (**) p ≤ 0.01 (Dunnett’s multiple comparison test).
Figure 7.
Transcriptional effect of H2O2 on the homeostasis of CK and auxin related genes. Effect of H2O2 applications on the expression of (a) cytokinin (CK) VvIPT3a, VvLOG1, and VvCKX3 and (b) auxin VvAMI1, VvYUC3 and VvPIN3 genes. Gene expression analysis was performed by RT-qPCR and normalised against VvUBIQITIN. Values are the average of three biological replicates with three technical repetitions ± SD; (*) indicates statistically significance differences p ≤ 0.05; (**) p ≤ 0.01 (Dunnett’s multiple comparison test).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pérez, F.J.; Noriega, X.; Rubio, S. Hydrogen Peroxide Increases during Endodormancy and Decreases during Budbreak in Grapevine (Vitis vinifera L.) Buds. Antioxidants 2021, 10, 873. https://doi.org/10.3390/antiox10060873
AMA Style
Pérez FJ, Noriega X, Rubio S. Hydrogen Peroxide Increases during Endodormancy and Decreases during Budbreak in Grapevine (Vitis vinifera L.) Buds. Antioxidants. 2021; 10(6):873. https://doi.org/10.3390/antiox10060873
Chicago/Turabian StylePérez, Francisco Javier, Ximena Noriega, and Sebastián Rubio. 2021. "Hydrogen Peroxide Increases during Endodormancy and Decreases during Budbreak in Grapevine (Vitis vinifera L.) Buds" Antioxidants 10, no. 6: 873. https://doi.org/10.3390/antiox10060873
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.