
Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer

 , and
, and 
Abstract
1. Introduction
1.1. Chemoprevention of Cancer
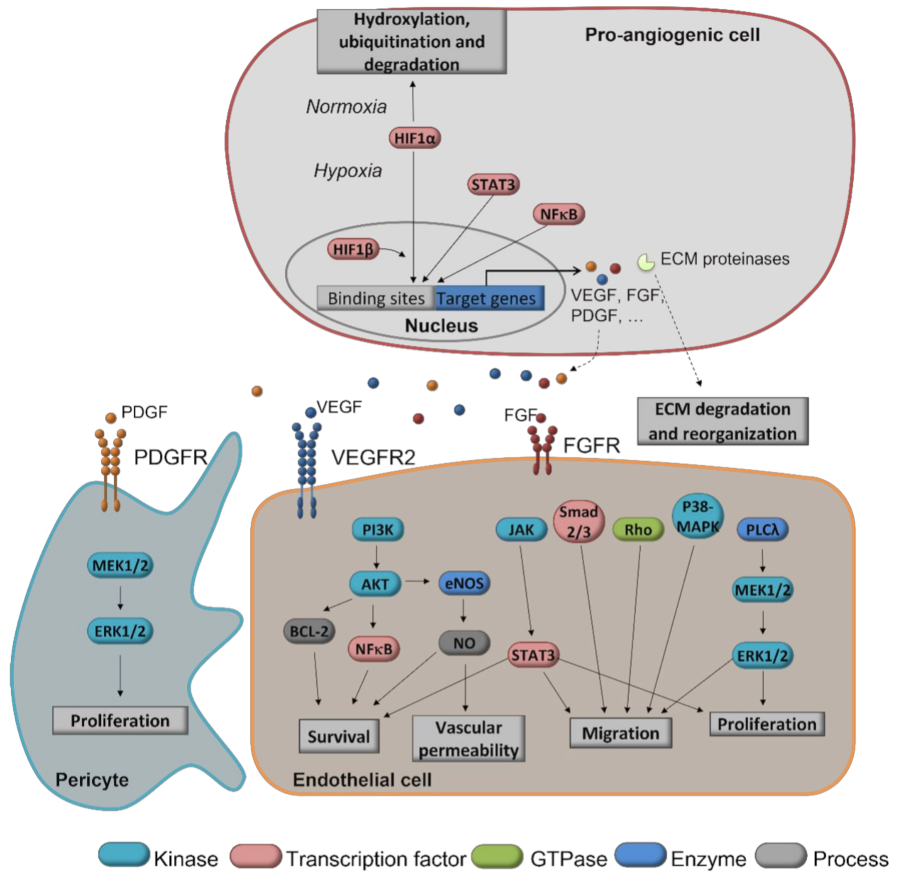
1.2. Angiogenesis as a Target for Chemoprevention of Cancer
2. Plant-Derived Compounds Constituent of Diet with Antiangiogenic Activity
2.1. Phenolic and Polyphenolic Phytochemicals
2.1.1. Flavonols
Quercetin
Myricetin
Other Flavonols
2.1.2. Isoflavones
2.1.3. Green Tea Polyphenols
2.1.4. Other Phenolic and Polyphenolic Compounds
2.2. Terpenes
2.2.1. Ursolic Acid
2.2.2. Kahweol
2.2.3. Other Terpenoids
2.3. Anthraquinones
2.3.1. Emodin
2.3.2. Aloe-Emodin
3. Concluding Remarks and Future Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 28 December 2021).
- Barry, M.J.; Simmons, L.H. Prevention of Prostate Cancer Morbidity and Mortality: Primary Prevention and Early Detection. Med. Clin. N. Am. 2017, 101, 787–806. [Google Scholar] [CrossRef] [PubMed]
- Cooperman, A.M.; Iskandar, M.E.; Wayne, M.G.; Steele, J.G. Prevention and Early Detection of Pancreatic Cancer. Surg. Clin. N. Am. 2018, 98, 1–12. [Google Scholar] [CrossRef] [PubMed]
- New, M.; Keith, R. Early Detection and Chemoprevention of Lung Cancer. F1000Research 2018, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Loomans-Kropp, H.A.; Umar, A. Cancer Prevention and Screening: The next Step in the Era of Precision Medicine. NPJ Precis. Oncol. 2019, 3, 3. [Google Scholar] [CrossRef]
- Sporn, M.B. Approaches to Prevention of Epithelial Cancer during the Preneoplastic Period. Cancer Res. 1976, 36, 2699–2702. [Google Scholar]
- Meyskens, F.L.; Mukhtar, H.; Rock, C.L.; Cuzick, J.; Kensler, T.W.; Yang, C.S.; Ramsey, S.D.; Lippman, S.M.; Alberts, D.S. Cancer Prevention: Obstacles, Challenges, and the Road Ahead. J. Natl. Cancer Inst. 2016, 108, djv309. [Google Scholar] [CrossRef]
- Blackburn, E.H. Highlighting the Science of Cancer Prevention. Cancer Prev. Res. 2010, 3, 393. [Google Scholar] [CrossRef][Green Version]
- Albini, A.; Tosetti, F.; Li, V.W.; Noonan, D.M.; Li, W.W. Cancer Prevention by Targeting Angiogenesis. Nat. Rev. Clin. Oncol. 2012, 9, 498–509. [Google Scholar] [CrossRef]
- Martínez-Poveda, B.; Torres-Vargas, J.A.; del Carmen Ocaña, M.; García-Caballero, M.; Medina, M.Á.; Quesada, A.R. The Mediterranean Diet, a Rich Source of Angiopreventive Compounds in Cancer. Nutrients 2019, 11, 2036. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor Angiogenesis: Causes, Consequences, Challenges and Opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Angiogenesis. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Elsevier Inc.: Amsterdan, The Netherlands, 2013; pp. 130–132. ISBN 978-0-0809-6156-9. [Google Scholar]
- Gerritsen, M.E. Angiogenesis. In Microcirculation, 2nd ed.; Tuma, R.F., Durán, W.N., Ley, K., Eds.; Elsevier Inc.: Amsterdan, The Netherlands, 2008; pp. 351–383. ISBN 978-0-1237-4530-9. [Google Scholar]
- Wang, Z.; Dabrosin, C.; Yin, X.; Fuster, M.M.; Arreola, A.; Rathmell, W.K.; Generali, D.; Nagaraju, G.P.; El-Rayes, B.; Ribatti, D.; et al. Broad Targeting of Angiogenesis for Cancer Prevention and Therapy. Semin. Cancer Biol. 2015, 35, S224–S243. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus Guidelines for the Use and Interpretation of Angiogenesis Assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef]
- Eccles, S.A.; Court, W.; Patterson, L. In Vitro Assays for Endothelial Cell Functions Required for Angiogenesis: Proliferation, Motility, Tubular Differentiation, and Matrix Proteolysis. In Angiogenesis Protocols; Martin, S., Hewett, P., Eds.; Humana Press: New York, NY, USA, 2016; Volume 1430, pp. 121–147. [Google Scholar]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Quesada, A.R.; Barbacid, M.M.; Mira, E.; Fernández-Resa, P.; Márquez, G.; Aracil, M. Evaluation of Fluorometric and Zymographic Methods as Activity Assays for Stromelysins and Gelatinases. Clin. Exp. Metastasis 1997, 15, 26–32. [Google Scholar] [CrossRef]
- Jin, J.; Hu, X.; Zhang, L.; Wang, J. Aorta Ring Assay. Bio-Protocol 2016, 6, e1856. [Google Scholar] [CrossRef]
- Naik, M.; Brahma, P.; Dixit, M. A Cost-Effective and Efficient Chick Ex-Ovo Cam Assay Protocol to Assess Angiogenesis. Methods Protoc. 2018, 1, 19. [Google Scholar] [CrossRef]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane (CAM). A Multifaceted Experimental Model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Currie, P.D. Animal Models of Human Disease: Zebrafish Swim into View. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Serbedzija, G.N.; Flynn, E.; Willett, C.E. Zebrafish Angiogenesis: A New Model for Drug Screening. Angiogenesis 1999, 3, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I.; Peterson, R.T. In Vivo Drug Discovery in the Zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef] [PubMed]
- García-Caballero, M.; Quesada, A.R.; Medina, M.Á.; Marí-Beffa, M. Fishing Anti(Lymph)Angiogenic Drugs with Zebrafish. Drug Discov. Today 2018, 23, 366–374. [Google Scholar] [CrossRef]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted Cancer Prevention by Quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef]
- Chien, S.-Y.; Wu, Y.-C.; Chung, J.-G.; Yang, J.-S.; Lu, H.-F.; Tsou, M.-F.; Wood, W.; Kuo, S.-J.; Chen, D.-R. Quercetin-Induced Apoptosis Acts through Mitochondrial- and Caspase-3-Dependent Pathways in Human Breast Cancer MDA-MB-231 Cells. Hum. Exp. Toxicol. 2009, 28, 493–503. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin Induces Protective Autophagy in Gastric Cancer Cells: Involvement of Akt-MTOR- and Hypoxia-Induced Factor 1α-Mediated Signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef]
- Mouria, M.; Gukovskaya, A.S.; Jung, Y.; Buechler, P.; Hines, O.J.; Reber, H.A.; Pandol, S.J. Food-Derived Polyphenols Inhibit Pancreatic Cancer Growth through Mitochondrial Cytochrome C Release and Apoptosis. Int. J. Cancer 2002, 98, 761–769. [Google Scholar] [CrossRef] [PubMed]
- McGill, C.M.; Brown, T.J.; Fisher, L.N.; Gustafson, S.J.; Dunlap, K.L.; Beck, A.J.; Toran, P.T.; Claxton, D.F.; Barth, B.M. Combinatorial Efficacy of Quercitin and Nanoliposomal Ceramide for Acute Myeloid Leukemia. Int. J. Biopharm. Sci. 2018, 1, 106. [Google Scholar] [CrossRef] [PubMed]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and Cancer: New Insights into Its Therapeutic Effects on Ovarian Cancer Cells. Cell Biosci. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin Inhibits Prostate Cancer by Attenuating Cell Survival and Inhibiting Anti-Apoptotic Pathways. World J. Surg. Oncol. 2018, 16, 108. [Google Scholar] [CrossRef]
- Najafi, M.; Tavakol, S.; Zarrabi, A.; Ashrafizadeh, M. Dual Role of Quercetin in Enhancing the Efficacy of Cisplatin in Chemotherapy and Protection against Its Side Effects: A Review. Arch. Physiol. Biochem. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Tan, W.; Lin, L.; Li, M.; Zhang, Y.-X.; Tong, Y.; Xiao, D.; Ding, J. Quercetin, a Dietary-Derived Flavonoid, Possesses Antiangiogenic Potential. Eur. J. Pharmacol. 2003, 459, 255–262. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Molecular Mechanisms and Clinical Applications of Angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef]
- Chung, A.S.; Lee, J.; Ferrara, N. Targeting the Tumour Vasculature: Insights from Physiological Angiogenesis. Nat. Rev. Cancer 2010, 10, 505–514. [Google Scholar] [CrossRef]
- Cross, M.J.; Claesson-Welsh, L. FGF and VEGF Function in Angiogenesis: Signalling Pathways, Biological Responses and Therapeutic Inhibition. Trends Pharmacol. Sci. 2001, 22, 201–207. [Google Scholar] [CrossRef]
- Pezzuto, A.; Carico, E. Role of HIF-1 in Cancer Progression: Novel Insights. A Review. Curr. Mol. Med. 2018, 18, 343–351. [Google Scholar] [CrossRef]
- García-Mediavilla, V.; Crespo, I.; Collado, P.S.; Esteller, A.; Sánchez-Campos, S.; Tuñón, M.J.; González-Gallego, J. The Anti-Inflammatory Flavones Quercetin and Kaempferol Cause Inhibition of Inducible Nitric Oxide Synthase, Cyclooxygenase-2 and Reactive C-Protein, and down-Regulation of the Nuclear Factor KappaB Pathway in Chang Liver Cells. Eur. J. Pharmacol. 2007, 557, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.J.T.; Venema, R.C. Quercetin Inhibits ENOS, Microtubule Polymerization, and Mitotic Progression in Bovine Aortic Endothelial Cells1. J. Nutr. 2006, 136, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Shi, D.; Liu, L.; Wang, J.; Xie, X.; Kang, T.; Deng, W. Quercetin Suppresses Cyclooxygenase-2 Expression and Angiogenesis through Inactivation of P300 Signaling. PLoS ONE 2011, 6, e22934. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Budhraja, A.; Son, Y.-O.; Wang, X.; Zhang, Z.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.-C.; Xu, M.; et al. Quercetin Inhibits Angiogenesis Mediated Human Prostate Tumor Growth by Targeting VEGFR- 2 Regulated AKT/MTOR/P70S6K Signaling Pathways. PLoS ONE 2012, 7, e47516. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Far, A.D.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and Apoptosis-Inducing Effects of Quercetin in Vitro and in Vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef]
- Angst, E.; Park, J.L.; Moro, A.; Lu, Q.Y.; Lu, X.; Li, G.; King, J.; Chen, M.; Reber, H.A.; Go, V.L.W.; et al. The Flavonoid Quercetin Inhibits Pancreatic Cancer Growth in Vitro and in Vivo. Pancreas 2013, 42, 223–229. [Google Scholar] [CrossRef]
- Fernández-Palanca, P.; Fondevila, F.; Méndez-Blanco, C.; Tuñón, M.J.; González-Gallego, J.; Mauriz, J.L. Antitumor Effects of Quercetin in Hepatocarcinoma In Vitro and In Vivo Models: A Systematic Review. Nutrients 2019, 11, 2875. [Google Scholar] [CrossRef]
- Lu, X.; Yang, F.; Chen, D.; Zhao, Q.; Chen, D.; Ping, H.; Xing, N. Quercetin Reverses Docetaxel Resistance in Prostate Cancer via Androgen Receptor and PI3K/Akt Signaling Pathways. Int. J. Biol. Sci. 2020, 16, 1121–1134. [Google Scholar] [CrossRef]
- Baksi, R.; Singh, D.P.; Borse, S.P.; Rana, R.; Sharma, V.; Nivsarkar, M. In Vitro and in Vivo Anticancer Efficacy Potential of Quercetin Loaded Polymeric Nanoparticles. Biomed. Pharmacother. 2018, 106, 1513–1526. [Google Scholar] [CrossRef]
- Ravichandiran, V.; Masilamani, K.; Senthilnathan, B.; Maheshwaran, A.; Wong, T.W.; Roy, P. Quercetin-Decorated Curcumin Liposome Design for Cancer Therapy: In-Vitro and In-Vivo Studies. Curr. Drug Deliv. 2017, 14, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Children’s Hospital Medical Center, Cincinnati. Quercetin Chemoprevention for Squamous Cell Carcinoma in Patients with Fanconi Anemia. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03476330?term=quercitin&cond=cancer&draw=3&rank=11 (accessed on 28 December 2021).
- Ribeiro de Lima, M.T.; Waffo-Téguo, P.; Teissedre, P.L.; Pujolas, A.; Vercauteren, J.; Cabanis, J.C.; Mérillon, J.M. Determination of Stilbenes (Trans-Astringin, Cis- and Trans-Piceid, and Cis- and Trans-Resveratrol) in Portuguese Wines. J. Agric. Food Chem. 1999, 47, 2666–2670. [Google Scholar] [CrossRef] [PubMed]
- Sellappan, S.; Akoh, C.C. Flavonoids and Antioxidant Capacity of Georgia-Grown Vidalia Onions. J. Agric. Food Chem. 2002, 50, 5338–5342. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kang, N.J.; Rogozin, E.A.; Kim, H.-G.; Cho, Y.Y.; Bode, A.M.; Lee, H.J.; Surh, Y.-J.; Bowden, G.T.; Dong, Z. Myricetin Is a Novel Natural Inhibitor of Neoplastic Cell Transformation and MEK1. Carcinogenesis 2007, 28, 1918–1927. [Google Scholar] [CrossRef]
- Semwal, D.; Semwal, R.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-Tumor Effects and Associated Molecular Mechanisms of Myricetin. Biomed. Pharmacother. 2019, 120, 109506. [Google Scholar] [CrossRef]
- Feng, J.; Chen, X.; Wang, Y.; Du, Y.; Sun, Q.; Zang, W.; Zhao, G. Myricetin Inhibits Proliferation and Induces Apoptosis and Cell Cycle Arrest in Gastric Cancer Cells. Mol. Cell. Biochem. 2015, 408, 163–170. [Google Scholar] [CrossRef]
- Zang, W.; Wang, T.; Wang, Y.; Li, M.; Xuan, X.; Ma, Y.; Du, Y.; Liu, K.; Dong, Z.; Zhao, G. Myricetin Exerts Anti-Proliferative, Anti-Invasive, and pro-Apoptotic Effects on Esophageal Carcinoma EC9706 and KYSE30 Cells via RSK2. Tumor Biol. 2014, 35, 12583–12592. [Google Scholar] [CrossRef]
- Huang, H.; Chen, A.Y.; Rojanasakul, Y.; Ye, X.; Rankin, G.O.; Chen, Y.C. Dietary Compounds Galangin and Myricetin Suppress Ovarian Cancer Cell Angiogenesis. J. Funct. Foods 2015, 15, 464–475. [Google Scholar] [CrossRef]
- Jung, S.K.; Lee, K.W.; Byun, S.; Lee, E.J.; Kim, J.E.; Bode, A.M.; Dong, Z.; Lee, H.J. Myricetin inhibits UVB-induced angiogenesis by regulating PI-3 kinase in vivo. Carcinogenesis 2010, 31, 911–917. [Google Scholar] [CrossRef]
- Kim, J.D.; Liu, L.; Guo, W.; Meydani, M. Chemical Structure of Flavonols in Relation to Modulation of Angiogenesis and Immune-Endothelial Cell Adhesion. J. Nutr. Biochem. 2006, 17, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D. Myricetin Inhibits Angiogenesis by Inducing Apoptosis and Suppressing PI3K/Akt/MTOR Signaling in Endothelial Cells. J. Cancer Prev. 2017, 22, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.K.; Jung, I.; Kim, M.E.; Bae, S.K.; Lee, J.S. Anti-Cancer Activity of Myricetin against Human Papillary Thyroid Cancer Cells Involves Mitochondrial Dysfunction–Mediated Apoptosis. Biomed. Pharmacother. 2017, 91, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Hu, Q.; Wang, J.; Liu, Z.; Wu, D.; Lu, W.; Huang, J. Myricetin Is a Novel Inhibitor of Human Inosine 5′-Monophosphate Dehydrogenase with Anti-Leukemia Activity. Biochem. Biophys. Res. Commun. 2016, 477, 915–922. [Google Scholar] [CrossRef]
- Zheng, A.W.; Chen, Y.Q.; Zhao, L.Q.; Feng, J.G. Myricetin Induces Apoptosis and Enhances Chemosensitivity in Ovarian Cancer Cells. Oncol. Lett. 2017, 13, 4974–4978. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, Q.; Wu, S.; Yi, D.; Yu, Y.; Liu, S.; Li, S.; Li, Z. Myricetin Induces Apoptosis via Endoplasmic Reticulum Stress and DNA Double-Strand Breaks in Human Ovarian Cancer Cells. Mol. Med. Rep. 2016, 13, 2094–2100. [Google Scholar] [CrossRef]
- Siegelin, M.D.; Gaiser, T.; Habel, A.; Siegelin, Y. Myricetin Sensitizes Malignant Glioma Cells to TRAIL-Mediated Apoptosis by down-Regulation of the Short Isoform of FLIP and Bcl-2. Cancer Lett. 2009, 283, 230–238. [Google Scholar] [CrossRef]
- Jiao, D.; Zhang, X.D. Myricetin Suppresses P21-Activated Kinase 1 in Human Breast Cancer MCF-7 Cells through Downstream Signaling of the β-Catenin Pathway. Oncol. Rep. 2016, 36, 342–348. [Google Scholar] [CrossRef]
- Knickle, A.; Fernando, W.; Greenshields, A.L.; Rupasinghe, H.P.V.; Hoskin, D.W. Myricetin-Induced Apoptosis of Triple-Negative Breast Cancer Cells Is Mediated by the Iron-Dependent Generation of Reactive Oxygen Species from Hydrogen Peroxide. Food Chem. Toxicol. 2018, 118, 154–167. [Google Scholar] [CrossRef]
- Jo, S.; Ha, T.K.; Han, S.H.; Kim, M.E.; Jung, I.; Lee, H.W.; Bae, S.K.; Lee, J.S. Myricetin Induces Apoptosis of Human Anaplastic Thyroid Cancer Cells via Mitochondria Dysfunction. Anticancer Res. 2017, 37, 1705–1710. [Google Scholar] [CrossRef]
- Park, H.; Park, S.; Bazer, F.W.; Lim, W.; Song, G. Myricetin Treatment Induces Apoptosis in Canine Osteosarcoma Cells by Inducing DNA Fragmentation, Disrupting Redox Homeostasis, and Mediating Loss of Mitochondrial Membrane Potential. J. Cell. Physiol. 2018, 233, 7457–7466. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lim, W.; Bazer, F.W.; Song, G. Myricetin Suppresses Invasion and Promotes Cell Death in Human Placental Choriocarcinoma Cells through Induction of Oxidative Stress. Cancer Lett. 2017, 399, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, S.-X.; Sun, S.-Y.; Shi, W.-N.; Song, Z.-Y.; Wang, S.-Q.; Yu, X.-F.; Gao, Z.-H.; Qu, X.-J. Chemoprevention of Intestinal Tumorigenesis by the Natural Dietary Flavonoid Myricetin in APCMin/+ Mice. Oncotarget 2016, 7, 60446–60460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Su, H.; Yan, J.Y.; Li, N.; Song, Z.Y.; Wang, H.J.; Huo, L.G.; Wang, F.; Ji, W.S.; Qu, X.J.; et al. Chemopreventive Effect of Myricetin, a Natural Occurring Compound, on Colonic Chronic Inflammation and Inflammation-Driven Tumorigenesis in Mice. Biomed. Pharmacother. 2018, 97, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, C.; Huang, H.; Yang, B.; Xiao, G.; Kong, D.; Tian, Q.; Song, Q.; Song, Y.; Tan, H.; et al. The Natural Compound Myricetin Effectively Represses the Malignant Progression of Prostate Cancer by Inhibiting PIM1 and Disrupting the PIM1/CXCR4 Interaction. Cell. Physiol. Biochem. 2018, 48, 1230–1244. [Google Scholar] [CrossRef]
- Phillips, P.A.; Sangwan, V.; Borja-Cacho, D.; Dudeja, V.; Vickers, S.M.; Saluja, A.K. Myricetin Induces Pancreatic Cancer Cell Death via the Induction of Apoptosis and Inhibition of the Phosphatidylinositol 3-Kinase (PI3K) Signaling Pathway. Cancer Lett. 2011, 308, 181–188. [Google Scholar] [CrossRef]
- Varela-Rodríguez, L.; Sánchez-Ramírez, B.; Hernández-Ramírez, V.I.; Varela-Rodríguez, H.; Castellanos-Mijangos, R.D.; González-Horta, C.; Chávez-Munguía, B.; Talamás-Rohana, P. Effect of Gallic Acid and Myricetin on Ovarian Cancer Models: A Possible Alternative Antitumoral Treatment. BMC Complement. Med. Ther. 2020, 20, 110. [Google Scholar] [CrossRef]
- Song, X.; Rao, H.; Guo, C.; Yang, B.; Ren, Y.; Wang, M.; Li, Y.; Cao, Z.; Pei, J. Myricetin Exhibit Selective Anti-Lymphoma Activity by Targeting BTK and Is Effective via Oral Administration in Vivo. Phytomedicine 2021, 93, 153802. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.J.; Tang, X.J.; Du, L.; Li, F. In Vitro and in Vivo Evaluation of Functionalized Chitosan-Pluronic Micelles Loaded with Myricetin on Glioblastoma Cancer. Nanomedicine 2016, 12, 1263–1278. [Google Scholar] [CrossRef]
- Li, M.; Zha, G.; Chen, R.; Chen, X.; Sun, Q.; Jiang, H. Anticancer Effects of Myricetin Derivatives in Non-Small Cell Lung Cancer in Vitro and in Vivo. Pharmacol. Res. Perspect. 2022, 10, e00905. [Google Scholar] [CrossRef]
- Wang, L.; Feng, J.; Chen, X.; Guo, W.; Du, Y.; Wang, Y.; Zang, W.; Zhang, S.; Zhao, G. Myricetin Enhance Chemosensitivity of 5-Fluorouracil on Esophageal Carcinoma in Vitro and in Vivo. Cancer Cell Int. 2014, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.-R.; Kunimasa, K.; Kumazawa, S.; Nakayama, T.; Kaji, K.; Uto, Y.; Hori, H.; Nagasawa, H.; Ohta, T. Correlation between Antiangiogenic Activity and Antioxidant Activity of Various Components from Propolis. Mol. Nutr. Food Res. 2009, 53, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and Inflammation: From Chemistry to Medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, J.H. Kaempferol Inhibits Pancreatic Cancer Cell Growth and Migration through the Blockade of EGFR-Related Pathway in Vitro. PLoS ONE 2016, 11, e0155264. [Google Scholar] [CrossRef] [PubMed]
- Kashafi, E.; Moradzadeh, M.; Mohamadkhani, A.; Erfanian, S. Kaempferol Increases Apoptosis in Human Cervical Cancer HeLa Cells via PI3K/AKT and Telomerase Pathways. Biomed. Pharmacother. 2017, 89, 573–577. [Google Scholar] [CrossRef]
- Kapoor, S. Attenuating Effect of Kaempferol on Neoplastic Growth in Different Urological Malignancies. Biomed. Pharmacother. 2018, 104, 848. [Google Scholar] [CrossRef]
- Han, X.; Liu, C.F.; Gao, N.; Zhao, J.; Xu, J. Kaempferol Suppresses Proliferation but Increases Apoptosis and Autophagy by Up-Regulating MicroRNA-340 in Human Lung Cancer Cells. Biomed. Pharmacother. 2018, 108, 809–816. [Google Scholar] [CrossRef]
- Li, S.; Yan, T.; Deng, R.; Jiang, X.; Xiong, H.; Wang, Y.; Yu, Q.; Wang, X.; Chen, C.; Zhu, Y. Low Dose of Kaempferol Suppresses the Migration and Invasion of Triple-Negative Breast Cancer Cells by Downregulating the Activities of RhoA and Rac1. OncoTargets Ther. 2017, 10, 4809–4819. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with Kaempferol Suppresses Breast Cancer Cell Growth Caused by Estrogen and Triclosan in Cellular and Xenograft Breast Cancer Models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef]
- Lee, G.-A.; Choi, K.-C.; Hwang, K.-A. Kaempferol, a Phytoestrogen, Suppressed Triclosan-Induced Epithelial-Mesenchymal Transition and Metastatic-Related Behaviors of MCF-7 Breast Cancer Cells. Environ. Toxicol. Pharmacol. 2017, 49, 48–57. [Google Scholar] [CrossRef]
- Lee, G.A.; Choi, K.C.; Hwang, K.A. Treatment with Phytoestrogens Reversed Triclosan and Bisphenol A-Induced Anti-Apoptosis in Breast Cancer Cells. Biomol. Ther. 2018, 26, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xue, L. Kaempferol Suppresses Proliferation and Induces Cell Cycle Arrest, Apoptosis, and DNA Damage in Breast Cancer Cells. Oncol. Res. 2019, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.W.; Chen, P.N.; Wu, H.C.; Wu, S.W.; Tsai, P.Y.; Hsieh, Y.S.; Chang, H.R. Kaempferol Inhibits the Invasion and Migration of Renal Cancer Cells through the Downregulation of AKT and FAK Pathways. Int. J. Med. Sci. 2017, 14, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Dang, Q.; Song, W.; Xu, D.; Ma, Y.; Li, F.; Zeng, J.; Zhu, G.; Wang, X.; Chang, L.S.; He, D.; et al. Kaempferol Suppresses Bladder Cancer Tumor Growth by Inhibiting Cell Proliferation and Inducing Apoptosis. Mol. Carcinog. 2015, 54, 831–840. [Google Scholar] [CrossRef]
- Song, H.; Bao, J.; Wei, Y.; Chen, Y.; Mao, X.; Li, J.; Yang, Z.; Xue, Y. Kaempferol Inhibits Gastric Cancer Tumor Growth: An in Vitro and in Vivo Study. Oncol. Rep. 2015, 33, 868–874. [Google Scholar] [CrossRef]
- Luo, H.; Rankin, G.O.; Liu, L.; Daddysman, M.K.; Jiang, B.-H.; Chen, Y.C. Kaempferol Inhibits Angiogenesis and VEGF Expression through Both HIF Dependent and Independent Pathways in Human Ovarian Cancer Cells. Nutr. Cancer 2009, 61, 554–563. [Google Scholar] [CrossRef]
- Chin, H.K.; Horng, C.T.; Liu, Y.S.; Lu, C.C.; Su, C.Y.; Chen, P.S.; Chiu, H.Y.; Tsai, F.J.; Shieh, P.C.; Yang, J.S. Kaempferol Inhibits Angiogenic Ability by Targeting VEGF Receptor-2 and Downregulating the PI3K/AKT, MEK and ERK Pathways in VEGF-Stimulated Human Umbilical Vein Endothelial Cells. Oncol. Rep. 2018, 39, 2351–2357. [Google Scholar] [CrossRef]
- Lee, C.-F.; Yang, J.; Tsai, F.-J.; Chiang, N.-N.; Lu, C.; Huang, Y.-S.; Chen, C.; Chen, F.-A. Kaempferol Induces ATM/P53-Mediated Death Receptor and Mitochondrial Apoptosis in Human Umbilical Vein Endothelial Cells. Int. J. Oncol. 2016, 48, 2007–2014. [Google Scholar] [CrossRef]
- Sundarraj, K.; Raghunath, A.; Perumal, E. A Review on the Chemotherapeutic Potential of Fisetin: In Vitro Evidences. Biomed. Pharmacother. 2018, 97, 928–940. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Sak, K.; Tuli, H.S.; Buttar, H.S.; Bishayee, A. Fisetin: A Bioactive Phytochemical with Potential for Cancer Prevention and Pharmacotherapy. Life Sci. 2018, 194, 75–87. [Google Scholar] [CrossRef]
- Jia, S.; Xu, X.; Zhou, S.; Chen, Y.; Ding, G.; Cao, L. Fisetin Induces Autophagy in Pancreatic Cancer Cells via Endoplasmic Reticulum Stress- and Mitochondrial Stress-Dependent Pathways. Cell Death Dis. 2019, 10, 142. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, Y.; Gao, Z.; Li, X.; Weng, M.; Shi, C.; Wang, C.; Sun, L. Fisetin Inhibits the Proliferation, Migration and Invasion of Pancreatic Cancer by Targeting PI3K/AKT/MTOR Signaling. Aging 2021, 13, 24753–24767. [Google Scholar] [CrossRef] [PubMed]
- Bhat, T.A.; Nambiar, D.; Pal, A.; Agarwal, R.; Singh, R.P. Fisetin Inhibits Various Attributes of Angiogenesis in Vitro and in Vivo-Implications for Angioprevention. Carcinogenesis 2012, 33, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jang, Y.J.; Choi, Y.J.; Jang, J.W.; Kim, J.H.; Rho, Y.K.; Kim, I.J.; Kim, H.J.; Leem, M.J.; Lee, S.T. Fisetin Inhibits Matrix Metalloproteinases and Reduces Tumor Cell Invasiveness and Endothelial Cell Tube Formation. Nutr. Cancer 2013, 65, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.S.; Shen, K.H.; Huang, J.S.; Ko, S.C.; Shih, Y.W. Antimetastatic Potential of Fisetin Involves Inactivation of the PI3K/Akt and JNK Signaling Pathways with Downregulation of MMP-2/9 Expressions in Prostate Cancer PC-3 Cells. Mol. Cell. Biochem. 2010, 333, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.; Samadder, T.; Gupta, S.; Surolia, A.; Shaha, C. Anticancer Activity of a Combination of Cisplatin and Fisetin in Embryonal Carcinoma Cells and Xenograft Tumors. Mol. Cancer Ther. 2011, 10, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Rogero, M.M.; Fisberg, M.; Waitzberg, D. Health Impact of Childhood and Adolescent Soy Consumption. Nutr. Rev. 2017, 75, 500–515. [Google Scholar] [CrossRef]
- Russo, M.; Russo, G.L.; Daglia, M.; Kasi, P.D.; Ravi, S.; Nabavi, S.F.; Nabavi, S.M. Understanding Genistein in Cancer: The “Good” and the “Bad” Effects: A Review. Food Chem. 2016, 196, 589–600. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy Isoflavones and Prostate Cancer: A Review of Molecular Mechanisms. J. Steroid Biochem. Mol. Biol. 2014, 140, 116–132. [Google Scholar] [CrossRef]
- Douglas, C.; Johnson, S.; Arjmandi, B. Soy and Its Isoflavones: The Truth Behind the Science in Breast Cancer. Anti-Cancer Agents Med. Chem. 2013, 13, 1178–1187. [Google Scholar] [CrossRef]
- Zhao, T.T.; Jin, F.; Li, J.G.; Xu, Y.Y.; Dong, H.T.; Liu, Q.; Xing, P.; Zhu, G.L.; Xu, H.; Miao, Z.F. Dietary Isoflavones or Isoflavone-Rich Food Intake and Breast Cancer Risk: A Meta-Analysis of Prospective Cohort Studies. Clin. Nutr. 2019, 38, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Oseni, T.; Patel, R.; Pyle, J.; Jordan, V.C. Selective Estrogen Receptor Modulators and Phytoestrogens. Planta Med. 2008, 74, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Varinska, L.; Gal, P.; Mojzisova, G.; Mirossay, L.; Mojzis, J. Soy and Breast Cancer: Focus on Angiogenesis. Int. J. Mol. Sci. 2015, 16, 11728–11749. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Li, Y.; Wang, Z.; Sarkar, F.H. Multi-Targeted Therapy of Cancer by Genistein. Cancer Lett. 2008, 269, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.C.; Peng, S.F.; Lai, K.C.; Liao, C.L.; Huang, Y.P.; Lin, C.C.; Lin, M.L.; Liu, K.C.; Tsai, C.C.; Ma, Y.S.; et al. Genistein Induces Apoptosis in Vitro and Has Antitumor Activity against Human Leukemia HL-60 Cancer Cell Xenograft Growth in Vivo. Environ. Toxicol. 2019, 34, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Ma, J.; Sun, J.; Yang, L.; Yang, F.; Zhang, W.; Li, R.; Wang, L.; Wang, Y.; Wang, H. Genistein and AG1024 Synergistically Increase the Radiosensitivity of Prostate Cancer Cells. Oncol. Rep. 2018, 40, 579–588. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Y.; Gu, J.; Liang, P.; Shen, M.; Xi, J.; Qin, J. Anti-Invasive Effect and Pharmacological Mechanism of Genistein against Colorectal Cancer. Biofactors 2020, 46, 620–628. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and Cancer: Current Status, Challenges, and Future Directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef]
- Naujokat, C.; McKee, D.L. The “Big Five” Phytochemicals Targeting Cancer Stem Cells: Curcumin, EGCG, Sulforaphane, Resveratrol and Genistein. Curr. Med. Chem. 2020, 28, 4321–4342. [Google Scholar] [CrossRef]
- Yu, X.; Zhu, J.; Mi, M.; Chen, W.; Pan, Q.; Wei, M. Anti-Angiogenic Genistein Inhibits VEGF-Induced Endothelial Cell Activation by Decreasing PTK Activity and MAPK Activation. Med. Oncol. 2012, 29, 349–357. [Google Scholar] [CrossRef]
- Büchler, P.; Reber, H.A.; Büchler, M.W.; Friess, H.; Lavey, R.S.; Hines, O.J. Antiangiogenic Activity of Genistein in Pancreatic Carcinoma Cells Is Mediated by the Inhibition of Hypoxia-Inducible Factor-1 and the down-Regulation of VEGF Gene Expression. Cancer 2004, 100, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y. Soy Isoflavones and Cancer Prevention. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H. Flavonoids Inhibit VEGF/BFGF-Induced Angiogenesis in Vitro by Inhibiting the Matrix-Degrading Proteases. J. Cell. Biochem. 2003, 89, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Su, S.-J.; Yeh, T.-M.; Chuang, W.-J.; Ho, C.-L.; Chang, K.-L.; Cheng, H.-L.; Liu, H.-S.; Cheng, H.-L.; Hsu, P.-Y.; Chow, N.-H. The Novel Targets for Anti-Angiogenesis of Genistein on Human Cancer Cells. Biochem. Pharmacol. 2005, 69, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, B.; Hammarström, C.; Yang, J.; Ramberg, H.; Diep, L.M.; Karlsen, S.J.; Kucuk, O.; Saatcioglu, F.; Taskèn, K.A.; Svindland, A. The Effects of Short-Term Genistein Intervention on Prostate Biomarker Expression in Patients with Localised Prostate Cancer before Radical Prostatectomy. Br. J. Nutr. 2012, 108, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Messing, E.; Gee, J.R.; Saltzstein, D.R.; Kim, K.; DiSant’Agnese, A.; Kolesar, J.; Harris, L.; Faerber, A.; Havighurst, T.; Young, J.M.; et al. A Phase 2 Cancer Chemoprevention Biomarker Trial of Isoflavone G-2535 (Genistein) in Presurgical Bladder Cancer Patients. Cancer Prev. Res. 2013, 5, 621–630. [Google Scholar] [CrossRef] [PubMed]
- El-Rayes, B.F.; Philip, P.A.; Sarkar, F.H.; Shields, A.F.; Ferris, A.M.; Hess, K.; Kaseb, A.O.; Javle, M.M.; Varadhachary, G.R.; Wolff, R.A.; et al. A Phase II Study of Isoflavones, Erlotinib, and Gemcitabine in Advanced Pancreatic Cancer. Investig. New Drugs 2011, 29, 694–699. [Google Scholar] [CrossRef]
- Khan, S.A.; Chatterton, R.T.; Michel, N.; Bryk, M.; Lee, O.; Ivancic, D.; Heinz, R.; Zalles, C.M.; Helenowski, I.; Jovanovic, B.; et al. Soy Isoflavone Supplementation for Breast Cancer Risk Reduction: A Randomized Phase II Trial. Cancer Prev. Res. 2012, 5, 309–319. [Google Scholar] [CrossRef]
- Pintova, S.; Dharmupari, S.; Moshier, E.; Zubizarreta, N.; Ang, C.; Holcombe, R.F. Genistein Combined with FOLFOX or FOLFOX–Bevacizumab for the Treatment of Metastatic Colorectal Cancer: Phase I/II Pilot Study. Cancer Chemother. Pharmacol. 2019, 84, 591–598. [Google Scholar] [CrossRef]
- Kim, T.L.; Jeong, G.H.; Yang, J.W.; Lee, K.H.; Kronbichler, A.; van der Vliet, H.J.; Grosso, G.; Galvano, F.; Aune, D.; Kim, J.Y.; et al. Tea Consumption and Risk of Cancer: An Umbrella Review and Meta-Analysis of Observational Studies. Adv. Nutr. 2020, 11, 1437–1452. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Xu, Q.; Lu, J.; Wang, P.; Zhang, H.W.; Zhou, L.; Ma, X.Q.; Zhou, Y.H. Tea Consumption and the Incidence of Cancer: A Systematic Review and Meta-Analysis of Prospective Observational Studies. Eur. J. Cancer Prev. 2015, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo, E.; Medina, M.Á.; Sánchez-Jiménez, F.; Urdiales, J.L. Targeting of Histamine Producing Cells by EGCG: A Green Dart against Inflammation? J. Physiol. Biochem. 2010, 66, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green Tea Catechin, Epigallocatechin-3-Gallate (EGCG): Mechanisms, Perspectives and Clinical Applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.W.; Wei, C.; Lung, W.Y.; Wei, X.Y.; Cheng, B.H.; Cai, Z.M.; Huang, W.R. EGCG Inhibited Bladder Cancer SW780 Cell Proliferation and Migration Both in Vitro and in Vivo via Down-Regulation of NF-ΚB and MMP-9. J. Nutr. Biochem. 2017, 41, 56–64. [Google Scholar] [CrossRef]
- Neuhaus, T.; Pabst, S.; Stier, S.; Weber, A.A.; Schrör, K.; Sachinidis, A.; Vetter, H.; Ko, Y.D. Inhibition of the Vascular-Endothelial Growth Factor-Induced Intracellular Signaling and Mitogenesis of Human Endothelial Cells by Epigallocatechin-3 Gallate. Eur. J. Pharmacol. 2004, 483, 223–227. [Google Scholar] [CrossRef]
- Moyle, C.W.A.; Cerezo, A.B.; Winterbone, M.S.; Hollands, W.J.; Alexeev, Y.; Needs, P.W.; Kroon, P.A. Potent Inhibition of VEGFR-2 Activation by Tight Binding of Green Tea Epigallocatechin Gallate and Apple Procyanidins to VEGF: Relevance to Angiogenesis. Mol. Nutr. Food Res. 2015, 59, 401–412. [Google Scholar] [CrossRef]
- Rashidi, B.; Malekzadeh, M.; Goodarzi, M.; Masoudifar, A.; Mirzaei, H. Green Tea and Its Anti-Angiogenesis Effects. Biomed. Pharmacother. 2017, 89, 949–956. [Google Scholar] [CrossRef]
- Li, X.; Feng, Y.; Liu, J.; Feng, X.; Zhou, K.; Tang, X. Epigallocatechin-3-Gallate Inhibits IGF-I-Stimulated Lung Cancer Angiogenesis through Downregulation of HIF-1a and VEGF Expression. Mol. Nutr. Food Res. 2013, 6, 169–178. [Google Scholar] [CrossRef]
- Gu, J.-W.; Makey, K.L.; Tucker, K.B.; Chinchar, E.; Mao, X.; Pei, I.; Thomas, E.Y.; Miele, L. EGCG, a Major Green Tea Catechin Suppresses Breast Tumor Angiogenesis and Growth via Inhibiting the Activation of HIF-1α and NFκB, and VEGF Expression. Vasc. Cell 2013, 5, 9. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, Y.J.; Wang, C.C.N.; Lan, Y.H.; Lan, S.J.; Sheu, M.J. Epigallocatechin-3-Gallate Inhibits Tumor Angiogenesis: Involvement of Endoglin/Smad1 Signaling in Human Umbilical Vein Endothelium Cells. Biomed. Pharmacother. 2019, 120, 109491. [Google Scholar] [CrossRef]
- Ohga, N.; Hida, K.; Hida, Y.; Muraki, C.; Tsuchiya, K.; Matsuda, K.; Ohiro, Y.; Totsuka, Y.; Shindoh, M. Inhibitory Effects of Epigallocatechin-3 Gallate, a Polyphenol in Green Tea, on Tumor-Associated Endothelial Cells and Endothelial Progenitor Cells. Cancer Sci. 2009, 100, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Mereles, D.; Hunstein, W. Epigallocatechin-3-Gallate (EGCG) for Clinical Trials: More Pitfalls than Promises? Int. J. Mol. Sci. 2011, 12, 5592–5603. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Adhami, V.M.; Mukhtar, H. Apoptosis by Dietary Agents for Prevention and Treatment of Prostate Cancer. Endocr. Relat. Cancer 2010, 17, R39–R52. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Varoni, E.M. Chemopreventive Potential of Flavonoids in Oral Squamous Cell Carcinoma in Human Studies. Nutrients 2013, 5, 2564–2576. [Google Scholar] [CrossRef]
- Kumar, N.B.; Pow-Sang, J.; Spiess, P.E.; Park, J.; Salup, R.; Williams, C.R.; Parnes, H.; Schell, M.J. Randomized, Placebo-Controlled Trial Evaluating the Safety of One-Year Administration of Green Tea Catechins. Oncotarget 2016, 7, 70794–70802. [Google Scholar] [CrossRef]
- Gescher, A.; Steward, W.P.; Brown, K. Resveratrol in the Management of Human Cancer: How Strong Is the Clinical Evidence? Ann. N. Y. Acad. Sci. 2013, 1290, 12–20. [Google Scholar] [CrossRef]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Sanchez-Rovira, P.; Ramirez-Tortosa, M.C. Hydroxytyrosol: From Laboratory Investigations to Future Clinical Trials. Nutr. Rev. 2010, 68, 191–206. [Google Scholar] [CrossRef]
- Yang, X.; Li, X.; Ren, J. From French Paradox to Cancer Treatment: Anti-Cancer Activities and Mechanisms of Resveratrol. Anti-Cancer Agents Med. Chem. 2014, 14, 806–825. [Google Scholar] [CrossRef]
- Li, D.; Wang, G.; Jin, G.; Yao, K.; Zhao, Z.; Bie, L.; Guo, Y.; Li, N.; Deng, W.; Chen, X.; et al. Resveratrol Suppresses Colon Cancer Growth by Targeting the AKT/STAT3 Signaling Pathway. Int. J. Mol. Med. 2019, 43, 630–640. [Google Scholar] [CrossRef]
- Park, S.Y.; Jeong, K.J.; Lee, J.; Yoon, D.S.; Choi, W.S.; Kim, Y.K.; Han, J.W.; Kim, Y.M.; Kim, B.K.; Lee, H.Y. Hypoxia Enhances LPA-Induced HIF-1α and VEGF Expression: Their Inhibition by Resveratrol. Cancer Lett. 2007, 258, 63–69. [Google Scholar] [CrossRef]
- Yu, H.-B.; Zhang, H.-F.; Zhang, X.; Li, D.-Y.; Xue, H.-Z.; Pan, C.-E.; Zhao, S.-H. Resveratrol Inhibits VEGF Expression of Human Hepatocellular Carcinoma Cells through a NF-Kappa B-Mediated Mechanism. Hepato-Gastroenterol. 2010, 57, 1241–1246. [Google Scholar]
- Trapp, V.; Parmakhtiar, B.; Papazian, V.; Willmott, L.; Fruehauf, J.P. Anti-Angiogenic Effects of Resveratrol Mediated by Decreased VEGF and Increased TSP1 Expression in Melanoma-Endothelial Cell Co-Culture. Angiogenesis 2010, 13, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Igura, K.; Ohta, T.; Kuroda, Y.; Kaji, K. Resveratrol and Quercetin Inhibit Angiogenesis in Vitro. Cancer Lett. 2001, 171, 11–16. [Google Scholar] [CrossRef]
- Bråkenhielm, E.; Cao, R.; Cao, Y. Suppression of Angiogenesis, Tumor Growth, and Wound Healing by Resveratrol, a Natural Compound in Red Wine and Grapes. FASEB J. 2001, 15, 1798–1800. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, H.; Zou, Y.; Liu, Q.; Guo, C.; Gao, G.; Shao, C.; Gong, Y. Resveratrol Modulates Angiogenesis through the GSK3β/β-Catenin/TCF-Dependent Pathway in Human Endothelial Cells. Biochem. Pharmacol. 2010, 80, 1386–1395. [Google Scholar] [CrossRef]
- Kasiotis, K.M.; Pratsinis, H.; Kletsas, D.; Haroutounian, S.A. Resveratrol and Related Stilbenes: Their Anti-Aging and Anti-Angiogenic Properties. Food Chem. Toxicol. 2013, 61, 112–120. [Google Scholar] [CrossRef]
- Carter, L.G.; D’Orazio, J.A.; Pearson, K.J. Resveratrol and Cancer: Focus on in Vivo Evidence. Endocr. Relat. Cancer 2014, 21, R209–R225. [Google Scholar] [CrossRef]
- Ko, J.H.; Sethi, G.; Um, J.Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef]
- Sinha, D.; Sarkar, N.; Biswas, J.; Bishayee, A. Resveratrol for Breast Cancer Prevention and Therapy: Preclinical Evidence and Molecular Mechanisms. Semin. Cancer Biol. 2016, 40, 209–232. [Google Scholar] [CrossRef]
- Yousef, M.; Vlachogiannis, I.A.; Tsiani, E. Effects of Resveratrol against Lung Cancer: In Vitro and In Vivo Studies. Nutrients 2017, 9, 1231. [Google Scholar] [CrossRef]
- Singh, C.K.; Ndiaye, M.A.; Ahmad, N. Resveratrol and Cancer: Challenges for Clinical Translation. Biochim. Biophys. Acta 2015, 1852, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Novelle, M.G.; Wahl, D.; Diéguez, C.; Bernier, M.; de Cabo, R. Resveratrol Supplementation: Where Are We Now and Where Should We Go? Ageing Res. Rev. 2015, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fortes, C.; García-Vilas, J.A.; Quesada, A.R.; Medina, M.Á. Evaluation of the Anti-Angiogenic Potential of Hydroxytyrosol and Tyrosol, Two Bio-Active Phenolic Compounds of Extra Virgin Olive Oil, in Endothelial Cell Cultures. Food Chem. 2012, 134, 134–140. [Google Scholar] [CrossRef]
- García-Vilas, J.A.; Quesada, A.R.; Medina, M.Á. Hydroxytyrosol Targets Extracellular Matrix Remodeling by Endothelial Cells and Inhibits Both Ex Vivo and in Vivo Angiogenesis. Food Chem. 2017, 221, 1741–1746. [Google Scholar] [CrossRef] [PubMed]
- López-Jiménez, A.; Gallardo, E.; Espartero, J.L.; Madrona, A.; Quesada, A.R.; Medina, M.Á. Comparison of the Anti-Angiogenic Potential of Hydroxytyrosol and Five Derivatives. Food Funct. 2018, 9, 4310–4316. [Google Scholar] [CrossRef]
- Marrero, A.D.; Castilla, L.; Espartero, J.L.; Madrona, A.; R. Quesada, A.; Medina, M.Á.; Martínez-Poveda, B. A Comparative Study of the Antiangiogenic Activity of Hydroxytyrosyl Alkyl Ethers. Food Chem. 2020, 333, 127476. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, R. Estrogenic Terpenes and Terpenoids: Pathways, Functions and Applications. Eur. J. Pharmacol. 2017, 815, 405–415. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic Triterpene Distribution in Various Plants—Rich Sources for a New Group of Multi-Potent Plant Extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef]
- Kondo, M.; Mackinnon, S.L.; Craft, C.C.; Matchett, M.D.; Hurta, R.A.R.; Neto, C.C. Ursolic Acid and Its Esters: Occurrence in Cranberries and Other Vaccinium Fruit and Effects on Matrix Metalloproteinase Activity in DU145 Prostate Tumor Cells. J. Sci. Food Agric. 2011, 91, 789–796. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Sharma, A.K. Ursolic Acid (UA): A Metabolite with Promising Therapeutic Potential. Life Sci. 2016, 146, 201–213. [Google Scholar] [CrossRef]
- Cargnin, S.T.; Gnoatto, S.B. Ursolic Acid from Apple Pomace and Traditional Plants: A Valuable Triterpenoid with Functional Properties. Food Chem. 2017, 220, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Majumdar, S.; Banerjee, S.; Aggarwal, B.B. Ursolic Acid Inhibits Nuclear Factor- κ B Activation Induced by Carcinogenic Agents through Suppression of IκBα Kinase and P65 Phosphorylation: Correlation with Down-Regulation of Cyclooxygenase 2, Matrix Metalloproteinase 9, and Cyclin D1. Cancer Res. 2003, 63, 4375–4383. [Google Scholar] [PubMed]
- Li, J.; Liang, X.; Yang, X. Ursolic Acid Inhibits Growth and Induces Apoptosis in Gemcitabine-Resistant Human Pancreatic Cancer via the JNK and PI3K/Akt/NF-ΚB Pathways. Oncol. Rep. 2012, 28, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Ong, T.H.; Kumar, A.P.; Lun, C.K.; Ho, P.C.; Wong, P.T.H.; Hui, K.M.; Sethi, G. Ursolic Acid Inhibits the Initiation, Progression of Prostate Cancer and Prolongs the Survival of TRAMP Mice by Modulating Pro-Inflammatory Pathways. PLoS ONE 2012, 7, e32476. [Google Scholar] [CrossRef]
- Prasad, S.; Yadav, V.R.; Sung, B.; Reuter, S.; Kannappan, R.; Deorukhkar, A.; Diagaradjane, P.; Wei, C.; Baladandayuthapani, V.; Krishnan, S.; et al. Ursolic Acid Inhibits Growth and Metastasis of Human, Colorectal Cancer in an Orthotopic Nude Mouse Model by Targeting Multiple Cell Signaling Pathways: Chemosensitization with Capecitabine. Clin. Cancer Res. 2012, 18, 4942–4953. [Google Scholar] [CrossRef]
- Prasad, S.; Yadav, V.R.; Sung, B.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Ursolic Acid Inhibits the Growth of Human Pancreatic Cancer and Enhances the Antitumor Potential of Gemcitabine in an Orthotopic Mouse Model through Suppression of the Inflammatory Microenvironment. Oncotarget 2016, 7, 13182–13196. [Google Scholar] [CrossRef]
- Ovesná, Z.; Vachálková, A.; Horváthová, K.; Tóthová, D. Pentacyclic Triterpenoic Acids: New Chemoprotective Compounds. Neoplasma 2004, 51, 327–333. [Google Scholar]
- Wang, W.; Zhao, C.; Jou, D.; Lü, J.; Zhang, C.; Lin, L.I.; Lin, J. Ursolic Acid Inhibits the Growth of Colon Cancer-Initiating Cells by Targeting STAT3. Anticancer Res. 2013, 33, 4279–4284. [Google Scholar]
- Lin, J.; Chen, Y.; Wei, L.; Hong, Z.; Sferra, T.J.; Peng, J. Ursolic Acid Inhibits Colorectal Cancer Angiogenesis through Suppression of Multiple Signaling Pathways. Int. J. Oncol. 2013, 43, 1666–1674. [Google Scholar] [CrossRef]
- Lin, C.C.; Huang, C.Y.; Mong, M.C.; Chan, C.Y.; Yin, M.C. Antiangiogenic Potential of Three Triterpenic Acids in Human Liver Cancer Cells. J. Agric. Food Chem. 2011, 59, 755–762. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.Á. Effects of Ursolic Acid on Different Steps of the Angiogenic Process. Biochem. Biophys. Res. Commun. 2004, 320, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kiran, M.S.; Viji, R.I.; Sameer Kumar, V.B.; Sudhakaran, P.R. Modulation of Angiogenic Factors by Ursolic Acid. Biochem. Biophys. Res. Commun. 2008, 371, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Saraswati, S.; Agrawal, S.S.; Alhaider, A.A. Ursolic Acid Inhibits Tumor Angiogenesis and Induces Apoptosis through Mitochondrial-Dependent Pathway in Ehrlich Ascites Carcinoma Tumor. Chem. Biol. Interact. 2013, 206, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Dai, X.; Kumar, A.P.; Tan, B.K.H.; Sethi, G.; Bishayee, A. Ursolic Acid in Cancer Prevention and Treatment: Molecular Targets, Pharmacokinetics and Clinical Studies. Biochem. Pharmacol. 2013, 85, 1579–1587. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.Á. Insights on the Antitumor Effects of Kahweol on Human Breast Cancer: Decreased Survival and Increased Production of Reactive Oxygen Species and Cytotoxicity. Biochem. Biophys. Res. Commun. 2014, 447, 452–458. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef]
- Wang, S.; Yoon, Y.C.; Sung, M.J.; Hur, H.J.; Park, J.H. Antiangiogenic Properties of Cafestol, a Coffee Diterpene, in Human Umbilical Vein Endothelial Cells. Biochem. Biophys. Res. Commun. 2012, 421, 567–571. [Google Scholar] [CrossRef]
- Moeenfard, M.; Cortez, A.; Machado, V.; Costa, R.; Luís, C.; Coelho, P.; Soares, R.; Alves, A.; Borges, N.; Santos, A. Anti-Angiogenic Properties of Cafestol and Kahweol Palmitate Diterpene Esters. J. Cell. Biochem. 2016, 117, 2748–2756. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.Á. Anti-Angiogenic and Anti-Inflammatory Properties of Kahweol, a Coffee Diterpene. PLoS ONE 2011, 6, e23407. [Google Scholar] [CrossRef]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Chen, P.Y.; Cheng, T.H.; Chao, H.H.; Liu, J.C.; Chen, J.J. Cafestol Inhibits Cyclic-Strain-Induced Interleukin-8, Intercellular Adhesion Molecule-1, and Monocyte Chemoattractant Protein-1 Production in Vascular Endothelial Cells. Oxidative Med. Cell. Longev. 2018, 2018, 7861518. [Google Scholar] [CrossRef]
- Cascão, R.; Fonseca, J.E.; Moita, L.F. Celastrol: A Spectrum of Treatment Opportunities in Chronic Diseases. Front. Med. 2017, 4, 69. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Mukherjee, T.; Bishayee, A. Molecular Targets of Celastrol in Cancer: Recent Trends and Advancements. Crit. Rev. Oncol./Hematol. 2018, 128, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhou, Y.; Fan, Y.; Zhou, D. Celastrol Inhibits the Growth of Human Glioma Xenografts in Nude Mice through Suppressing VEGFR Expression. Cancer Lett. 2008, 264, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Yi, Z.; Zhang, J.; Lu, B.; Sung, B.; Qu, W.; Aggarwal, B.B.; Liu, M. Celastrol Suppresses Angiogenesis-Mediated Tumor Growth through Inhibition of AKT/Mammalian Target of Rapamycin Pathway. Cancer Res. 2010, 70, 1951–1959. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, Z.; Zhang, S.; Ren, J.; Zhang, R.; Zeng, H.; Li, Q.; Wu, G. Inhibitory Action of Celastrol on Hypoxia-Mediated Angiogenesis and Metastasis via the HIF-1α Pathway. Int. J. Mol. Med. 2011, 27, 407–415. [Google Scholar] [CrossRef]
- Ni, H.; Zhao, W.; Kong, X.; Li, H.; Ouyang, J. Celastrol Inhibits Lipopolysaccharide-Induced Angiogenesis by Suppressing TLR4-Triggered Nuclear Factor-Kappa B Activation. Acta Haematol. 2014, 131, 102–111. [Google Scholar] [CrossRef]
- López-Jiménez, A.; García-Caballero, M.; Medina, M.Á.; Quesada, A.R. Anti-Angiogenic Properties of Carnosol and Carnosic Acid, Two Major Dietary Compounds from Rosemary. Eur. J. Nutr. 2013, 52, 85–95. [Google Scholar] [CrossRef]
- Kashyap, D.; Kumar, G.; Sharma, A.; Sak, K.; Tuli, H.S.; Mukherjee, T.K. Mechanistic Insight into Carnosol-Mediated Pharmacological Effects: Recent Trends and Advancements. Life Sci. 2017, 169, 27–36. [Google Scholar] [CrossRef]
- Malik, E.M.; Müller, C.E. Anthraquinones As Pharmacological Tools and Drugs. Med. Res. Rev. 2016, 36, 705–748. [Google Scholar] [CrossRef]
- Sánchez, M.; González-Burgos, E.; Iglesias, I.; Gómez-Serranillos, M.P. Pharmacological Update Properties of Aloe Vera and Its Major Active Constituents. Molecules 2020, 25, 1324. [Google Scholar] [CrossRef]
- Hsu, S.C.; Chung, J.G. Anticancer Potential of Emodin. BioMedicine 2012, 2, 108–116. [Google Scholar] [CrossRef]
- Liu, A.; Chen, H.; Wei, W.; Ye, S.; Liao, W.; Gong, J.; Jiang, Z.; Wang, L.; Lin, S. Antiproliferative and Antimetastatic Effects of Emodin on Human Pancreatic Cancer. Oncol. Rep. 2011, 26, 81–89. [Google Scholar] [CrossRef]
- Ok, S.; Kim, S.-M.; Kim, C.; Nam, D.; Shim, B.S.; Kim, S.-H.; Ahn, K.S.; Choi, S.-H.; Ahn, K.S. Emodin Inhibits Invasion and Migration of Prostate and Lung Cancer Cells by Downregulating the Expression of Chemokine Receptor CXCR4. Immunopharmacol. Immunotoxicol. 2012, 34, 768–778. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, M.S.; Cha, E.Y.; Sul, J.Y.; Lee, J.S.; Kim, J.S.; Park, J.B.; Kim, J.Y. Inhibitory Effect of Emodin on Fatty Acid Synthase, Colon Cancer Proliferation and Apoptosis. Mol. Med. Rep. 2017, 15, 2163–2173. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Cui, C.; Yang, L.; Wang, L.; Jiang, X. Emodin Inhibits Colon Cancer Cell Invasion and Migration by Suppressing Epithelialmesenchymal Transition via the Wnt/β-Catenin Pathway. Oncol. Res. 2018, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Saunders, I.T.; Mir, H.; Kapur, N.; Singh, S. Emodin Inhibits Colon Cancer by Altering BCL-2 Family Proteins and Cell Survival Pathways. Cancer Cell Int. 2019, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Dong, T.; Li, R.; Lu, J.; Wei, X.; Liu, P. Emodin Inhibits Epithelial to Mesenchymal Transition in Epithelial Ovarian Cancer Cells by Regulation of GSK-3β/β-Catenin/ZEB1 Signaling Pathway. Oncol. Rep. 2016, 35, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.T.; Lin, S.Z.; Liu, D.L.; Wang, Z.H. The Distinct Mechanisms of the Antitumor Activity of Emodin in Different Types of Cancer (Review). Oncol. Rep. 2013, 30, 2555–2562. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, G.; Babykutty, S.; Sathiadevan, P.P.; Srinivas, P. Molecular Mechanism of Emodin Action: Transition from Laxative Ingredient to an Antitumor Agent. Med. Res. Rev. 2007, 27, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Kaneshiro, T.; Morioka, T.; Inamine, M.; Kinjo, T.; Arakaki, J.; Chiba, I.; Sunagawa, N.; Suzui, M.; Yoshimi, N. Anthraquinone Derivative Emodin Inhibits Tumor-Associated Angiogenesis through Inhibition of Extracellular Signal-Regulated Kinase 1/2 Phosphorylation. Eur. J. Pharmacol. 2006, 553, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, X.; Zhou, Q.; Lu, Y.; Zhang, H.; Chen, Q.; Zhao, M.; Su, S. Inhibitory Effect of Emodin on Migration, Invasion and Metastasis of Human Breast Cancer MDA-MB-231 Cells in Vitro and in Vivo. Oncol. Rep. 2015, 33, 338–346. [Google Scholar] [CrossRef]
- Lin, S.-Z.; Wei, W.-T.; Chen, H.; Chen, K.-J.; Tong, H.-F.; Wang, Z.-H.; Ni, Z.-L.; Liu, H.-B.; Guo, H.-C.; Liu, D.-L. Antitumor Activity of Emodin against Pancreatic Cancer Depends on Its Dual Role: Promotion of Apoptosis and Suppression of Angiogenesis. PLoS ONE 2012, 7, e42146. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Z.; Xu, J.B.; Ji, X.; Chen, H.; Xu, H.T.; Hu, P.; Chen, L.; Guo, J.Q.; Chen, M.Y.; Lu, D.; et al. Emodin Inhibits Angiogenesis in Pancreatic Cancer by Regulating the Transforming Growth Factor-β/Drosophila Mothers against Decapentaplegic Pathway and Angiogenesis-Associated MicroRNAs. Mol. Med. Rep. 2015, 12, 5865–5871. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lu, H.; Wang, S.; Chen, B.; Liu, Z.; Ke, X.; Liu, T.; Fu, J. The Anthraquinone Derivative Emodin Inhibits Angiogenesis and Metastasis through Downregulating Runx2 Activity in Breast Cancer. Int. J. Oncol. 2015, 46, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Park, Y.H.; Yang, E.Y.; Park, W.S.; Park, K.S. Inhibition of Tamoxifen’s Therapeutic Effects by Emodin in Estrogen Receptor-Positive Breast Cancer Cell Lines. Ann. Surg. Treat. Res. 2019, 97, 230. [Google Scholar] [CrossRef]
- Pecere, T.; Gazzola, M.V.; Mucignat, C.; Tumors, N.; Parolin, C.; Vecchia, F.D.; Cavaggioni, A.; Basso, G.; Diaspro, A.; Salvato, B.; et al. Aloe-Emodin Is a New Type of Anticancer Agent with Selective Activity against Neuroectodermal Tumors. Cancer Res. 2000, 60, 2800–2804. [Google Scholar]
- Cárdenas, C.; Quesada, A.R.; Medina, M.Á. Evaluation of the Anti-Angiogenic Effect of Aloe-Emodin. Cell. Mol. Life Sci. 2006, 63, 3083–3089. [Google Scholar] [CrossRef]
- Suboj, P.; Babykutty, S.; Valiyaparambil Gopi, D.R.; Nair, R.S.; Srinivas, P.; Gopala, S. Aloe Emodin Inhibits Colon Cancer Cell Migration/Angiogenesis by Downregulating MMP-2/9, RhoB and VEGF via Reduced DNA Binding Activity of NF-ΚB. Eur. J. Pharm. Sci. 2012, 45, 581–591. [Google Scholar] [CrossRef]
- Wu, J.M.; Ke, X.; Wang, W.; Zhang, H.; Ma, N.; Fu, W.; Zhao, M.; Gao, X.; Hao, X.; Zhang, Z. Aloe-Emodin Suppresses Hypoxia-Induced Retinal Angiogenesis via Inhibition of HIF-1α/VEGF Pathway. Int. J. Biol. Sci. 2016, 12, 1363–1371. [Google Scholar] [CrossRef]
- Lin, L.X.; Wang, P.; Wang, Y.T.; Huang, Y.; Jiang, L.; Wang, X.M. Aloe Vera and Vitis Vinifera Improve Wound Healing in an in Vivo Rat Burn Wound Model. Mol. Med. Rep. 2016, 13, 1070–1076. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Description | References | |
|---|---|---|---|
| In vitro | Proliferation/survival (MTT) | Angiogenesis depends directly on the active proliferation of EC. The MTT assay measures cellular viability based on the reduction of soluble MTT(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) to a blue-colored tetrazolium salt by mitochondrial reductases, followed by detection in a spectrophotometer. | [20,21,22] |
| Tubular-like structure formation | Reorganization of EC to create tubular-like networks resembling blood vessel formation. Matrigel, a solubilized murine extract of basal lamina, is used as a substrate. | [21] | |
| Migration/Invasion | Migration throughout the ECM and its degradation are the main capacities for EC to form new vessels. The migration and invasion fluorometric assays are based on the disposal of fluorescence-labelled EC on a transwell system that allows the movement of EC towards chemotactic stimuli contained in the media in the well. Specific to the invasion assay, a thin layer of Matrigel is placed over the filter of the transwell. Afterward, the light emitted by fluorescence-labeled cells is measured. | [21] | |
| Zymography (detection of gelatinase and caseinase activities) | Matrix metalloproteinases (MMPs) and serin proteases degrade the ECM to ease the rearrangement of EC to form new vessels. Finding inhibitors of these enzymes can limit the angiogenic process. The detection of their activity involves the electrophoresis of the secreted protease enzymes through discontinuous nondenaturing polyacrylamide gels containing enzyme’s substrate (either type III gelatin or β-casein). Staining the gel with a protein dye allows the detection of the proteolytic activity as clear bands of lysis against a stained background. | [21,23] | |
| Ex vivo | Mouse aortic ring | Culture of transversal sections of rat aorta placed on Matrigel induces the outgrowth of the ring at day 7–14 that can be observed in the microscope. | [24] |
| In vivo | Chorioallantoic membrane | The strong blood irrigation of the chorioallantoic membrane makes it an economic, feasible model to study angiogenesis. Methylcellulose discs containing the studied compounds can be directly placed on the membrane to test their effect on embryonic angiogenesis. | [20,25,26] |
| Zebrafish intersegmental vessel models | Zebrafish is currently a model system for the study of angiogenesis during its embryonic development. After being treated with the compounds to test, the circulatory system of the embryos (transparent) can be easily observed by microscopy or binocular scopes, and abnormalities in the development can be assessed. The detection can be improved by in situ hybridizations, among other approaches. | [27,28,29,30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrero, A.D.; Quesada, A.R.; Martínez-Poveda, B.; Medina, M.Á. Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer. Antioxidants 2022, 11, 302. https://doi.org/10.3390/antiox11020302
Marrero AD, Quesada AR, Martínez-Poveda B, Medina MÁ. Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer. Antioxidants. 2022; 11(2):302. https://doi.org/10.3390/antiox11020302
Chicago/Turabian StyleMarrero, Ana Dácil, Ana R. Quesada, Beatriz Martínez-Poveda, and Miguel Ángel Medina. 2022. "Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer" Antioxidants 11, no. 2: 302. https://doi.org/10.3390/antiox11020302
APA StyleMarrero, A. D., Quesada, A. R., Martínez-Poveda, B., & Medina, M. Á. (2022). Antiangiogenic Phytochemicals Constituent of Diet as Promising Candidates for Chemoprevention of Cancer. Antioxidants, 11(2), 302. https://doi.org/10.3390/antiox11020302