
The Effects of Nuclear Factor Erythroid 2 (NFE2)-Related Factor 2 (Nrf2) Activation in Preclinical Models of Peripheral Neuropathic Pain




Abstract
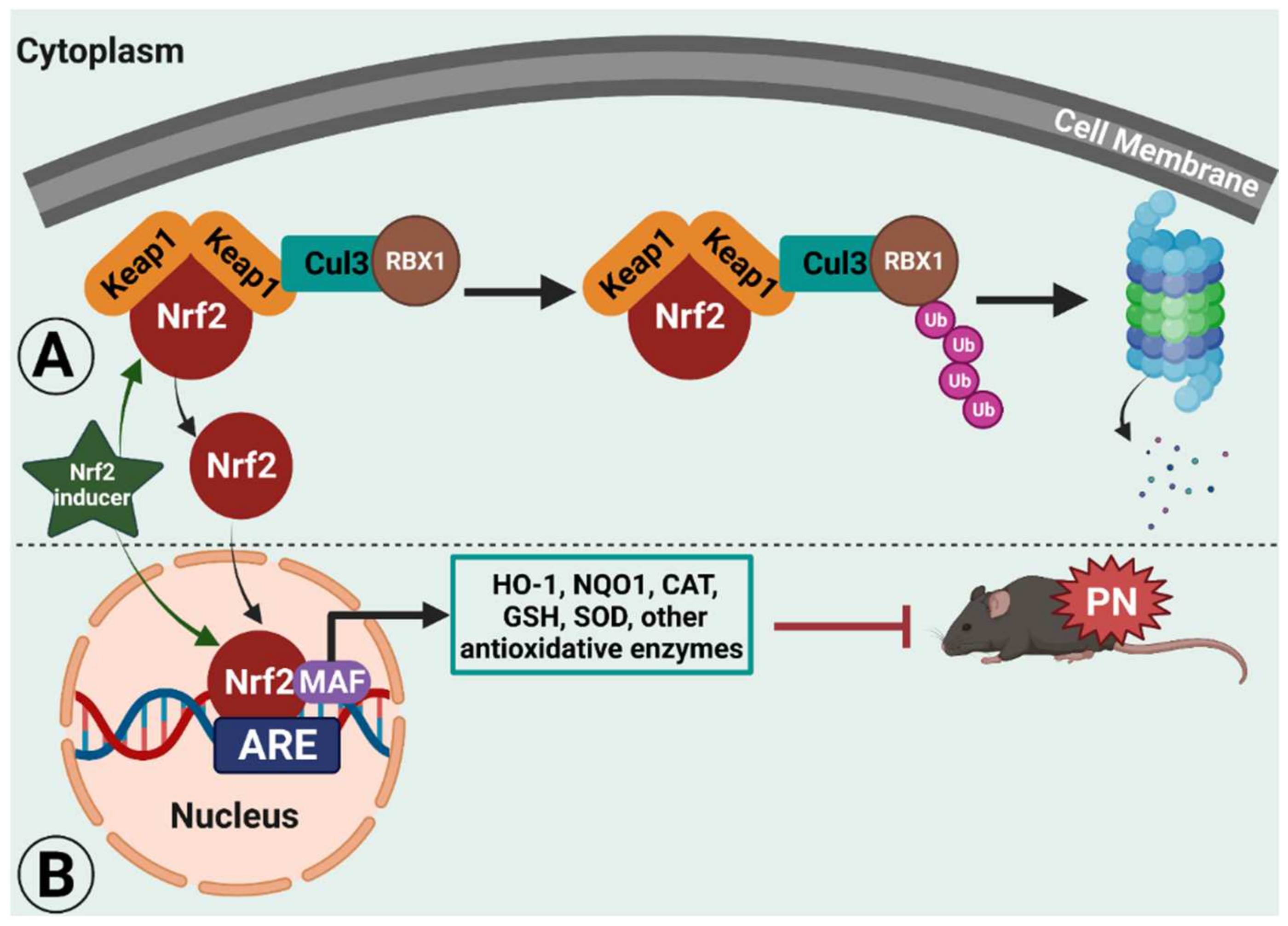
:1. Peripheral Neuropathic Pain and Erythroid 2 (NFE2)-Related Factor 2 (Nrf2)
2. Nrf2 Signaling and Its Inducers in Different Peripheral Neuropathic Pain Conditions
2.1. Diabetic Neuropathy (DN)

{kind=link}
| Nrf2 Inducer | Animals (Sex, Strain) | Dose (mg/kg) and Administration Route | Mechanism of Action | Reference |
|---|---|---|---|---|
| Resveratrol | Nrf2−/− and Nrf2+/+ CD1/ICR mice | 10 mL/kg 10%, intragastric | Nrf2 pathway | [67] |
| Polydatin | Male Sprague Dawley rats | 25 and 50 mg/kg, oral 10, 20 µM in neuro2a—mouse neuroblastoma cells—in vitro | SIRT1/Nrf2 pathway | [74] |
| Oltipraz | Rat Schwann cell line | 20 µM on RSC96 cells—Schwann cell line—in vitro | Nrf2/NQO1 pathway | [73] |
| Bardoxolone methyl | Male Sprague Dawley rats | 15 and 30 mg/kg/day, oral | keap1-Nrf2-ARE pathway | [72] |
| Diosgenin | Male C57 mice | 50 and 100 mg/kg, intragastric | Nrf2/HO-1 pathway | [63] |
| Bergenin | Male C57BL/6 mice | 3.125–25 mg/kg, i.p. | Nrf2 pathway | [62] |
| Diphenyl diselenide | Male Sprague Dawley rats | 5 and 15 mg/kg, oral | Nrf2/Keap1 pathway | [70] |
| Deguelin | Male Sprague Dawley rats | 4, 6, and 8 mg/kg, oral gavage | Nrf2 pathway | [64] |
| Tanshinone IIA | Male Sprague Dawley rats | 25 mg/kg, i.p. | Nrf2/ARE pathway | [71] |
| Rutin | Male Wistar rats | Rutin—100 and 200 mg/kg, i.p. Nimesulid—5 and 10 mg/kg, i.p. Combination—200 mg/kg rutin + 10 mg/kg nimesulid, i.p. | Nrf2/HO-1/NF-κB and COX pathway | [79] |
| Sulforaphane | BKS.Cg-m+/+Leprdb/J and db/db mice | Sulforaphane—2.5, 5, and 10 mg/kg, s.c. DPDPE and SNC-80—0.15, 0.5, 1, and 5 mg/ kg, s.c. Combination—10 mg/kg, i.p. sulforaphane alone or 10 mg/kg, i.p. sulforaphane + 0.15 mg/kg, s.c DPDPE, 10 mg/kg, i.p. sulforaphane + 0.5 mg/kg, s.c. SNC-80 | Nrf-2/HO-1 pathway | [81] |
| Quercetin | Sprague Dawley rats’ embryos | 10 mmol/L quercetin, 1 IU/mL hirudin, 100 nmol/L cinnamaldehyde on DRG neurons from 15d embryos of Sprague Dawley rats—in vitro Combination—quercetin + hirudin, quercetin + cinnamaldehyde, cinnamaldehyde + hirudin, quercetin + cinnamaldehyde + hirudin | Nrf-2/HO-1 pathway | [80] |
| Tangluoning | Male Sprague Dawley rats | 10.9 g and 21.8 g crude drug/kg/day, intragastric | PERK/Nrf2 pathway | [55] |
| Isoliquiritigenin | Male Sprague Dawley rats | 10 and 20 mg/kg, oral 2.5 and 5 µM on neuro2a—mouse neuroblastoma cells—in vitro | SIRT1/Nrf2 pathway | [78] |
| Anomalin | Male ICR mice | 50 mg/kg, i.p. 50 µM on DRG primary cells, N2a—mouse neuroblastoma cell line | Nrf2 pathway | [66] |
| Fisetin | Male Sprague Dawley rats | 5 and 10 mg/kg, oral | Nrf2 pathway | [65] |
| Rutin | Male Sprague Dawley rats | 5, 25, and 50 mg/kg, i.p. | Nrf2 pathway | [68] |
| Paeoniflorin | Rat Schwann cell line | 1, 10, and 100 µM in RSC96 cells—Schwann cell line—in vitro | Nrf2/ARE pathway | [61] |
| Taurine | Male Wistar rats | 2% w/v, oral | Nrf2/HO-1 pathway | [69] |
2.2. Chemotherapy-Induced Peripheral Neuropathy (CIPN)
| Nrf2 Inducer | Animals (Sex, Strain) | Dose (mg/kg), Route of Administration | Mechanism of Action | Reference |
|---|---|---|---|---|
| PEA-OXA | Male Wistar rats | 10 mg/kg, oral | NF-κB/Nrf-2 pathway | [97] |
| Oleuropein | Male Wistar rats | Oleuropein—20 mg/kg, oral Combination—Oleuropein—20 mg/kg, oral + suvorexant—an orexin receptor antagonist—20 mg/kg, oral | Nrf2 pathway | [104] |
| Curcumin | Male Sprague Dawley rats | 100 and 200 mg/kg, oral | Nrf2/HO-1 pathway | [98] |
| Mitoquinone | Male ICR mice | 2.5, 5 and 10 mg/kg, intragastric | Nrf2 pathway | [96] |
| Formononetin | Male C57BL/6 mice | 10 mg/kg, i.p. 10 µM on mouse ND7/23 neuron cells, colon cancer cells (CT-26), human colorectal carcinoma cells (Caco-2, DLD-1, and HCT-116), human lung adenocarcinoma cells (PC9, A649, H1975, and HCC8827), human lung squamous cell carcinoma cells (H520), and human pancreatic cancer cells (BxPC3 and Panc1)—in vitro | Keap1-Nrf2-GSTP1 pathway | [101] |
| Resveratrol | Male Sprague Dawley rats | 7 and 14 mg/kg, oral | Nrf2/HO-1 pathway | [100] |
| Quercetin | Male Sprague Dawley rats | 25 and 50 mg/kg, oral | Nrf2/HO-1 pathway | [99] |
| Oltipraz | Male Sprague Dawley rats | 10, 50, 100 mg/kg/day, i.p. | Nrf2/HO-1 pathway | [91] |
| Rosiglitazone | Male Sprague Dawley rats | 5, 25, and 50 mg/kg, i.p. | Nrf2/HO-1 pathway | [92] |
| Levo-corydalmine | Male ICR mice | 5, 10, and 20 mg/kg, intragastric | Nrf2/HO-1/CO pathway | [94] |
| Berberine | Male Wistar rats | 10 and 20 mg/kg, i.p. | Nrf2 pathway | [102] |
| Alphalipoic acid | Male Sprague Dawley rats | 15, 30, and 60 mg/kg, i.p. | Nrf2 pathway | [95] |
| L-carnosine | Male and female Egyptian patients | 500 mg, oral in patients—clinical trial | Nrf2 pathway | [107] |
| Dimethyl fumarate and its metabolite monomethyl fumarate | Rat | 0.3, 1, 3, or 10 mM dimethyl fumarate or monomethyl fumarate on PC12 cell—a rat pheochromocytoma cell | Nrf2 pathway | [103] |
| Sulforaphane | Nrf2+/+ and Nrf2−/− C57BL/6 mice | 5 mg/kg, i.p. 10 µM on DRG neurons | Nrf2 pathway | [89] |
2.3. Other Peripheral Nerve Injury Models
2.3.1. Sciatic Nerve Chronic Constriction Injury (CCI)
| Nrf2 Inducer | Animals (Sex, Strain) | Dose (mg/kg), Route of Administration | Mechanism of Action | Reference |
|---|---|---|---|---|
| Carvacrol | Male Sprague Dawley rats | 30 and 60 mg/kg, oral | Keap1/Nrf-2/p62 pathway | [122] |
| Oleuropein | Male Wistar rats | Oleuropein—10 and 20 mg/kg, oral Combination—Oleuropein—20 mg/kg, oral + suvorexant—an orexin receptor antagonist—10 mg/kg, oral Oleuropein—20 mg/kg, oral + suvorexant—20 mg/kg, oral | Nrf2 pathway | [104] |
| 5-fluoro-2-oxindole | Male C57BL/6J mice | 10 mg/kg, i.p. | Nrf2/HO-1/NQO1 pathway | [133] |
| Dexmedetomidine | Male Sprague Dawley rats | 15 g/kg at 5 g/kg/h, i.p. | Keap1–Nrf2–HO-1 pathway | [128] |
| Dexmedetomidine | Male Sprague Dawley rats | 1, 2, and 5 µg/kg, i.p. | NLRP3/Nrf2 pathway | [129] |
| RTA-408 | Male C57BL/6J mice | 1, 5, and 10 μg, i.t. | Nrf2 pathway | [120] |
| Bromelain | Male Wistar rats | 30 and 50 mg/kg, oral | Nrf2 pathway | [114] |
| Paeoniflorin | Male Sprague Dawley rats | 25, 50, and 100 mg/kg, i.p. | Keap1-Nrf2 pathway | [115] |
| Thymus algeriensis and T. fontanesii extracts | Male Wistar rats | 200 and 400 mg/kg, oral | Nrf2 pathway | [116] |
| Diallyl disulfide and diallyl trisulfide | Male Wistar albino rats | Diallyl disulfide—25 and 50 mg/kg, oral Diallyl trisulfide—20 and 40 mg/kg, oral | H2S-BDNF-Nrf2 pathway | [112] |
| NaHS (a common donor for H2S) | Male Sprague Dawley rats | 15, 30, 60 mg/kg, abdominal cavity administration | Nrf2/HO-1 pathway | [109] |
| Oltipraz | Male C57BL/6J mice | 10 mg/kg, i.p. | Nrf2/HO-1/NQO1 pathway | [124] |
| UFP-512 | Male C57BL/6J mice | UFP-512—1, 3, 10, 20, and 30 mg/kg i.p. Combination—1 mg/kg, i.p. UFP-512 + sulforaphane—10 mg/kg, i.p. | Nrf2/HO-1 pathway | [119] |
| Sulforaphane | Male C57BL/6J mice | 10 mg/kg, i.p. | Nrf2/HO-1/NQO1 pathway | [132] |
| Plumbagin | Male Sprague Dawley rats | 10 and 20 mg/kg, oral | Nrf2 pathway | [118] |
| Quercetin + PARP inhibitor-4-ANI | Male Sprague Dawley rats | Quercetin—25 mg/kg, oral 4-ANI—3 mg/kg, oral Combination—quercetin + 4-ANI | Nrf2 pathway | [117] |
| Sulforaphane | Male C57BL/6J mice | 0.1–100 mg/kg, i.p. | Nrf2 pathway | [134] |
2.3.2. Sciatic Nerve Crush (SNC) Injury
| Nrf2 Inducer | Animals (Sex, Strain) | Dose (mg/kg), Route of Administration | Mechanism of Action | Reference |
|---|---|---|---|---|
| Sciatic nerve crush (SNC) injury | ||||
| Isoquercitrin | Male ICR mice | 20 mg/kg/day, i.p. 40 to 320 μM isoquercitrin on primary Schwann cells isolated from sciatic nerves of neonatal 1-day-old Sprague Dawley rats—in vitro | Nrf2 pathway | [139] |
| Curcumin | Male Sprague Dawley rats | 0.2 mg/day, continuous delivery through mini-osmotic pumps | Nrf2 pathway | [140] |
| Sesame oil | Male SPF C57BL/6 mice | 0.5, 1 and 2 mL/kg, oral | Nrf2 pathway | [141] |
| Partial sciatic nerve ligation (PSNL) | ||||
| ECN | Male albino mice | 1 and 5 mg/kg, i.p. | Nrf2/HO-1/NQO1 pathway | [142] |
| Spared nerve injury (SNI) | ||||
| tBHQ | Male Sprague Dawley rats | 1 and 10 μM, i.t. | Nrf2 pathway | [143] |
| Dimethyl fumarate | Male Sprague Dawley rats and male and female wild type and Nfe2l2−/− mice | 30, 100, and 300 5 mL−1 kg−1, oral | Nrf2 pathway | [144] |
| Sulforaphane | Male Sprague Dawley rats | 30 mg/kg, i.p. | Keap1-Nrf2 signaling | [145] |
| Spinal nerve ligation (SNL) | ||||
| Dimethylitaconate | Male C57BL/6 mice | 10 mg or 20 mg, i.p. 250 µM on BV2 microglial cell line | Nrf2 pathway | [146] |
2.3.3. Partial Sciatic Nerve Ligation (PSNL)
2.3.4. Spared Nerve Injury (SNI)
2.3.5. Spinal Nerve Ligation (SNL)
3. Conclusions and Future Directions
Funding
Conflicts of Interest
Abbreviations
| 4-ANI | 4-amino 1, 8-naphthalimide |
| 8-iso | PGF2α 8-isoprostaglandin F2α |
| 8-OHdG | 8-hydroxy-2′-deoxyguanosine |
| AREs | antioxidant response elements |
| BDNF | brain-derived neurotrophin factor |
| BRD4 | bromodomain-containing protein 4 |
| BTB | bric-a-brac, tramtrack, broad-complex |
| bZip | basic region leucine zipper |
| CCI | chronic constriction injury |
| CIPN | chemotherapy-induced peripheral neuropathy |
| CoPP | cobalt protoporphyrin IX |
| COX-2 | cyclooxygenase-2 |
| Cul | cullin |
| DN | diabetic neuropathy |
| DOR | delta opioid receptor |
| DPDPE | [dPen(2),d-Pen(5)]-Enkephalin] |
| DRG | dorsal root ganglion |
| ECN | 7β-(3-Ethyl-cis-crotonoyloxy)-1α-(2-methylbutyryloxy)-3,14-dehydro-Z-notonipetranone |
| ERK | extracellular signal-regulated kinase |
| GSH | glutathione |
| GSTP1 | glutathione S-transferase pi 1 |
| H2S | hydrogen sulfide |
| HCAR2 | hydroxycarboxylic acid receptor 2 |
| HO-1 | heme oxygenase-1 |
| i.p. | intraperitoneal |
| IL-1β | interleukin-1 beta |
| IL-6 | interleukin-6 |
| JNK | c-Jun N-terminal kinase |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MAF | muscle aponeurosis fibromatosis |
| MCQ | mitochondrial quality control |
| mPFC | medial prefrontal cortex |
| NAD | nicotinamide adenosine dinucleotide |
| NF-κB | nuclear factor kappa B |
| NLRP3 | NOD-like receptor protein 3 |
| NOX4 | nicotinamide adenine dinucleotide phosphate, reduced (NADPH) oxidase 4 |
| NQO1 | NAD(P)H:quinone oxidoreductase1 |
| Nrf2 | nuclear factor erythroid 2 (NFE2)-related factor 2 (Nrf2) |
| PARP | poly (ADP-ribose) polymerase |
| PEA | palmitoylethanolamide |
| PEA-OXA | 2-pentadecyl-2-oxazoline of palmitoylethanolamide |
| PERK | protein kinase RNA-like endoplasmic reticulum kinase |
| PGC-1α | peroxisome proliferator activated receptor-gamma coactivator-1α |
| PN | peripheral neuropathy |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| PSNL | partial sciatic nerve ligation |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| s.c. | subcutaneous |
| SC | Schwann cell |
| SIRT | sirtuin |
| SNC | sciatic nerve crush |
| SNC-80 | (+)-4-[(α(R)-α-[(2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl]-(3-methoxybenzyl)-N,N-diethylbenzamide] |
| SNI | spared nerve injury |
| SNL | spinal nerve ligation |
| SOD | superoxide dismutase |
| STZ | streptozotocin |
| TNF-α | tumor necrosis factor alpha |
| TRP | transient receptor potential |
| WT | wildtype |
References
- Treede, R.-D.; Jensen, T.S.; Campbell, J.N.; Cruccu, G.; Dostrovsky, J.O.; Griffin, J.W.; Hansson, P.; Hughes, R.; Nurmikko, T.; Serra, J. Neuropathic pain. Redefinition and a grading system for clinical and research purposes. Neurology 2008, 70, 1630–1635. [Google Scholar] [CrossRef]
- Jensen, T.S.; Baron, R.; Haanpää, M.; Kalso, E.; Loeser, J.D.; Rice, A.S.C.; Treede, R.-D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef] [PubMed]
- van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.H.; Torrance, N. Epidemiology of Neuropathic Pain and Its Impact on Quality of Life. Curr. Pain Headache Rep. 2012, 16, 191–198. [Google Scholar] [CrossRef]
- Meacham, K.; Shepherd, A.; Mohapatra, D.P.; Haroutounian, S. Neuropathic Pain: Central vs. Peripheral Mechanisms. Curr. Pain Headache Rep. 2017, 21, 28. [Google Scholar] [CrossRef]
- Martyn, C.N.; Hughes, R.A. Epidemiology of peripheral neuropathy. J. Neurol. Neurosurg. Psychiatry 1997, 62, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Betteridge, D.; Betteridge, D.J. What is oxidative stress? Metab. Clin. Exp. 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Paiva, C.; Bozza, M. Are Reactive Oxygen Species Always Detrimental to Pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [Green Version]
- Flatters, S.J.; Bennett, G.J. Studies of peripheral sensory nerves in paclitaxel-induced painful peripheral neuropathy: Evidence for mitochondrial dysfunction. Pain 2006, 122, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Zheng, H.; Bennett, G. Characterization of oxaliplatin-induced chronic painful peripheral neuropathy in the rat and comparison with the neuropathy induced by paclitaxel. Neuroscience 2012, 203, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.H.; Zheng, H.; Zheng, F.Y.; Nuydens, R.; Meert, T.F.; Bennett, G.J. Mitochondrial abnormality in sensory, but not motor, axons in paclitaxel-evoked painful peripheral neuropathy in the rat. Neuroscience 2011, 199, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Du, X.L.; Edelstein, D.; Dimmeler, S.; Ju, Q.; Sui, C.; Brownlee, M. Hyperglycemia inhibits endothelial nitric oxide synthase activity by posttranslational modification at the Akt site. J. Clin. Investig. 2001, 108, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, S.S.; Skulachev, V.P.; Starkov, A.A. High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 1997, 416, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen-Heininger, Y.; Mossman, B.; Heintz, N.; Forman, H.; Kalyanaraman, B.; Finkel, T.; Stamler, J.; Rhee, S.G.; van der Vliet, A. Redox-based regulation of signal transduction: Principles, pitfalls, and promises. Free Radic. Biol. Med. 2008, 45, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell Biol. 2003, 15, 247–254. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Fattori, V.; Manchope, M.F.; Mizokami, S.S.; Casagrande, R.; Verri, W.A. Naringenin reduces inflammatory pain in mice. Neuropharmacology 2016, 105, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Verri, W.A., Jr.; Vicentini, F.T.; Baracat, M.M.; Georgetti, S.R.; Cardoso, R.D.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Fonseca, M.J.; Casagrande, R. Flavonoids as anti-inflammatory and analgesic drugs: Mechanisms of action and perspectives in the development of pharmaceutical forms. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2012; Volume 36, pp. 297–330. [Google Scholar]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Hennig, P.; Garstkiewicz, M.; Grossi, S.; Di Filippo, M.; French, L.E.; Beer, H.-D. The crosstalk between Nrf2 and inflammasomes. Int. J. Mol. Sci. 2018, 19, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Sherratt, P.J.; Pickett, C.B. Regulatory mechanisms controlling gene expression mediated by the antioxidant response element. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 233–260. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Tong, K.I.; Mio, K.; Maruyama, Y.; Kurokawa, H.; Sato, C.; Yamamoto, M. Keap1 is a forked-stem dimer structure with two large spheres enclosing the intervening, double glycine repeat, and C-terminal domains. Proc. Natl. Acad. Sci. USA 2010, 107, 2842–2847. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, D.; Hannink, M.; Beamer, L.J. Crystal structure of the Kelch domain of human Keap1. J. Biol. Chem. 2004, 279, 54750–54758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, B.; Tong, K.I.; Ohta, T.; Nakamura, Y.; Scharlock, M.; Ohtsuji, M.; Kang, M.-I.; Kobayashi, A.; Yokoyama, S.; Yamamoto, M. Structural basis for defects of Keap1 activity provoked by its point mutations in lung cancer. Mol. Cell 2006, 21, 689–700. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chen, M.G.; Lin, G.X.; Ma, Q. Arsenic induces NAD (P) H-quinone oxidoreductase I by disrupting the Nrf2· Keap1· Cul3 complex and recruiting Nrf2· Maf to the antioxidant response element enhancer. J. Biol. Chem. 2006, 281, 23620–23631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, A.; Kang, M.-I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.D.; Lo, S.-C.; Cross, J.V.; Templeton, D.J.; Hannink, M. Keap1 is a redox-regulated substrate adaptor protein for a Cul3-dependent ubiquitin ligase complex. Mol. Cell. Biol. 2004, 24, 10941–10953. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, A.R.; Dos Santos, N.B.; Scavone, C.; Munhoz, C.D. Nrf2/ARE pathway modulation by dietary energy regulation in neurological disorders. Front. Pharmacol. 2019, 10, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, S.; Pergola, P.E.; Zager, R.A.; Vaziri, N.D. Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 2013, 83, 1029–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Yamamoto, M. Nrf2–Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Ma, Q.; He, X. Molecular basis of electrophilic and oxidative defense: Promises and perils of Nrf2. Pharmacol. Rev. 2012, 64, 1055–1081. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.; Han, X.-D.; Kan, Y.W. An important function of Nrf2 in combating oxidative stress: Detoxification of acetaminophen. Proc. Natl. Acad. Sci. USA 2001, 98, 4611–4616. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, C.D.; Reisman, S.A. Nrf2 the rescue: Effects of the antioxidative/electrophilic response on the liver. Toxicol. Appl. Pharmacol. 2010, 244, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.M.; Cho, H.-Y.; Kleeberger, S.R. Oxidative stress and antioxidants in the pathogenesis of pulmonary fibrosis: A potential role for Nrf2. Antioxid. Redox Signal. 2008, 10, 321–332. [Google Scholar] [CrossRef]
- de la Vega, M.R.; Dodson, M.; Gross, C.; Mansour, H.M.; Lantz, R.C.; Chapman, E.; Wang, T.; Black, S.M.; Garcia, J.G.; Zhang, D.D. Role of Nrf2 and autophagy in acute lung injury. Curr. Pharmacol. Rep. 2016, 2, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Pajares, M.; Jiménez-Moreno, N.; Dias, I.H.; Debelec, B.; Vucetic, M.; Fladmark, K.E.; Basaga, H.; Ribaric, S.; Milisav, I.; Cuadrado, A. Redox control of protein degradation. Redox Biol. 2015, 6, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-Q.; Zhang, B.; He, X.-M.; Li, D.-D.; Shi, J.-S.; Zhang, F. Naringenin targets on astroglial Nrf2 to support dopaminergic neurons. Pharmacol. Res. 2019, 139, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Wati, S.M.; Matsumaru, D.; Motohashi, H. NRF2 pathway activation by KEAP1 inhibition attenuates the manifestation of aging phenotypes in salivary glands. Redox Biol. 2020, 36, 101603. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Jiménez-Moreno, N.; García-Yagüe, Á.J.; Escoll, M.; de Ceballos, M.L.; Van Leuven, F.; Rábano, A.; Yamamoto, M.; Rojo, A.I.; Cuadrado, A. Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 2016, 12, 1902–1916. [Google Scholar] [CrossRef] [Green Version]
- Pajares, M.; Cuadrado, A.; Rojo, A.I. Modulation of proteostasis by transcription factor NRF2 and impact in neurodegenerative diseases. Redox Biol. 2017, 11, 543–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Vinik, A.I.; Maser, R.E.; Mitchell, B.D.; Freeman, R. Diabetic Autonomic Neuropathy. Diabetes Care 2003, 26, 1553–1579. [Google Scholar] [CrossRef] [Green Version]
- Ganesh Yerra, V.; Negi, G.; Sharma, S.S.; Kumar, A. Potential therapeutic effects of the simultaneous targeting of the Nrf2 and NF-κB pathways in diabetic neuropathy. Redox Biol. 2013, 1, 394–397. [Google Scholar] [CrossRef] [Green Version]
- WADA, R.; Yagihashi, S. Role of Advanced Glycation End Products and Their Receptors in Development of Diabetic Neuropathy. Ann. N. Y. Acad. Sci. 2005, 1043, 598–604. [Google Scholar] [CrossRef]
- Das Evcimen, N.; King, G.L. The role of protein kinase C activation and the vascular complications of diabetes. Pharmacol. Res. 2007, 55, 498–510. [Google Scholar] [CrossRef]
- Banning, A.; Brigelius-Flohé, R. NF-κB, Nrf2, and HO-1 Interplay in Redox-Regulated VCAM-1 Expression. Antioxid. Redox Signal. 2005, 7, 889–899. [Google Scholar] [CrossRef]
- Vincent, A.; Edwards, J.; Sadidi, M.; Feldman, E. The Antioxidant Response as a Drug Target in Diabetic Neuropathy. Curr. Drug Targets 2008, 9, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Negi, G.; Kumar, A.; Sharma, S.S. Melatonin modulates neuroinflammation and oxidative stress in experimental diabetic neuropathy: Effects on NF-κB and Nrf2 cascades. J. Pineal Res. 2011, 50, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.-S.; Yu, S.; Kong, A.-N. Activation of Nrf2-antioxidant signaling attenuates NFκB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.-H.; Qu, J.; Shen, X. NF-κB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 713–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Li, H.; Liu, Q.; Liu, F.; Tang, L.; Li, C.; Yuan, Y.; Zhan, Y.; Xu, W.; Li, W.; et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell. Signal. 2011, 23, 883–892. [Google Scholar] [CrossRef]
- Soares, M.P.; Seldon, M.P.; Gregoire, I.P.; Vassilevskaia, T.; Berberat, P.O.; Yu, J.; Tsui, T.-Y.; Bach, F.H. Heme Oxygenase-1 Modulates the Expression of Adhesion Molecules Associated with Endothelial Cell Activation. J. Immunol. 2004, 172, 3553–3563. [Google Scholar] [CrossRef] [Green Version]
- Low, P.A.; Lagerlund, T.D.; McManis, P.G. Nerve blood flow and oxygen delivery in normal, diabetic, and ischemic neuropathy. Int. Rev. Neurobiol. 1989, 31, 355–438. [Google Scholar]
- Tang, W.; Chen, X.; Liu, H.; Lv, Q.; Zou, J.; Shi, Y.; Liu, Z. Expression of Nrf2 promotes schwann cell-mediated sciatic nerve recovery in diabetic peripheral neuropathy. Cell. Physiol. Biochem. 2018, 46, 1879–1894. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yao, W.; Liu, H.; Gao, Y.; Liu, R.; Xu, L. Tangluoning, a traditional Chinese medicine, attenuates in vivo and in vitro diabetic peripheral neuropathy through modulation of PERK/Nrf2 pathway. Sci. Rep. 2017, 7, 1014. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yao, W.; Shi, H.; Liu, H.; Li, Y.; Gao, Y.; Liu, R.; Xu, L. Paeoniflorin protects Schwann cells against high glucose induced oxidative injury by activating Nrf2/ARE pathway and inhibiting apoptosis. J. Ethnopharmacol. 2016, 185, 361–369. [Google Scholar] [CrossRef]
- Villarreal, C.F.; Santos, D.S.; Lauria, P.S.; Gama, K.B.; Espírito-Santo, R.F.; Juiz, P.J.; Alves, C.Q.; David, J.M.; Soares, M.B. Bergenin reduces experimental painful diabetic neuropathy by restoring redox and immune homeostasis in the nervous system. Int. J. Mol. Sci. 2020, 21, 4850. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Li, X.; Tian, H.; Liu, C.; Guo, Y.; Zhang, S.; Chu, Y.; Li, J.; Wang, Y.; Zhang, L. Neuroprotective effect of diosgenin in a mouse model of diabetic peripheral neuropathy involves the Nrf2/HO-1 pathway. BMC Complement. Med. Ther. 2020, 20, 126. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, W.; Yi, H.; Hu, X.; Peng, L.; Yang, F. The natural rotenoid deguelin ameliorates diabetic neuropathy by decreasing oxidative stress and plasma glucose levels in rats via the Nrf2 signalling pathway. Cell. Physiol. Biochem. 2018, 48, 1164–1176. [Google Scholar] [CrossRef]
- Sandireddy, R.; Yerra, V.G.; Komirishetti, P.; Areti, A.; Kumar, A. Fisetin imparts neuroprotection in experimental diabetic neuropathy by modulating Nrf2 and NF-κB pathways. Cell. Mol. Neurobiol. 2016, 36, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Choi, R.J.; Lee, J.; Kim, Y.S. Attenuation of neuropathic pain and neuroinflammatory responses by a pyranocoumarin derivative, anomalin in animal and cellular models. Eur. J. Pharmacol. 2016, 774, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yu, H.; Lin, Q.; Liu, X.; Cheng, Y.; Deng, B. Anti-inflammatory effect of resveratrol attenuates the severity of diabetic neuropathy by activating the Nrf2 pathway. Aging 2021, 13, 10659. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Yang, W.; Xue, Q.; Gao, L.; Huo, J.; Ren, D.; Chen, X. Rutin ameliorates diabetic neuropathy by lowering plasma glucose and decreasing oxidative stress via Nrf2 signaling pathway in rats. Eur. J. Pharmacol. 2016, 771, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Agca, C.A.; Tuzcu, M.; Hayirli, A.; Sahin, K. Taurine ameliorates neuropathy via regulating NF-κB and Nrf2/HO-1 signaling cascades in diabetic rats. Food Chem. Toxicol. 2014, 71, 116–121. [Google Scholar] [CrossRef]
- Wang, X.; Huan, Y.; Li, C.; Cao, H.; Sun, S.; Lei, L.; Liu, Q.; Liu, S.; Ji, W.; Liu, H. Diphenyl diselenide alleviates diabetic peripheral neuropathy in rats with streptozotocin-induced diabetes by modulating oxidative stress. Biochem. Pharmacol. 2020, 182, 114221. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.-B.; Qiu, H.-Y. Neuroprotective effect of tanshinone IIA against neuropathic pain in diabetic rats through the Nrf2/ARE and NF-κB signaling pathways. Kaohsiung J. Med. Sci. 2018, 34, 428–437. [Google Scholar] [CrossRef]
- Kalvala, A.K.; Kumar, R.; Sherkhane, B.; Gundu, C.; Arruri, V.K.; Kumar, A. Bardoxolone methyl ameliorates hyperglycemia induced mitochondrial dysfunction by activating the keap1-Nrf2-ARE pathway in experimental diabetic neuropathy. Mol. Neurobiol. 2020, 57, 3616–3631. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Bian, M.; Wu, J.; Li, D.; Ding, L.; Zeng, Q. Oltipraz Prevents High Glucose-Induced Oxidative Stress and Apoptosis in RSC96 Cells through the Nrf2/NQO1 Signalling Pathway. BioMed Res. Int. 2020, 2020, 5939815. [Google Scholar] [CrossRef]
- Bheereddy, P.; Yerra, V.G.; Kalvala, A.K.; Sherkhane, B.; Kumar, A. SIRT1 Activation by Polydatin Alleviates Oxidative Damage and Elevates Mitochondrial Biogenesis in Experimental Diabetic Neuropathy. Cell. Mol. Neurobiol. 2020, 41, 1563–1577. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Marshall, B.L.; Borra, M.T.; Denu, J.M.; Verdin, E. The human Sir2 ortholog, SIRT2, is an NAD+-dependent tubulin deacetylase. Mol. Cell 2003, 11, 437–444. [Google Scholar] [CrossRef]
- Huang, K.; Gao, X.; Wei, W. The crosstalk between Sirt1 and Keap1/Nrf2/ARE anti-oxidative pathway forms a positive feedback loop to inhibit FN and TGF-β1 expressions in rat glomerular mesangial cells. Exp. Cell Res. 2017, 361, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Li, J.; Chen, Z.; Huang, J.; Chen, Q.; Cai, W.; Liu, P.; Huang, H. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating CKIP-1 to resist HG-induced up-regulation of FN and ICAM-1 in GMCs and diabetic mice kidneys. Free Radic. Biol. Med. 2017, 106, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Yerra, V.G.; Kalvala, A.K.; Kumar, A. Isoliquiritigenin reduces oxidative damage and alleviates mitochondrial impairment by SIRT1 activation in experimental diabetic neuropathy. J. Nutr. Biochem. 2017, 47, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Kumar, A.; Singh, D.P.; Bishnoi, M.; Nag, T.C. Ameliorative potential of rutin in combination with nimesulide in STZ model of diabetic neuropathy: Targeting Nrf2/HO-1/NF-kB and COX signalling pathway. Inflammopharmacology 2018, 26, 755–768. [Google Scholar] [CrossRef]
- Shi, Y.; Liang, X.-c.; Zhang, H.; Sun, Q.; Wu, Q.-l.; Qu, L. Combination of quercetin, cinnamaldehyde and hirudin protects rat dorsal root ganglion neurons against high glucose-induced injury through Nrf-2/HO-1 activation and NF-κB inhibition. Chin. J. Integr. Med. 2017, 23, 663–671. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, C.; Leánez, S.; Pol, O. The induction of the transcription factor Nrf2 enhances the antinociceptive effects of delta-opioid receptors in diabetic mice. PLoS ONE 2017, 12, e0180998. [Google Scholar] [CrossRef] [PubMed]
- Hershman, D.L.; Lacchetti, C.; Dworkin, R.H.; Lavoie Smith, E.M.; Bleeker, J.; Cavaletti, G.; Chauhan, C.; Gavin, P.; Lavino, A.; Lustberg, M.B. Prevention and management of chemotherapy-induced peripheral neuropathy in survivors of adult cancers: American Society of Clinical Oncology clinical practice guideline. J. Clin. Oncol. 2014, 32, 1941–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banach, M.; Juranek, J.K.; Zygulska, A.L. Chemotherapy-induced neuropathies—A growing problem for patients and health care providers. Brain Behav. 2017, 7, e00558. [Google Scholar] [CrossRef] [PubMed]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of chemotherapy-induced peripheral neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, T.J.; Sedhom, R.; Gupta, A. Chemotherapy-induced peripheral neuropathy. JAMA Oncol. 2019, 5, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carozzi, V.; Canta, A.; Chiorazzi, A. Chemotherapy-induced peripheral neuropathy: What do we know about mechanisms? Neurosci. Lett. 2015, 596, 90–107. [Google Scholar] [CrossRef] [PubMed]
- Gamelin, E.; Gamelin, L.; Bossi, L.; Quasthoff, S. Clinical aspects and molecular basis of oxaliplatin neurotoxicity: Current management and development of preventive measures. Proc. Semin. Oncol. 2002, 29, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Tofthagen, C.; Donovan, K.A.; Morgan, M.A.; Shibata, D.; Yeh, Y. Oxaliplatin-induced peripheral neuropathy’s effects on health-related quality of life of colorectal cancer survivors. Support. Care Cancer 2013, 21, 3307–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Luo, L.; Cai, X.; Fang, Y.; Wang, J.; Chen, G.; Yang, J.; Zhou, Q.; Sun, X.; Cheng, X. Nrf2 inhibits oxaliplatin-induced peripheral neuropathy via protection of mitochondrial function. Free Radic. Biol. Med. 2018, 120, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Xu, J.; Xu, D.; Ma, X.; Zhao, X.; Liu, L. Nociceptive behavior induced by chemotherapeutic paclitaxel and beneficial role of antioxidative pathways. Physiol. Res. 2019, 68, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-Q.; Liu, D.-Q.; Chen, S.-P.; Chen, N.; Sun, J.; Wang, X.-M.; Cao, F.; Tian, Y.-K.; Ye, D.-W. Nrf2 activation ameliorates mechanical allodynia in paclitaxel-induced neuropathic pain. Acta Pharmacol. Sin. 2020, 41, 1041–1048. [Google Scholar] [CrossRef]
- Zhou, Y.-Q.; Liu, D.-Q.; Chen, S.-P.; Chen, N.; Sun, J.; Wang, X.-M.; Li, D.-Y.; Tian, Y.-K.; Ye, D.-W. PPARγ activation mitigates mechanical allodynia in paclitaxel-induced neuropathic pain via induction of Nrf2/HO-1 signaling pathway. Biomed. Pharmacother. 2020, 129, 110356. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Wang, R.; Cui, G.; Li, X.; Wang, T.; Li, X. Engagement of microRNA-155 in exaggerated oxidative stress signal and TRPA1 in the dorsal horn of the spinal cord and neuropathic pain during chemotherapeutic oxaliplatin. Neurotox. Res. 2019, 36, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ao, L.; Yan, Y.; Li, C.; Li, W.; Ye, A.; Liu, J.; Hu, Y.; Fang, W.; Li, Y. Levo-corydalmine attenuates vincristine-induced neuropathic pain in mice by upregulating the Nrf2/HO-1/CO pathway to inhibit connexin 43 expression. Neurotherapeutics 2020, 17, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, X.; Wang, Z.; Wang, P.; Zhang, Z.; Dong, J.; Zhuang, R.; Zhou, Y.; Ma, G.; Cai, W. Alphalipoic acid prevents oxidative stress and peripheral neuropathy in nab-paclitaxel-treated rats through the Nrf2 signalling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 3142732. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-J.; Wang, L.; Song, X.-Y. Mitoquinone alleviates vincristine-induced neuropathic pain through inhibiting oxidative stress and apoptosis via the improvement of mitochondrial dysfunction. Biomed. Pharmacother. 2020, 125, 110003. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Lanza, M.; Paterniti, I.; Filippone, A.; Ardizzone, A.; Casili, G.; Scuderi, S.A.; Puglisi, C.; Mare, M.; Memeo, L. PEA-OXA Mitigates Oxaliplatin-Induced Painful Neuropathy through NF-κB/Nrf-2 Axis. Int. J. Mol. Sci. 2021, 22, 3927. [Google Scholar] [CrossRef] [PubMed]
- Yardım, A.; Kandemir, F.M.; Çomaklı, S.; Özdemir, S.; Caglayan, C.; Kucukler, S.; Çelik, H. Protective effects of curcumin against paclitaxel-induced spinal cord and sciatic nerve injuries in rats. Neurochem. Res. 2021, 46, 379–395. [Google Scholar] [CrossRef]
- Yardim, A.; Kandemir, F.M.; Ozdemir, S.; Kucukler, S.; Comakli, S.; Gur, C.; Celik, H. Quercetin provides protection against the peripheral nerve damage caused by vincristine in rats by suppressing caspase 3, NF-κB, ATF-6 pathways and activating Nrf2, Akt pathways. Neurotoxicology 2020, 81, 137–146. [Google Scholar] [CrossRef]
- Recalde, M.; Miguel, C.A.; Noya-Riobó, M.V.; Gonzalez, S.L.; Villar, M.J.; Coronel, M.F. Resveratrol exerts anti-oxidant and anti-inflammatory actions and prevents oxaliplatin-induced mechanical and thermal allodynia. Brain Res. 2020, 1748, 147079. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Ye, J.; Zhao, B.; Sun, J.; Gu, N.; Chen, X.; Ren, L.; Chen, J.; Cai, X.; Zhang, W. Formononetin ameliorates oxaliplatin-induced peripheral neuropathy via the KEAP1-NRF2-GSTP1 axis. Redox Biol. 2020, 36, 101677. [Google Scholar] [CrossRef]
- Singh, J.; Saha, L.; Singh, N.; Kumari, P.; Bhatia, A.; Chakrabarti, A. Study of nuclear factor-2 erythroid related factor-2 activator, berberine, in paclitaxel induced peripheral neuropathy pain model in rats. J. Pharm. Pharmacol. 2019, 71, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Kawashiri, T.; Miyagi, A.; Shimizu, S.; Shigematsu, N.; Kobayashi, D.; Shimazoe, T. Dimethyl fumarate ameliorates chemotherapy agent-induced neurotoxicity in vitro. J. Pharmacol. Sci. 2018, 137, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ma, D.; Zhang, H.; Tang, Y.; Wang, J.; Li, R.; Wen, W.; Zhang, Y. Antinociceptive effects of oleuropein in experimental models of neuropathic pain in male rats. Korean J. Pain 2021, 34, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, Y. Suppressing BRD4 exhibits protective effects against vincristine-induced peripheral neuropathy by alleviating inflammation and oxidative stress. Biochem. Biophys. Res. Commun. 2020, 532, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, L.; Wang, Y.; Wang, G.; Zhao, Y.; Zhang, Y. Electroacupuncture enhances antioxidative signal pathway and attenuates neuropathic pain induced by chemotherapeutic paclitaxel. Physiol. Res. 2019, 68, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.; Saleh, S.; El Abhar, H.; Saad, A.S.; Schaalan, M. L-Carnosine protects against Oxaliplatin-induced peripheral neuropathy in colorectal cancer patients: A perspective on targeting Nrf-2 and NF-κB pathways. Toxicol. Appl. Pharmacol. 2019, 365, 41–50. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.-K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Chen, H.; Xie, K.; Chen, Y.; Wang, Y.; Wang, Y.; Lian, N.; Zhang, K.; Yu, Y. Nrf2/HO-1 signaling pathway participated in the protection of hydrogen sulfide on neuropathic pain in rats. Int. Immunopharmacol. 2019, 75, 105746. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kida, K.; Marutani, E.; Nguyen, R.K.; Ichinose, F. Inhaled hydrogen sulfide prevents neuropathic pain after peripheral nerve injury in mice. Nitric Oxide 2015, 46, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yang, Y.; Wang, C.; Huang, J.; Wang, X.; Liu, Y.; Wang, H. Exploring the role and mechanisms of diallyl trisulfide and diallyl disulfide in chronic constriction-induced neuropathic pain in rats. Korean J. Pain 2020, 33, 216. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Chaiwut, P.; Rawdkuen, S. Extraction of bromelain from pineapple peels. Food Sci. Technol. Int. 2011, 17, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Bakare, A.O.; Owoyele, B.V. Antinociceptive and neuroprotective effects of bromelain in chronic constriction injury-induced neuropathic pain in Wistar rats. Korean J. Pain 2020, 33, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Cheng, J.; Ma, S.; Zhou, J. Paeoniflorin attenuates chronic constriction injury-induced neuropathic pain by suppressing spinal NLRP3 inflammasome activation. Inflammopharmacology 2020, 28, 1495–1508. [Google Scholar] [CrossRef]
- Rezq, S.; Alsemeh, A.E.; D’Elia, L.; El-Shazly, A.M.; Monti, D.M.; Sobeh, M.; Mahmoud, M.F. Thymus algeriensis and Thymus fontanesii exert neuroprotective effect against chronic constriction injury-induced neuropathic pain in rats. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Komirishetty, P.; Areti, A.; Gogoi, R.; Sistla, R.; Kumar, A. Combination strategy of PARP inhibitor with antioxidant prevent bioenergetic deficits and inflammatory changes in CCI-induced neuropathy. Neuropharmacology 2017, 113, 137–147. [Google Scholar] [CrossRef]
- Arruri, V.; Komirishetty, P.; Areti, A.; Dungavath, S.K.N.; Kumar, A. Nrf2 and NF-κB modulation by Plumbagin attenuates functional, behavioural and biochemical deficits in rat model of neuropathic pain. Pharmacol. Rep. 2017, 69, 625–632. [Google Scholar] [CrossRef]
- Polo, S.; Díaz, A.F.; Gallardo, N.; Leánez, S.; Balboni, G.; Pol, O. Treatment with the delta opioid agonist UFP-512 alleviates chronic inflammatory and neuropathic pain: Mechanisms implicated. Front. Pharmacol. 2019, 10, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Li, J.-Y.; Zhang, L.-Q.; Li, D.-Y.; Wu, J.-Y.; Gao, S.-J.; Zhou, Y.-Q.; Mei, W. Nrf2 Activation Attenuates Chronic Constriction Injury-induced Neuropathic Pain via Induction of PGC-1α-mediated Mitochondrial Biogenesis in the Spinal Cord. Oxid. Med. Cell. Longev. 2021, 2021, 9577874. [Google Scholar] [CrossRef] [PubMed]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The bioactivity and toxicological actions of carvacrol. Crit. Rev. Food Sci. Nutr. 2015, 55, 304–318. [Google Scholar] [CrossRef]
- Arruri, V.K.; Gundu, C.; Kalvala, A.K.; Sherkhane, B.; Khatri, D.K.; Singh, S.B. Carvacrol abates NLRP3 inflammasome activation by augmenting Keap1/Nrf-2/p62 directed autophagy and mitochondrial quality control in neuropathic pain. Nutr. Neurosci. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Mankowski, R.T.; Burman, J.L.; Donisi, L.; Kim, J.-S.; Marzetti, E.; Leeuwenburgh, C. Mitochondrial quality control mechanisms as molecular targets in cardiac ageing. Nat. Rev. Cardiol. 2018, 15, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.F.; Polo, S.; Gallardo, N.; Leánez, S.; Pol, O. Analgesic and antidepressant effects of oltipraz on neuropathic pain in mice by modulating microglial activation. J. Clin. Med. 2019, 8, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Hernández, D.; Bris, Á.G.; MacDowell, K.S.; García-Bueno, B.; Madrigal, J.L.; Leza, J.C.; Caso, J.R. Modulation of the antioxidant nuclear factor (erythroid 2-derived)-like 2 pathway by antidepressants in rats. Neuropharmacology 2016, 103, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Cortinez, L.I.; Hsu, Y.-W.; Sum-Ping, S.T.; Young, C.; Keifer, J.C.; MacLeod, D.; Robertson, K.M.; Wright, D.R.; Moretti, E.W.; Somma, J. Dexmedetomidine pharmacodynamics: Part II: Crossover comparison of the analgesic effect of dexmedetomidine and remifentanil in healthy volunteers. J. Am. Soc. Anesthesiol. 2004, 101, 1077–1083. [Google Scholar] [CrossRef]
- Hsu, Y.-W.; Cortinez, L.I.; Robertson, K.M.; Keifer, J.C.; Sum-Ping, S.T.; Moretti, E.W.; Young, C.C.; Wright, D.R.; MacLeod, D.B.; Somma, J. Dexmedetomidine pharmacodynamics: Part I: Crossover comparison of the respiratory effects of dexmedetomidine and remifentanil in healthy volunteers. J. Am. Soc. Anesthesiol. 2004, 101, 1066–1076. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, W.; Wang, X.-Q.; Wan, Z.-H.; Liu, Y.-Q.; Zhang, M.-J. Dexmedetomidine Relieves Neuropathic Pain in Rats With Chronic Constriction Injury via the Keap1–Nrf2 Pathway. Front. Cell Dev. Biol. 2021, 9, 714996. [Google Scholar] [CrossRef]
- Shan, W.; Liao, X.; Tang, Y.; Liu, J. Dexmedetomidine alleviates inflammation in neuropathic pain by suppressing NLRP3 via Nrf2 activation. Exp. Ther. Med. 2021, 22, 1–9. [Google Scholar] [CrossRef]
- Riego, G.; Redondo, A.; Leánez, S.; Pol, O. Mechanism implicated in the anti-allodynic and anti-hyperalgesic effects induced by the activation of heme oxygenase 1/carbon monoxide signaling pathway in the central nervous system of mice with neuropathic pain. Biochem. Pharmacol. 2018, 148, 52–63. [Google Scholar] [CrossRef]
- Shan, Y.; Lambrecht, R.W.; Donohue, S.E.; Bonkovsky, H.L.; Shan, Y.; Lambrecht, R.W.; Donohue, S.E.; Bonkovsky, H.L. Role of Bach1 and Nrf2 in up-regulation of the heme oxygenase-1 gene by cobalt protoporphyrin. FASEB J. 2006, 20, 2651–2653. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Leánez, S.; Pol, O. Sulforaphane inhibited the nociceptive responses, anxiety-and depressive-like behaviors associated with neuropathic pain and improved the anti-allodynic effects of morphine in mice. Front. Pharmacol. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Pol, O. Treatment with 5-fluoro-2-oxindole increases the antinociceptive effects of morphine and inhibits neuropathic pain. Cell. Mol. Neurobiol. 2021, 41, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, C. Anti-nociceptive and anti-inflammatory actions of sulforaphane in chronic constriction injury-induced neuropathic pain mice. Inflammopharmacology 2017, 25, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Green-Fulgham, S.M.; Harland, M.E.; Ball, J.B.; D’Angelo, H.; Dreher, R.A.; Li, J.; Lacagnina, M.J.; Lorca, S.A.; Kwilasz, A.J.; Maier, S.F. Preconditioning by voluntary wheel running attenuates later neuropathic pain via Nrf2 antioxidant signaling in rats. bioRxiv 2021. [Google Scholar] [CrossRef]
- Robinson, L.R. Traumatic injury to peripheral nerves. Muscle Nerve 2000, 23, 863–873. [Google Scholar] [CrossRef]
- Zhang, L.; Johnson, D.; Johnson, J.A. Deletion of Nrf2 impairs functional recovery, reduces clearance of myelin debris and decreases axonal remyelination after peripheral nerve injury. Neurobiol. Dis. 2013, 54, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, W.; Deng, B.; Duan, W.; Li, Y.; Liu, Y.; Li, Z.; Xia, W.; Li, C. Schwann Cell Plasticity is Regulated by a Weakened Intrinsic Antioxidant Defense System in Acute Peripheral Nerve Injury. Neuroscience 2018, 382, 1–13. [Google Scholar] [CrossRef]
- Qiu, J.; Yang, X.; Wang, L.; Zhang, Q.; Ma, W.; Huang, Z.; Bao, Y.; Zhong, L.; Sun, H.; Ding, F. Isoquercitrin promotes peripheral nerve regeneration through inhibiting oxidative stress following sciatic crush injury in mice. Ann. Transl. Med. 2019, 7, 680. [Google Scholar] [CrossRef]
- Caillaud, M.; Chantemargue, B.; Richard, L.; Vignaud, L.; Favreau, F.; Faye, P.-A.; Vignoles, P.; Sturtz, F.; Trouillas, P.; Vallat, J.-M.; et al. Local low dose curcumin treatment improves functional recovery and remyelination in a rat model of sciatic nerve crush through inhibition of oxidative stress. Neuropharmacology 2018, 139, 98–116. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Huang, H.-C.; Wu, P.-T.; Tai, T.-W.; Jou, I.M. Sesame oil improves functional recovery by attenuating nerve oxidative stress in a mouse model of acute peripheral nerve injury: Role of Nrf-2. J. Nutr. Biochem. 2016, 38, 102–106. [Google Scholar] [CrossRef]
- Khan, A.; Khan, A.; Khalid, S.; Shal, B.; Kang, E.; Lee, H.; Laumet, G.; Seo, E.K.; Khan, S. 7β-(3-Ethyl-cis-crotonoyloxy)-1α-(2-methylbutyryloxy)-3,14-dehydro-Z Notonipetranone Attenuates Neuropathic Pain by Suppressing Oxidative Stress, Inflammatory and Pro-Apoptotic Protein Expressions. Molecules 2021, 26, 181. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, X.; Tao, X.; Zhang, B.; Sun, C.; Wang, P.; Song, T. Sirt2 in the spinal cord regulates chronic neuropathic pain through Nrf2-mediated oxidative stress pathway in rats. Front. Pharmacol. 2021, 12, 646477. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, J.; Lacagnina, M.J.; Lorca, S.; Odem, M.A.; Walters, E.T.; Kavelaars, A.; Grace, P.M. Oral dimethyl fumarate reduces peripheral neuropathic pain in rodents via NFE2L2 antioxidant signaling. Anesthesiology 2020, 132, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, C.; Fang, X.; Zhan, G.; Huang, N.; Gao, J.; Xu, H.; Hashimoto, K.; Luo, A. Role of Keap1-Nrf2 Signaling in Anhedonia Symptoms in a Rat Model of Chronic Neuropathic Pain: Improvement with Sulforaphane. Front. Pharmacol. 2018, 9, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Yu, L.; Lin, J.; Ma, L.; Gao, D.S.; Sun, N.; Liu, Y.; Fang, L.; Cheng, Z.; Sun, K. Itaconate as a Promising Therapy for Neuropathic Pain and Inflammatory Pain. Res. Square 2021, 1–25. [Google Scholar] [CrossRef]
- Seltzer, Z.e.; Dubner, R.; Shir, Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain 1990, 43, 205–218. [Google Scholar] [CrossRef]
- De Logu, F.; Nassini, R.; Materazzi, S.; Carvalho Gonçalves, M.; Nosi, D.; Rossi Degl’Innocenti, D.; Marone, I.M.; Ferreira, J.; Li Puma, S.; Benemei, S.; et al. Schwann cell TRPA1 mediates neuroinflammation that sustains macrophage-dependent neuropathic pain in mice. Nat. Commun. 2017, 8, 1887. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Satoh, Y.; Otsubo, Y.; Kazama, T. Molecular hydrogen attenuates neuropathic pain in mice. PLoS ONE 2014, 9, e100352. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.V.; Deumens, R.; Goursaud, S.; Schäfer, S.; Lavand’homme, P.; Joosten, E.A.; Hermans, E. Enhanced neuroinflammation and pain hypersensitivity after peripheral nerve injury in rats expressing mutated superoxide dismutase 1. J. Neuroinflam. 2011, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, K.; Kawamoto, A.; Isami, K.; Maeda, S.; Kusano, A.; Asakura, K.; Shirakawa, H.; Mori, Y.; Nakagawa, T.; Kaneko, S. TRPM2 contributes to inflammatory and neuropathic pain through the aggravation of pronociceptive inflammatory responses in mice. J. Neurosci. 2012, 32, 3931–3941. [Google Scholar] [CrossRef] [Green Version]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Shields, S.D.; Eckert III, W.A.; Basbaum, A.I. Spared nerve injury model of neuropathic pain in the mouse: A behavioral and anatomic analysis. J. Pain 2003, 4, 465–470. [Google Scholar] [CrossRef]
- North, B.J.; Verdin, E. Sirtuins: Sir2-related NAD-dependent protein deacetylases. Genome Biol. 2004, 5, 224. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The role of sirtuins in antioxidant and redox signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Parodi, B.; Sanna, A.; Cedola, A.; Uccelli, A.; Kerlero de Rosbo, N. Hydroxycarboxylic Acid Receptor 2, a Pleiotropically Linked Receptor for the Multiple Sclerosis Drug, Monomethyl Fumarate. Possible Implications for the Inflammatory Response. Front. Immunol. 2021, 12, 1921. [Google Scholar] [CrossRef] [PubMed]
- Parodi, B.; Rossi, S.; Morando, S.; Cordano, C.; Bragoni, A.; Motta, C.; Usai, C.; Wipke, B.; Scannevin, R.; Mancardi, G.; et al. Fumarates modulate microglia activation through a novel HCAR2 signaling pathway and rescue synaptic dysregulation in inflamed CNS. Acta Neuropathol. 2015, 130, 279–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Assmann, J.C.; Krenz, A.; Rahman, M.; Grimm, M.; Karsten, C.M.; Köhl, J.; Offermanns, S.; Wettschureck, N.; Schwaninger, M. Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J. Clin. Investig. 2014, 124, 2188–2192. [Google Scholar] [CrossRef] [PubMed]
- Boccella, S.; Guida, F.; De Logu, F.; De Gregorio, D.; Mazzitelli, M.; Belardo, C.; Iannotta, M.; Serra, N.; Nassini, R.; de Novellis, V.; et al. Ketones and pain: Unexplored role of hydroxyl carboxylic acid receptor type 2 in the pathophysiology of neuropathic pain. FASEB J. 2019, 33, 1062–1073. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Mogil, J.S.; Wilson, S.G.; Bon, K.; Lee, S.E.; Chung, K.; Raber, P.; Pieper, J.O.; Hain, H.S.; Belknap, J.K.; Hubert, L. Heritability of nociception I: Responses of 11 inbred mouse strains on 12 measures of nociception. Pain 1999, 80, 67–82. [Google Scholar] [CrossRef]
- Li, J.; Tian, M.; Hua, T.; Wang, H.; Yang, M.; Li, W.; Zhang, X.; Yuan, H. Combination of autophagy and NFE2L2/NRF2 activation as a treatment approach for neuropathic pain. Autophagy 2021, 17, 4062–4082. [Google Scholar] [CrossRef] [PubMed]
- Sorge, R.E.; Totsch, S.K. Sex differences in pain. J. Neurosci. Res. 2017, 95, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Rosen, S.; Ham, B.; Mogil, J.S. Sex differences in neuroimmunity and pain. J. Neurosci. Res. 2017, 95, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Plesh, O.; Adams, S.H.; Gansky, S.A. Racial/ethnic and gender prevalences in reported common pains in a national sample. J. Orofac. Pain 2011, 25, 25. [Google Scholar]
- Dance, A. Why the sexes don’t feel pain the same way. Nature 2019, 567, 448–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.R.; Chin, M.P.; Delatycki, M.B.; Subramony, S.H.; Corti, M.; Hoyle, J.C.; Boesch, S.; Nachbauer, W.; Mariotti, C.; Mathews, K.D.; et al. Safety and Efficacy of Omaveloxolone in Friedreich Ataxia (MOXIe Study). Ann. Neurol. 2021, 89, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Bauman, J.E.; Zang, Y.; Sen, M.; Li, C.; Wang, L.; Egner, P.A.; Fahey, J.W.; Normolle, D.P.; Grandis, J.R.; Kensler, T.W.; et al. Prevention of Carcinogen-Induced Oral Cancer by Sulforaphane. Cancer Prev. Res. 2016, 9, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Zager, R.A.; Johnson, A.C.M. The NRF2 stimulating agent, tin protoporphyrin, activates protective cytokine pathways in healthy human subjects and in patients with chronic kidney disease. Physiol. Rep. 2020, 8, e14566. [Google Scholar] [CrossRef]

| 5-fluoro-2-oxindole | Alphalipoic Acid | Bardoxolone Methyl | Berberine |
|---|---|---|---|
 |  |  |  |
| Bergenin | Bromelain | Carvacrol | Curcumin |
 |  |  |  |
| Deguelin | Dexmedetomidine | Diallyl Disulfide | Diallyl Trisulfide |
 |  |  |  |
| Dimethyl Fumarate | Dimethylitaconate | Diosgenin | Diphenyl Diselenide |
 |  |  |  |
| Fisetin | Formononetin | Isoliquiritigenin | Isoquercitrin |
 |  |  |  |
| L-carnosine | Levo-corydalmine | Mitoquinone | Monomethyl Fumarate |
 |  |  |  |
| Oleuropein | Oltipraz | Paeoniflorin | Plumbagin |
 |  |  |  |
| Polydatin | Quercetin | Resveratrol | Rosiglitazone |
 |  |  |  |
| RTA-408 | Rutin | Sulforaphane | Tanshinone IIA |
 |  |  |  |
| Taurine | tBHQ | UFP-512 | |
 |  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basu, P.; Averitt, D.L.; Maier, C.; Basu, A. The Effects of Nuclear Factor Erythroid 2 (NFE2)-Related Factor 2 (Nrf2) Activation in Preclinical Models of Peripheral Neuropathic Pain. Antioxidants 2022, 11, 430. https://doi.org/10.3390/antiox11020430
Basu P, Averitt DL, Maier C, Basu A. The Effects of Nuclear Factor Erythroid 2 (NFE2)-Related Factor 2 (Nrf2) Activation in Preclinical Models of Peripheral Neuropathic Pain. Antioxidants. 2022; 11(2):430. https://doi.org/10.3390/antiox11020430
Chicago/Turabian StyleBasu, Paramita, Dayna L. Averitt, Camelia Maier, and Arpita Basu. 2022. "The Effects of Nuclear Factor Erythroid 2 (NFE2)-Related Factor 2 (Nrf2) Activation in Preclinical Models of Peripheral Neuropathic Pain" Antioxidants 11, no. 2: 430. https://doi.org/10.3390/antiox11020430