

L-Arginine Alleviates the Reduction in Photosynthesis and Antioxidant Activity Induced by Drought Stress in Maize Seedlings
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Vegetative Growth and Relative Leaf Water Content
2.4. Arginine Content
2.5. Ultrastructure of the Chloroplasts
2.6. Photosynthetic Leaf Pigment
2.7. Leaf SPAD and Chlorophyll Fluorescence Parameters
2.8. Oxidative Stress Markers
2.9. Activity of Antioxidant Enzymes
2.10. Statistical Analysis
3. Results
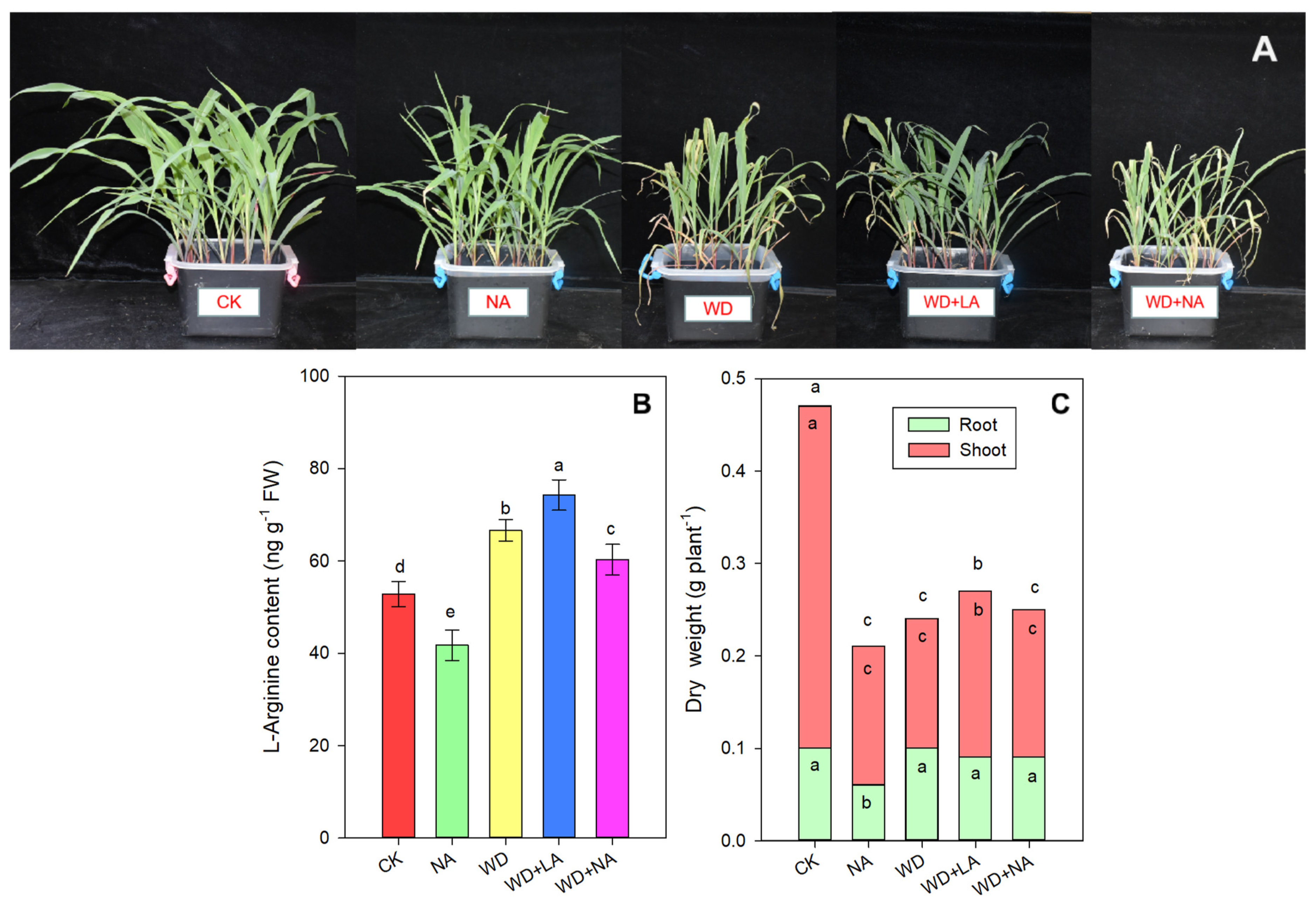
3.1. L-Arginine Alleviated the Drought-Induced Growth Inhibition of Maize Seedlings
3.2. Effects of L-Arginine on Photosynthetic Performance under Drought
3.2.1. Leaf Water Conditions and Leaf Area
3.2.2. Ultrastructure of the Chloroplasts
3.2.3. Photosynthetic Performance
3.3. Effect of L-Arginine on the Membrane System and Antioxidant Activity under Drought Stress
3.3.1. H2O2, O2•−, and MDA
3.3.2. Antioxidant Enzyme Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hao, Z.; Singh, V.P. Drought Characterization from a Multivariate Perspective: A Review. J. Hydrol. 2015, 527, 668–678. [Google Scholar] [CrossRef]
- Zhang, Q.; Yao, Y.; Li, Y.; Huang, J.; Ma, Z.; Wang, Z.; Wang, S.; Wang, Y.; Zhang, Y. Causes and Changes of Drought in China: Research Progress and Prospects. J. Meteorol. Res. 2020, 34, 460–481. [Google Scholar] [CrossRef]
- Kaur, M.; Sood, S.K. Hydro-Meteorological Hazards and Role of ICT during 2010-2019: A Scientometric Analysis. Earth Sci. Inf. 2020, 13, 1201–1223. [Google Scholar] [CrossRef]
- Nitoi, M. Investigating the Recurrence of Meteorological Hazards. Prog. Nucl. Energy 2016, 89, 39–45. [Google Scholar] [CrossRef]
- Kim, W.; Iizumi, T.; Nishimori, M. Global Patterns of Crop Production Losses Associated with Droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Su, B.; Huang, J.; Fischer, T.; Wang, Y.; Kundzewicz, Z.W.; Zhai, J.; Sun, H.; Wang, A.; Zeng, X.; Wang, G.; et al. Drought Losses in China Might Double between the 1.5 °C and 2.0 °C Warming. Proc. Natl. Acad. Sci. USA 2018, 115, 10600–10605. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Wang, S.-Q.; Wang, J.-B.; Lu, H.-Q.; Guo, A.-H.; Zhu, Z.-C.; Myneni, R.B.; Shugart, H.H. Assessing Spatiotemporal Variation of Drought in China and Its Impact on Agriculture during 1982–2011 by Using PDSI Indices and Agriculture Drought Survey Data. J. Geophys. Res. Atmos. 2016, 121, 2283–2298. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic Variation in Growth and Physiological Response to Drought Stress and Re-Watering Reveals the Critical Role of Recovery in Drought Adaptation in Maize Seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef]
- Ma, X.; He, Q.; Zhou, G. Sequence of Changes in Maize Responding to Soil Water Deficit and Related Critical Thresholds. Front. Plant Sci. 2018, 9, 511. [Google Scholar] [CrossRef] [PubMed]
- Sadhukhan, A.; Prasad, S.S.; Mitra, J.; Siddiqui, N.; Sahoo, L.; Kobayashi, Y.; Koyama, H. How Do Plants Remember Drought? Planta 2022, 256, 7. [Google Scholar] [CrossRef]
- Zi, X.; Zhou, S.; Wu, B. Alpha-Linolenic Acid Mediates Diverse Drought Responses in Maize (Zea mays, L.) at Seedling and Flowering Stages. Molecules 2022, 27, 771. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Wang, G.-Y.; Muhammad, I.; Chi, Y.-X.; Zeeshan, M.; Nasar, J.; Zhou, X.-B. Interactive Effects of Melatonin and Nitrogen Improve Drought Tolerance of Maize Seedlings by Regulating Growth and Physiochemical Attributes. Antioxidants 2022, 11, 359. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin Improves Drought Stress Tolerance of Tomato by Modulating Plant Growth, Root Architecture, Photosynthesis, and Antioxidant Defense System. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Butt, M.; Irfan, M.; Rizwan, M.; Ali, H.; Cheema, M. Interactive Effect of Biochar and Silicon on Improving Morpho-Physiological and Biochemical Attributes of Maize by Reducing Drought Hazards. J. Soil Sci. Plant Nutr. 2020, 20, 1819–1826. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.; Xiemuxiding, A.; Zhang, X.; Duan, L.; Li, R. Fulvic Acid, Brassinolide, and Uniconazole Mediated Regulation of Morphological and Physiological Traits in Maize Seedlings Under Water Stress. J. Plant Growth Regul. 2022, 1–13. [Google Scholar] [CrossRef]
- Yang, L.; Fountain, J.C.; Wang, H.; Ni, X.; Ji, P.; Lee, R.D.; Kemerait, R.C.; Scully, B.T.; Guo, B. Stress Sensitivity Is Associated with Differential Accumulation of Reactive Oxygen and Nitrogen Species in Maize Genotypes with Contrasting Levels of Drought Tolerance. Int. J. Mol. Sci. 2015, 16, 24791–24819. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of Drought Stress and Water Recovery on Physiological Responses and Gene Expression in Maize Seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine Metabolism: Boundaries of Our Knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef]
- Patel, P.; Kadur Narayanaswamy, G.; Kataria, S.; Baghel, L. Involvement of Nitric Oxide in Enhanced Germination and Seedling Growth of Magnetoprimed Maize Seeds. Plant Signal Behav. 2017, 12, e1293217. [Google Scholar] [CrossRef]
- Shu, P.; Min, D.; Ai, W.; Li, J.; Zhou, J.; Li, Z.; Zhang, X.; Shi, Z.; Sun, Y.; Jiang, Y.; et al. L-Arginine Treatment Attenuates Postharvest Decay and Maintains Quality of Strawberry Fruit by Promoting Nitric Oxide Synthase Pathway. Postharvest Biol. Technol. 2020, 168, 111253. [Google Scholar] [CrossRef]
- Li, B.; Ding, Y.; Tang, X.; Wang, G.; Tang, X. Effect of L-Arginine on Maintaining Storage Quality of the White Button Mushroom (Agaricus bisporus). Food Bioprocess Technol. 2019, 12, 563–574. [Google Scholar] [CrossRef]
- Babalar, M.; Pirzad, F.; Sarcheshmeh, M.A.A.; Talaei, A.; Lessani, H. Arginine Treatment Attenuates Chilling Injury of Pomegranate Fruit during Cold Storage by Enhancing Antioxidant System Activity. Postharvest Biol. Technol. 2018, 137, 31–37. [Google Scholar] [CrossRef]
- Hassan, M.M. Hala Ezzat L-Arginine Pretreatment Enhances Drought Resistance of Sunflower (Helianthus annuus, L.) Plants by Increase in Polyamines Content. J. Plant Growth Regul. 2019, 38, 600–605. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Luo, Z.; Jannatizadeh, A.; Sheikh-Assadi, M.; Sharafi, Y.; Farmani, B.; Fard, J.R.; Razavi, F. Employing Exogenous Melatonin Applying Confers Chilling Tolerance in Tomato Fruits by Upregulating ZAT2/6/12 Giving Rise to Promoting Endogenous Polyamines, Proline, and Nitric Oxide Accumulation by Triggering Arginine Pathway Activity. Food Chem. 2019, 275, 549–556. [Google Scholar] [CrossRef]
- Wills, R.B.H.; Li, Y. Use of Arginine to Inhibit Browning on Fresh Cut Apple and Lettuce. Postharvest Biol. Technol. 2016, 113, 66–68. [Google Scholar] [CrossRef]
- Tossi, V.; Cassia, R.; Lamattina, L. Apocynin-Induced Nitric Oxide Production Confers Antioxidant Protection in Maize Leaves. J. Plant Physiol. 2009, 166, 1336–1341. [Google Scholar] [CrossRef]
- Shao, R.; Zhang, J.; Shi, W.; Wang, Y.; Tang, Y.; Liu, Z.; Sun, W.; Wang, H.; Guo, J.; Meng, Y.; et al. Mercury Stress Tolerance in Wheat and Maize Is Achieved by Lignin Accumulation Controlled by Nitric Oxide. Environ. Pollut. 2022, 307, 119488. [Google Scholar] [CrossRef]
- Zhuang, J.; Zhou, L.; Wang, Y.; Chi, Y. Nitrogen Allocation Regulates the Relationship between Maximum Carboxylation Rate and Chlorophyll Content along the Vertical Gradient of Subtropical Forest Canopy. Agric. For. Meteorol. 2021, 307, 108512. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, H.; Raj, S.; Soni, V. Chlorophyll a Fluorescence Kinetics of Mung Bean (Vigna radiata, L.) Grown under Artificial Continuous Light. Biochem. Biophys. Rep. 2020, 24, 100813. [Google Scholar] [CrossRef]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological Implications of Arginine Metabolism in Plants. Front Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [Green Version]
- El-Bassiouny, H.; Mostafa, H.A.; El-Khawas, S.A.; Hassanein, R.A.; Khalil, S.I.; El-Monem, A.A. Physiological Responses of Wheat Plant to Foliar Treatments with Arginine or Putrescine. Aust. J. Basic Appl. Sci. 2008, 2, 1390–1403. [Google Scholar]
- Nasibi, F.; Yaghoobi, M.; Kalantari, K. Effect of Exogenous Arginine on Alleviation of Oxidative Damage in Tomato Plant Underwater Stress. J. Plant Interact. 2011, 6, 291–296. [Google Scholar] [CrossRef]
- Hamid, Z.H.; Amer, M.; Wahab, A. Effect of Arginine on Growth and Yield of Tomato Plant (Lycopersicon esculentum) under Drought Stress. Plant Arch. 2019, 19, 4441–4444. [Google Scholar]
- Matysiak, K.; Kierzek, R.; Siatkowski, I.; Kowalska, J.; Krawczyk, R.; Miziniak, W. Effect of Exogenous Application of Amino Acids L-Arginine and Glycine on Maize under Temperature Stress. Agronomy 2020, 10, 769. [Google Scholar] [CrossRef]
- Zeid, I.M. Effect of Arginine and Urea on Polyamines Content and Growth of Bean under Salinity Stress. Acta Physiol. Plant. 2009, 31, 65–70. [Google Scholar] [CrossRef]
- Hummel, I.; Gouesbet, G.; El Amrani, A.; Aïnouche, A.; Couée, I. Characterization of the Two Arginine Decarboxylase (Polyamine biosynthesis) Paralogues of the Endemic Subantarctic Cruciferous Species Pringlea antiscorbutica and Analysis of Their Differential Expression during Development and Response to Environmental Stress. Gene 2004, 342, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Nejadalimoradi, H.; Nasibi, F.; Khosrow, M.; Kalantari; Zanganeh, R. Effect of Seed Priming with L-Arginine and Sodium Nitroprusside on Some Physiological Parameters and Antioxidant Enzymes of Sunflower Plants Exposed to Salt Stress. Agric. Commun. 2014, 2, 23–30. [Google Scholar]
- García-Mata, C.; Lamattina, L. Nitric Oxide Induces Stomatal Closure and Enhances the Adaptive Plant Responses against Drought Stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Guo, F.-Q. New Insights into Nitric Oxide Metabolism and Regulatory Functions. Trends Plant Sci. 2005, 10, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Xu, X.M.; Cui, J. The Importance of Blue Light for Leaf Area Expansion, Development of Photosynthetic Apparatus, and Chloroplast Ultrastructure of Cucumis sativus Grown under Weak Light. Photosynthetica 2015, 53, 213–222. [Google Scholar] [CrossRef]
- Zargar, S.M.; Gupta, N.; Nazir, M.; Mahajan, R.; Malik, F.A.; Sofi, N.R.; Shikari, A.B.; Salgotra, R.K. Impact of Drought on Photosynthesis: Molecular Perspective. Plant Gene 2017, 11, 154–159. [Google Scholar] [CrossRef]
- Nasibi, F.; Heidari, T.; Asrar, Z.; Mansoori, H. Effect of Arginine Pre-Treatment on Nickel Accumulation and Alleviation of the Oxidative Stress in Hyoscyamus niger. J. Soil Sci. Plant Nutr. 2013, 13, 680–689. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The Relationship between Leaf Area Growth and Biomass Accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical Gains in Soybean (Glycine max Merr.) Seed Yield Are Driven by Linear Increases in Light Interception, Energy Conversion, and Partitioning Efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef]
- Tezara, W.; Mitchell, V.; Driscoll, S.; Lawlor, D. Water Stress Inhibits Plant Photosynthesis by Decreasing Coupling Factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Kume, A.; Akitsu, T.; Nasahara, K.N. Why Is Chlorophyll b Only Used in Light-Harvesting Systems? J. Plant Res. 2018, 131, 961–972. [Google Scholar] [CrossRef]
- Björn, L.O.; Papageorgiou, G.C.; Blankenship, R.E.; Govindjee. A Viewpoint: Why Chlorophyll a? Photosynth. Res. 2009, 99, 85–98. [Google Scholar] [CrossRef]
- Shalaby, M.A.; Ahmed, M.A.; Khater, M.A. Physiological Responses of Some Barley Cultivars to Foliar Treatments with Arginine under Water Stress Conditions. Middle East J. Agric. Res. 2018, 7, 1102–1123. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous Nitric Oxide Donor and Arginine Provide Protection against Short-Term Drought Stress in Wheat Seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef]
- Zhang, R.H.; Zhang, X.H.; Camberato, J.; Xue, J.Q. Photosynthetic Performance of Maize Hybrids to Drought Stress. Russ. J. Plant Physiol. 2015, 62, 788–796. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Stöckel, J.; Srivastava, A.; Strasser, R.J. Multiple Effects of Chromate on the Photosynthetic Apparatus of Spirodela polyrhiza as Probed by OJIP Chlorophyll a Fluorescence Measurements. Environ. Pollut. 2001, 115, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Song, L.U. Application of Chlorophyll Fluorescence Kinetics in Researches on Plant Stress Resistance. Sclykj 2013, 34, 69–71. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Stoyanova, D.; Tchakalova, E.; Yordanov, I. Influence of Different Soil Moisture on Anatomy of Maize Leaves and Ultrastructure of Chloroplasts. J. Plant Physiol. 2002, 28, 11–20. [Google Scholar]
- Vani, B.; Pardha Saradhi, P.; Mohanty, P. Alteration in Chloroplast Structure and Thylakoid Membrane Composition Due to in Vivo Heat Treatment of Rice Seedlings: Correlation with the Functional Changes. J. Plant Physiol. 2001, 158, 583–592. [Google Scholar] [CrossRef]
- Meher; Shivakrishna, P.; Ashok Reddy, K.; Manohar Rao, D. Effect of PEG-6000 Imposed Drought Stress on RNA Content, Relative Water Content (RWC), and Chlorophyll Content in Peanut Leaves and Roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar] [CrossRef]
- Nasibi, F.; Kalantari, K.M.; Barand, A. Effect of Seed Pre-Treatment with L-Arginine on Improvement of Seedling Growth and Alleviation of Oxidative Damage in Canola Plants Subjected to Salt Stress. Iran J. Plant Physiol. 2015, 5, 1217–1224. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of Enzymatic and Nonenzymatic Antioxidants in Plants during Abiotic Stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant Growth under Water/Salt Stress: ROS Production; Antioxidants and Significance of Added Potassium under Such Conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef]
- Yang, P.M.; Huang, Q.C.; Qin, G.Y.; Zhao, S.P.; Zhou, J.G. Different Drought-Stress Responses in Photosynthesis and Reactive Oxygen Metabolism between Autotetraploid and Diploid Rice. Photosynthetica 2014, 52, 193–202. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved Drought Tolerance by AMF Inoculation in Maize (Zea mays) Involves Physiological and Biochemical Implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.; Hu, C.; Chen, X.; Sun, X.; Xu, N. Physiological and Transcriptome Analysis of Exogenous L-Arginine in the Alleviation of High-Temperature Stress in Gracilariopsis lemaneiformis. Front. Mar. Sci. 2021, 08, 784586. [Google Scholar] [CrossRef]
- Nasr, F.; Razavi, F.; Rabiei, V.; Gohari, G.; Ali, S.; Hano, C. Attenuation of Chilling Injury and Improving Antioxidant Capacity of Persimmon Fruit by Arginine Application. Foods 2022, 11, 2419. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Miao, F.; Wang, Y.; Liu, H.; Wang, X.; Wang, H.; Guo, J.; Shao, R.; Yang, Q. L-Arginine Alleviates the Reduction in Photosynthesis and Antioxidant Activity Induced by Drought Stress in Maize Seedlings. Antioxidants 2023, 12, 482. https://doi.org/10.3390/antiox12020482
Sun Y, Miao F, Wang Y, Liu H, Wang X, Wang H, Guo J, Shao R, Yang Q. L-Arginine Alleviates the Reduction in Photosynthesis and Antioxidant Activity Induced by Drought Stress in Maize Seedlings. Antioxidants. 2023; 12(2):482. https://doi.org/10.3390/antiox12020482
Chicago/Turabian StyleSun, Yifei, Feng Miao, Yongchao Wang, Hecheng Liu, Xintao Wang, Hao Wang, Jiameng Guo, Ruixin Shao, and Qinghua Yang. 2023. "L-Arginine Alleviates the Reduction in Photosynthesis and Antioxidant Activity Induced by Drought Stress in Maize Seedlings" Antioxidants 12, no. 2: 482. https://doi.org/10.3390/antiox12020482