
Natural Products in Renal-Associated Drug Discovery

1
Institute for Stem Cell Research and Regenerative Medicine, Medical Faculty, Heinrich Heine University, Moorenstr. 5, 40225 Düsseldorf, Germany
2
Department of Biochemistry and Molecular Biology, Faculty of Biosciences, University for Development Studies, Nyankpala P.O. Box TL 1882, Ghana
3
EGA Institute for Women’s Health, Zayed Centre for Research into Rare Diseases in Children (ZCR), University College London (UCL), 20 Guilford Street, London WC1N 1DZ, UK
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Antioxidants 2023, 12(8), 1599; https://doi.org/10.3390/antiox12081599
Submission received: 21 July 2023
/
Accepted: 8 August 2023
/
Published: 11 August 2023
(This article belongs to the Special Issue Antioxidant Research in Germany)
Abstract
:The global increase in the incidence of kidney failure constitutes a major public health problem. Kidney disease is classified into acute and chronic: acute kidney injury (AKI) is associated with an abrupt decline in kidney function and chronic kidney disease (CKD) with chronic renal failure for more than three months. Although both kidney syndromes are multifactorial, inflammation and oxidative stress play major roles in the diversity of processes leading to these kidney malfunctions. Here, we reviewed various publications on medicinal plants with antioxidant and anti-inflammatory properties with the potential to treat and manage kidney-associated diseases in rodent models. Additionally, we conducted a meta-analysis to identify gene signatures and associated biological processes perturbed in human and mouse cells treated with antioxidants such as epigallocatechin gallate (EGCG), the active ingredient in green tea, and the mushroom Ganoderma lucidum (GL) and in kidney disease rodent models. We identified EGCG- and GL-regulated gene signatures linked to metabolism; inflammation (NRG1, E2F1, NFKB1 and JUN); ion signalling; transport; renal processes (SLC12A1 and LOX) and VEGF, ERBB and BDNF signalling. Medicinal plant extracts are proving to be effective for the prevention, management and treatment of kidney-associated diseases; however, more detailed characterisations of their targets are needed to enable more trust in their application in the management of kidney-associated diseases.
1. Introduction
Chronic kidney disease (CKD) and acute kidney injury (AKI) have emerged as major public health burdens with close connections to each other, AKI as a risk factor for CKD and vice versa and both increasing the risk of cardiovascular disease [1]. CKD is defined by a low glomerular filtration rate (GFR) or the presence of kidney damage for more than 3 months [2,3]. Proteins in urine (proteinuria) and decreased GFR as indicators of kidney damage directly reflect the physical properties of the filter between blood and urine constituted by an endothelial layer, the glomerular basement membrane (GMB) and podocytes [4]. While, under physiological conditions, most proteins cannot traverse this barrier, in proteinuria, larger proteins such as albumin, immunoglobulins G and M and α1-microglobulin and β2-microglobulin, correlating with the severity of the histologic lesions [5], can. These proteins can, as a consequence, impair the reabsorption of other, smaller molecules by proximal tubular epithelial cells and ultimately lead to toxic damage [5]. As risk factors of CKD, there has been a global rise in incidences of diabetes and hypertension [6,7].
AKI is defined by a rapid increase in serum creatinine concentrations and/or decline of urine output. The number of incidences is approximately 10–15% of patients admitted to hospital and approximately more than 50% in intensive care units [8,9]. Distinct time intervals of endurance of the pathological conditions are used to distinguish between AKI (<7 days), acute kidney disease (AKD; 7 days–90 days) and CKD (>90 days) [9]. AKI is now regarded as a multiorgan dysfunctional disease and classified as prerenal AKI, acute postrenal obstructive nephropathy and intrinsic acute kidney disease, of which only the latter is a true renal disease [10,11].
2. Plant-Based Extracts with Antioxidant and Anti-Inflammatory Properties
Natural products have, for centuries, been used in the management of various disease. Mother nature has served and is still serving us well. Plant-based extracts serve as natural sources of antioxidant and anti-inflammatory agents in combating stress, inflammation and cell death. Numerous oxygen-based metabolic activities generate reactive oxygen species, which can serve as signalling molecules. These signalling molecules serve as precursors of various beneficial events for the body. Increased levels of reactive oxygen species (ROS) and/or reactive nitrogen species (RNS) oxidative stress are an imbalance in the levels of ROS and the body’s natural antioxidant capacity. This creates complications due to ROS reacting with membranes and biomolecules such as lipids and proteins, thus leading to organ damage.
Oxidative stress and inflammation are known to cause various diseases, hence the need to have antioxidants and anti-inflammatory agents that will serve in combating oxidative stress-associated diseases. Increased levels of ROS and RNS are known as potential inducers of kidney injury [12,13,14,15], and molecules associated with ROS and RNS are major regulators of solute and water reabsorption in the kidney [16].
In the assessing and diagnosis of AKI, interleukin-6 (IL-6), interleukin-1 (IL-1), tumour necrosis factor (TNF), adipokines, adhesion molecules and the CD40 ligand are proinflammatory cytokines, which are indicative of the extent of stress or inflammation [17,18,19,20]. Ferguson et al. 2008 [21], Shinke et al. 2015 [22] and Zhou et al. 2008 [23] have also stated the importance of kidney injury molecule-1 (KIM-1) and neutrophil gelatinase-associated lipocalin (NGAL) as additional urinary biomarkers.
Medicinal plants naturally have antioxidant systems that help in combating oxidative stress. Superoxide dismutase (SOD), catalase, glutathione (GSH), which helps in drug metabolism detoxification, and glutathione peroxidase (GPx) are the systems that help in combating oxidative stress. Each medicinal plant extract has its own antioxidant-based mechanism for managing oxidative stress. Some increase the levels of SOD [24] and activate the SOD and catalase levels [25,26], while some plants increase the levels and activities of all the three antioxidant enzymes: SOD, catalase and GPx [27,28].
Natural medicinal plants have showed in vivo and in vitro efficacy in downregulating proinflammatory cytokines and upregulating natural antioxidants such as glutathione, which modulates oxidative stress and inflammation. Though modern treatment approaches have also afforded substantial progress in the fight against AKI, potent therapies are still meagre due to a lack of oxidative stress- and inflammation-specific AKI targets. The cost effects coupled with harsh side effects approaches have led to the search for more novel natural biological products, especially those derived from herbs and natural spices. These can all be accessed in medicinal plants due to the various Phyto components present in them. Their uses and applications are promising, since they are able to react, bind, conjugate and possibly eliminate through excretion all reactive oxygen spices (ROS and RNS), which form the bases of most ailments by cell degradation, lipid peroxidation and inflammation, among others.
3. Plant Sources and Activity
Plants have various phytochemicals with the potential to combat ailments. Research on medicinal plants is being carried out mostly in Asia, followed by Africa and Europe. Below are plants, herbs and spices that have been scientifically proven to manage, protect or cure acute kidney injury.
Aronia melanocarpa of the black chokeberry family mostly found in North America contains anthocyanins (cyanidins), which are able to potentially decrease inflammation, oxidative stress and lipid peroxidation, as well as apoptosis, in acute renal ischaemia effectively [29].
Kang et al. 2021 [30] showed that green tea is rich in bioactive compounds. Its aqueous high content of antioxidants has made it active in managing oxidative stress, resulting in the development of various healthy and nutritious detoxification products.
Punica granatum is a plant originally from India. Administration of the fruit peel ethanolic extract in Wistar rats showed improvement in kidney function biomarkers, exerted antioxidant activity and ameliorated histological changes prerenal and intrinsic gentamicin-induced AKI [31,32].
The methanolic peel extract of passion fruit (Passiflora spp.), which is predominantly found in North America, contains gallic acid, ellagic acid, kaempferol and quercetin glycosides. The extract is able to protect the kidneys by maintaining the levels of urea and creatinine at normal units during paracetamol-induced nephrotoxicity in albino rats [33]. The methanolic extract of its upper parts reduced the urea and creatinine levels during thioacetamide-induced nephrotoxicity in Sprague–Dawley rats [34].
Pistacia atlantica, an exotic berry-like fruit plant, is mostly be found in North Africa, the Middle East, Iran and Afghanistan. Leaf hydroethanolic extracts of Pistacia atlantica have the ability to decrease the levels of urea, creatinine and uric acid during gentamicin-inducted nephrotoxicity in Wistar rats [35].
Eurycoma longifolia is an herbal medicinal plant mostly found in Southeast Asia, Indonesia. The standardised aqueous extract of the roots has been shown to increase the levels and activities of antioxidant enzymes and improves kidney function during paracetamol-induced nephrotoxicity in rats [36].
Costus afer is an African indigenous plant used traditionally for the treatment of several diseases, such as rheumatoid arthritis, hepatic diseases, measles and malaria, and can also serve as an antidote for snake poisoning [37]. The aqueous extract of the leaf has been shown to decrease the serum potassium and BUN levels [38]. It also uses its antioxidant and anti-inflammatory potential to provide neuroprotection against low-dose heavy metal mixed neurotoxicity [39].
Ocimum americanum (family Lamiaceae) grows in Africa, India, China and Southeast Asia and is used as a spice. In Ghana, it is widely cultivated (called akokobesa) [40] and also used by locals to manage diabetes [41]. Nyarko et al. reported that it reduces blood glucose in mice and improved insulin release in beta cells isolated from rats [41]. Genfi et al. reported a hepatoprotective effect of Ocimum americanum, probably due to the inhibition of oxidative stress and the downregulation of proinflammatory cytokines [40].
Cranberry (Vaccinium sp.) natural extracts from North America decrease E. coli adhesion and reduce bacterial motility and biofilm formation in urinary tract infections [42]. Its polyphenols have anti-inflammatory and antioxidant effects and also have positive effects on the gut flora [43].
Descurainia sophia is a dominant weed with several local names and mostly found in Europe and Northern Africa. Csikós et al. 2021 [44] studied its effect on Wistar rats; its aqueous seed extract decreases the deposition of calcium oxalate in ammonium chloride and ethylene glycol-induced gallbladder stones.
An extract of the aerial parts Equisetum arvense, a fern-like plant mostly found in Spain, heals urinary retention and urinary infections, among others [45,46].
The aqueous leaf extract of Anchomanes difformis decreases the levels of oxidative stress-associated biomarkers and increases the CAT and SOD levels in African Wistar rats. It has anti-inflammatory effects by reducing the expression of NF-κB and Bcl2 and decreasing the levels of IL-10, IL-18 and TNF [47].
Hibiscus sabdariffa is a plant used for indigenous beverages in most parts of Asia, Africa and Central America. The aqueous extract of the dried flower bulb contains anthocyanins and chlorogenic acid, which increase both the enzymatic and nonenzymatic antioxidant systems [48].
Curcuma longa, a rhizome, is found mostly in India but now has been planted in Ghana. It contains polyphenol and is used for antioxidant, anti-inflammatory, antimicrobial and antitumour activity, among others [49].
The Lamiaceae family Melissa officinalis (lemon balm) is a well-known herb indigenously used to cure a variety of ailments [50]. It has glycosides that give it antioxidant and cytotoxic properties [51].
Mostly found across Europe is Digitalis purpurea L., a member of the Scrophulariaceae family [52]. The glycosides of D. purpurea have antioxidant and cytotoxic properties. Lycopene, β-carotene and the vitamins of tomato fruits also help to reduce oxidative stress and reduce the risk of cancer [53,54]. Oxidative regulation is paramount in the management of AKI. The aerial parts of Tylophora indica contain alkaloid and tylophorine, which serve as anti-inflammatory and immunosuppressive agents [55]. Lavandula intermedia leaves and flowers have been shown to contain polyphenols, which are significant in providing UV protection [56].
4. Active Ingredients
These plants, herbs and spices serve as antioxidants, anti-inflammatories, anti-malarias, anti-hyperglycaemias and hepatic protectants, among others. This is possible due to the active ingredients in them: flavonoids, alkaloids, saponins, tannins, coumarins, cyanides, anthocyanidins, phenols, phenolics, carotenoids, phytoestrogens [57,58], capsaicin [59,60], curcumin [61], β-carotene [62,63], catechins [64,65], resveratrol [66], vitamins, flavonoids (hyperoxide) and xanthones, as well as naphthodianthrone hypericin (antiviral action), the phloroglucinol derivative hyperforin (antibacterial effect) [67], cardiac glycosides [68], flavonoids, anthraquinones and triterpenes [69,70,71,72].
5. Meta-Analysis of Transcriptome Datasets Related to Natural Products
Table 1 lists datasets found by searches in the public database NCBI GEO (National Center for Biotechnology Information Gene Expression Omnibus). We excluded cancer-related datasets from the results, as we were more interested in the earlier inflammatory processes that may be transferred to AKI and CKD. We found three datasets associated with the same compound, EGCG (epigallocatechin gallate), the active ingredient of green tea, supplemented in human skin and T cells and mouse colon-derived cells. Taking into account the variability of the cell types, our strategy was to identify a gene signature common to skin and T cells. This could then be further refined by identifying conserved genes from mice in the EGCG-treated mouse colon dataset. In the final step, we aimed to transfer this signature to the kidneys by checking overlaps with the kidney datasets, which were made with different compounds but at least with antioxidant properties.
In the EGCG-treated transcriptome datasets (GSE152781) associated with the publication by Ud-Din et al. [73], GSE53448 associated with the publication by Kehrmann et al. [74] and GSE41644 associated with the publication by Barnett et al. [75], we looked for genes up- and downregulated with p < 0.05 (test of the R package limma [76]) and a positive or negative (downregulation) logarithmic fold change when comparing the EGCG treated with the control condition.
5.1. Comparison and Functional Annotation of Differentially Regulated Genes
Differentially up- and downregulated genes were compared in Venn diagrams via the R package VennDiagram [77]. Subsets of the Venn diagrams were analysed for overrepresented gene ontologies (GOs) via the R package GOstats [78]. The most significant GO terms were plotted in a dot plot via the R package ggplot2 [79]. A further enrichment analysis was performed via the R package EnrichR [80].
5.2. Construction of a Gene Regulatory Network
We looked for enrichment of the EGCG down- and upregulated genes with respect to the transcription factor dataset TRANSFAC_and_JASPAR_PWMs in the EnrichR database [80]. The transcription factor enriched for most EGCG-dysregulated genes were subjected to a protein interaction network analysis via STRING-DB [81]. Within the STRING-DB online tool, the network of the uploaded proteins was expanded in one step by five proteins, and the resulting network was analysed for overrepresented Reactome and WikiPathway pathways.
5.3. EGCG Upregulated Gene Signature
The Venn diagram in Figure 1A shows 23 (11 + 12) genes, which were identified as upregulated by EGCG in common in the skin and T cells. Further intersection with EGCG-treated mouse colon cells identified the 11 genes in the centre of the Venn diagram, which were thus supposed to have a conserved response to EGCG. Figure 1B,C show the most significant GO terms overrepresented in the 11 conserved genes and the 23 (11 + 12) human gene signatures. The large overlap between them indicates that most of these biological processes are conserved between men and mice. The significant GO terms can be classified into metabolic (amine and phenol metabolism), ion transport, membrane and secretory categories.
5.4. Gene Signature Downregulated by EGCG
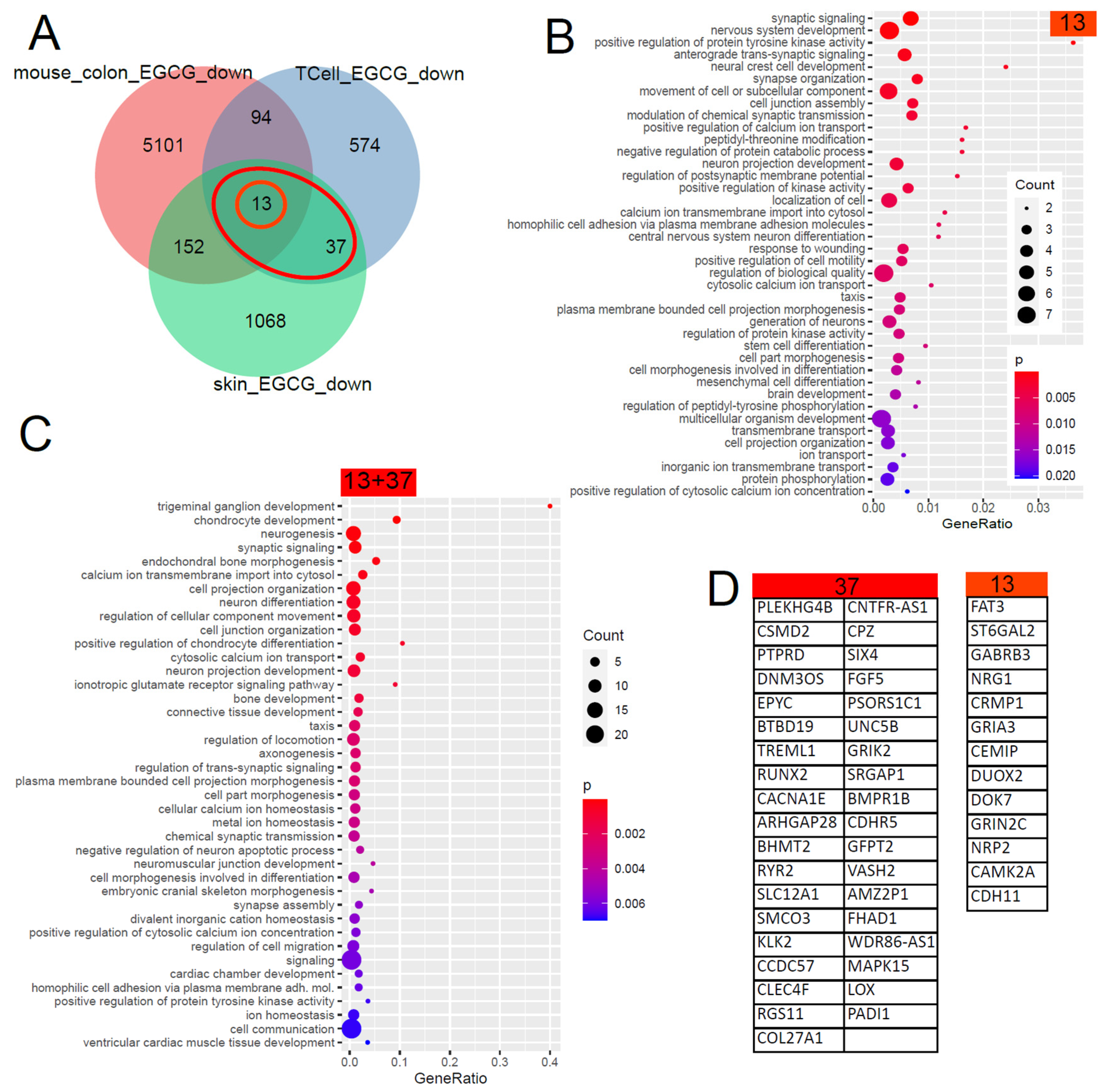
The Venn diagram in Figure 2 shows that 50 (13 + 37) genes were found downregulated by EGCG in common in the skin and in T cells and, with further comparisons with EGCG-treated mouse colons, unveiled 13 conserved genes. The large overlap between both GO analysis results shows that most of these biological processes are conserved between men and mice. The results can be grouped into calcium signalling, neuronal and synaptic, inflammatory (response to wounding), connective tissue, ion transport and taxis categories. With respect to inflammatory processes in kidney diseases, the GO term “response to wounding” found significantly overrepresented in the downregulated EGCG signature (Figure 2B) was relevant. We compared this result with transcriptome data (GSE186823 and GSE171240) from two previous publications in which we investigated kidney organoids treated with the nephrotoxin puromycin aminonucleoside (PAN) [82] and urine-derived podocytes stimulated with angiotensin II (ANG II) [83]. The GO term “response to wounding” was also found significantly overrepresented in the genes downregulated in urine-derived podocytes upon ANGII treatment (p = 0.0015) and kidney organoids upon PAN treatment (p = 0.0354). Among the genes associated with “response to wounding”, we identified NRG1 as overlapping in the downregulated EGCG signature (Figure 2B) and also in the downregulated genes upon PAN treatment. In the corresponding comparison with wounding-related GO terms in the downregulated EGCG signature, no overlapping genes were identified in the ANGII-stimulated podocytes.
5.5. Refined Characterisation and Transcription Factor Analysis of the EGCG Gene Signatures
We further characterised the gene signatures upregulated by EGCG (23, 11 + 12 genes) and downregulated by EGCG (50, 13 + 37 genes) via the R package EnrichR [80]. Figure 3A,B show that the 50 genes downregulated by the EGCG gene signature are overrepresented in the kidney-associated terms “microalbumin in urine” from the UK_Biobank_GWAS_v1 dataset collection and “kidney cancer” from the “Jensen Diseases” dataset collection.
The enrichment analysis revealed that most EGCG-downregulated genes are associated with transcription factor E2F1 (Figure 3C), while most EGCG-upregulated genes are associated with transcription factors JUN and NFKB1 (Figure 3D). The protein interaction network of these transcription factors expanded by five interacting proteins was generated by STRING-DB (Figure 3E) and could be characterised by the Reactome (Figure 3F) and WikiPathway (Figure 3G) pathways related to neuroinflammation and other inflammatory processes.
5.6. Transfer of EGCG-Associated Gene Signatures to Kidney Datasets
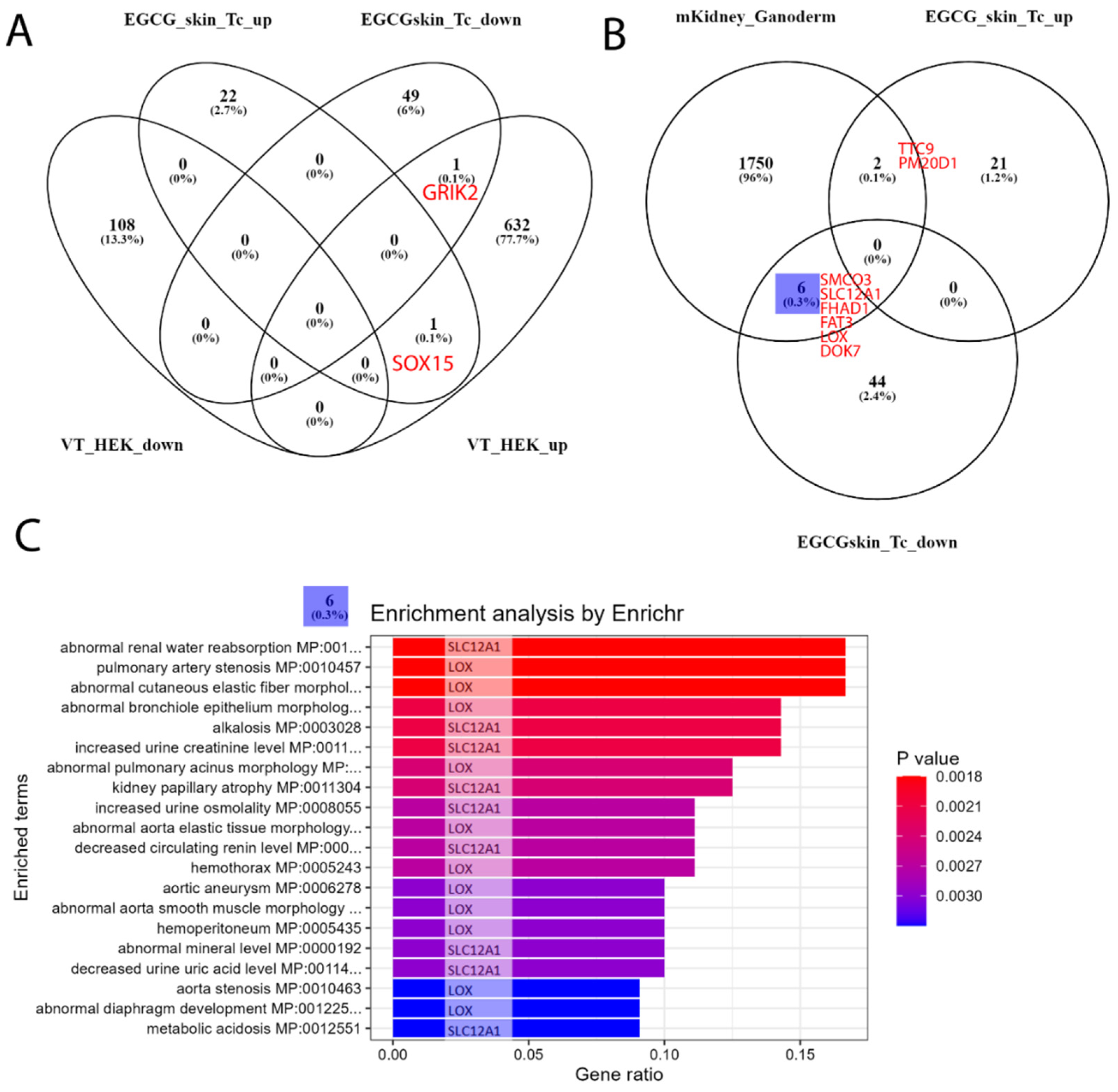
We followed up our analysis to investigate to what extent our EGCG-associated gene signatures are expressed in kidney transcriptome data. First, we used the dataset GSE198890, in which HEK (human embryonic kidney) cells were treated with the compound VT01454 derived from marine natural products. In the associated publication, Li et al. reported activation of the Hippo pathway by VT01454 via phosphatidylinositol transfer proteins α and β (PITPα/β) as the direct molecular targets [84]. The overlap with our EGCG signature (Figure 4A) consisted only of the genes GRIK2 (upregulated by VT01454, downregulated by EGCG) and SOX15 (upregulated by VT01454, upregulated by EGCG). Second, we used the dataset GSE159656 containing transcriptome data of mouse kidneys treated with the mushroom Ganoderma lucidum. Romero-Córdoba et al., the authors of the associated study, reported anti-hypercholesterolemic, hypoglycaemic and antioxidant activities for the compounds derived from the mushroom [85]. The treatment with Ganoderma lucidum resulted in a larger overlap with the EGCG signatures (Figure 4B) of six genes downregulated by EGCG (SMCO3, SLC12A1, FHAD1, FAT3, LOX and DOK7) and two genes upregulated by EGCG (TTC9 and PM20D1). We further subjected the biggest subset of the six genes from the Venn diagram comparison to an enrichment analysis via the R package EnrichR [80] and found several processes related to renal disease (Figure 4C), most significantly “abnormal renal water reabsorption”, which were only each due to one single gene, either SLC12A1 or LOX.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
NCBI GEO datasets used in this transcriptome analysis of natural products.
| GEO_Search | Accession | Cell Type | Treatment | PMID/DOI |
|---|---|---|---|---|
| (Camellia) AND “Homo sapiens” | GSE152781 | human skin | EGCG (green tea) | PMID: 33917842 [73] |
| green tea AND kidney | GSE41644 | Mouse colon | EGCG (green tea) | PMID: 23643524 [75] |
| human T cells EGCG | GSE53448 | human CD4+CD25− T cells | EGCG (green tea) | PMID: 24476360 [74] |
| kidney AND “natural products” | GSE198890 | HEK cells | VT01454 | https://doi.org/10.1038/s41589-022-01061-z (accessed on 1 June 2023) [84] |
| kidney AND “natural products” | GSE159656 | mouse kidney | Ganoderma lucidum | PMID: 33374283 [85] |
5.7. Comparison of Ganoderma Lucidum-Associated Gene Signatures to Datasets of Kidney Disease Models
As mentioned above, we investigated kidney organoids treated with nephrotoxin puromycin aminonucleoside (PAN) and urine-derived podocytes treated with angiotensin II (ANG II) in two previous publications [82,83]. We compared the GL-treated mouse kidney cell-derived gene signatures with the PAN and ANGII (Figure 5B) gene signatures and found the biggest subset of 100 genes overlapping between GL-upregulated and PAN-downregulated (Figure 5A), 36 genes between GL-downregulated and PAN-downregulated (Figure 5A), 90 genes between GL-upregulated and ANGII-downregulated (Figure 5B) and 76 genes between GL-upregulated and ANGII-upregulated (Figure 5B). These subsets were further explored for overrepresented GO terms, of which the dot plots in Figure 5C–F show the most significant terms. In the 100 genes upregulated in GL and downregulated in PAN, these include VEGF-signalling (Figure 5C,D). The 36 genes downregulated in GL and downregulated in PAN are associated with GO terms related to sterol synthesis and blood circulation/pressure (Figure 5D). The most significant GO terms in the 90 genes upregulated in GL and downregulated in ANGII include epithelial cell development, inflammatory processes such as interferon-alpha and leukaemia inhibitory factor responses and ERBB and BDNF signalling (Figure 5E) and, in the 76 genes upregulated in GL and upregulated in ANGII, include ADP synthesis and metabolism, several other metabolic processes and the positive regulation of acute inflammatory processes (Figure 5F).
Taking into account the inflammatory processes playing major roles in the development of CKD and AKI, we focussed on GOs associated with inflammation and wound responses and found an overlap of the GL treatment with the ANGII (Table 2) but not with the PAN treatment. The genes upregulated in the GL treatment and upregulated in the ANGII treatment (C3 and OSMR) are associated with the “positive regulation of the acute inflammatory response” (p = 0.0044), and the genes upregulated in the GL treatment and downregulated in the ANGII treatment (DDR1, DSP, HMOX1, ID3, PLAT, PLEC, USF1, ZFP36 and ZFP36L2) are associated with “response to wounding” (p = 0.0207).
6. Discussion
In this meta-analysis, we first determined the gene signatures in human skin and T cells and mouse colons up- and downregulated by treatment with EGCG, the active ingredient in green tea. The anticarcinogenic, antioxidative and anti-inflammatory properties of EGCG have been shown in a large body of literature [86,87,88]. The large overlap we found between the GO terms associated with EGCG signatures from different cell types in men and mice indicates the conservation of these biological processes. The EGCG-upregulated signatures can be classified by GO terms related to metabolism (amine and phenol metabolism), ion transport, membranes and secretion. The EGCG-downregulated signatures can be classified by GO terms related to calcium signalling, neuronal and synaptic, inflammatory (response to wounding), connective tissue, ion transport and taxis and—concerning the kidneys—with microalbumin in urine and renal cancer. Calcium signalling and the influx and overload of cytosol and mitochondria, have been implicated in oxidative stress and the subsequent development of diseases [89,90]. Among the genes associated with the “response to wounding”, we found neuregulin-1 (NRG1) overlapping with downregulated genes in PAN-treated kidney organoids. NRG1 has been reported to attenuate the development of nephropathy in a diabetes I mouse model [91].
As significantly enriched transcription factors E2F1, NFKB1 and JUN were identified and furthermore found at the core of a gene regulatory network controlling inflammatory processes, E2F1 and NFKB1 have been shown to mediate inflammatory processes [92,93] and are connected to JUN/JNK [94,95]. That the same mechanisms are active in renal cells was shown by Liang et al., who reported that EGCG reduces kidney cell inflammation through the NF-κB pathway in HEK cells [96].
We compared this signature to transcriptomes of kidney cells treated with other natural antioxidants. The comparison to HEK cells treated with the marine natural product-derived compound VT01454 identified only GRIK2 and SOX15. The comparison to mouse kidneys treated with the mushroom Ganoderma lucidum resulted in a bigger overlap of six genes downregulated by EGCG (SMCO3, SLC12A1, FHAD1, FAT3, LOX and DOK7) and two genes upregulated by EGCG (TTC9 and PM20D1). Genes downregulated by EGCG, especially SLC12A1 or LOX, can be associated with several processes related to renal disease, most significantly “abnormal renal water reabsorption”.
We compared the Ganoderma lucidum-associated gene signatures to datasets of kidney disease models: (i) kidney organoids treated with the nephrotoxin PAN [82] and (ii) urine-derived podocytes treated with ANG II [83]. The largest intersection subset found in the 100 genes upregulated in GL and downregulated in PAN points at the therapeutic effects of GL on damage induced by PAN as a model for kidney disease. A GO term associated with these 100 genes was VEGF signalling, which we also found to be a major player in CKD in previous publications [4,97,98]. Also, in comparison to ANGII treatment, the largest subset was found in the 90 genes upregulated in GL and downregulated in ANGII. These 90 genes are associated with GO terms related to epithelial cell development, inflammatory processes such as interferon-alpha and leukaemia inhibitory factor responses and ERBB and BDNF signalling, while the 76 genes upregulated in both GL and in ANGII are associated with ADP synthesis and metabolism, several other metabolic processes and the positive regulation of acute inflammatory processes. Following up on these inflammatory processes in more detail, we identified the genes C3 and OSMR upregulated in the GL and ANGII stimulations, associated with the “positive regulation of the acute inflammatory response” (p = 0.0044), and the genes DDR1, DSP, HMOX1, ID3, PLAT, PLEC, USF1, ZFP36 and ZFP36L2 upregulated in the GL treatment and downregulated in the ANGII treatment, associated with the “response to wounding” (p = 0.0207).
7. Conclusions
Plants with antioxidative and anti-inflammatory properties are abundant in nature and can be found globally. The studies so far have shown that plant-based extracts contain various phytoconstituents that give them their respective properties. Many cases of AKI are too serious to be able to manage them directly using natural products. However, they may be useful in reducing the risk of AKI and in the exploration of disease-associated mechanisms and drug targets for AKI and CKD.
Our meta-analysis of transcriptome data and comparisons to in vitro kidney disease models has demonstrated that EGCG and GL have great potential for therapeutic effects mitigating inflammatory processes in kidney diseases. This has partially already been shown for EGCG in other in vitro models, e.g., by Kanlaya et al. [88]; this, together with our findings, suggests that further studies evaluating the therapeutic effects of GL and EGCG in models of human kidney disease might be fruitful. There are numerous unexploited plant species and herbs that might have antioxidant and anti-inflammatory activities; hence, we recommend the exploitation, and detailed studies on the mechanisms and targets, of these natural antioxidants and anti-inflammatories and the potential use of these drug targets for the management of kidney-associated diseases such as AKI and CKD.
Author Contributions
Conceptualisation: J.A. and A.K.A.G.; Methodology, W.W., A.K.A.G. and J.A.; Software, W.W.; Validation, W.W.; Formal Analysis, W.W.; Investigation, A.K.A.G.; Data Curation, W.W.; Writing—Original Draft Preparation, W.W. and A.K.A.G.; Writing—Review and Editing, J.A.; Visualisation, W.W.; Supervision, J.A. and Project Administration, J.A. All authors have read and agreed to the published version of the manuscript.
Funding
James Adjaye acknowledges support from the medical faculty of Heinrich-Heine University, Duesseldorf. This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chawla, L.S.; Eggers, P.W.; Star, R.A.; Kimmel, P.L. Acute Kidney Injury and Chronic Kidney Disease as Interconnected Syndromes. N. Engl. J. Med. 2014, 371, 58–66. [Google Scholar] [CrossRef]
- Levey, A.S.; Coresh, J. Chronic Kidney Disease. Lancet 2012, 379, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Coresh, J.; Balk, E.; Kausz, A.T.; Levin, A.; Steffes, M.W.; Hogg, R.J.; Perrone, R.D.; Lau, J.; Eknoyan, G.; et al. National Kidney Foundation Practice Guidelines for Chronic Kidney Disease: Evaluation, Classification, and Stratification. Ann. Intern. Med. 2003, 139, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Wruck, W.; Boima, V.; Erichsen, L.; Thimm, C.; Koranteng, T.; Kwakyi, E.; Antwi, S.; Adu, D.; Adjaye, J. Urine-Based Detection of Biomarkers Indicative of Chronic Kidney Disease in a Patient Cohort from Ghana. J. Pers. Med. 2022, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, G.; Bazzi, C. Pathophysiology of Proteinuria. Kidney Int. 2003, 63, 809–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schena, F.P.; Gesualdo, L. Pathogenetic Mechanisms of Diabetic Nephropathy. J. Am. Soc. Nephrol. 2005, 16, S30–S33. [Google Scholar] [CrossRef] [Green Version]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Al-Jaghbeer, M.; Dealmeida, D.; Bilderback, A.; Ambrosino, R.; Kellum, J.A. Clinical Decision Support for In-Hospital AKI. J. Am. Soc. Nephrol. JASN 2018, 29, 654–660. [Google Scholar] [CrossRef] [Green Version]
- Ronco, C.; Bellomo, R.; Kellum, J.A. Acute Kidney Injury. Lancet 2019, 394, 1949–1964. [Google Scholar] [CrossRef]
- Makris, K.; Spanou, L. Acute Kidney Injury: Definition, Pathophysiology and Clinical Phenotypes. Clin. Biochem. Rev. 2016, 37, 85–98. [Google Scholar]
- Erichsen, L.; Thimm, C.; Wruck, W.; Kaierle, D.; Schless, M.; Huthmann, L.; Dimski, T.; Kindgen-Milles, D.; Brandenburger, T.; Adjaye, J. Secreted Cytokines within the Urine of AKI Patients Modulate TP53 and SIRT1 Levels in a Human Podocyte Cell Model. Int. J. Mol. Sci. 2023, 24, 8228. [Google Scholar] [CrossRef]
- Wu, L.; Gokden, N.; Mayeux, P.R. Evidence for the Role of Reactive Nitrogen Species in Polymicrobial Sepsis-Induced Renal Peritubular Capillary Dysfunction and Tubular Injury. J. Am. Soc. Nephrol. 2007, 18, 1807–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonventre, J.V.; Zuk, A. Ischemic Acute Renal Failure: An Inflammatory Disease? Kidney Int. 2004, 66, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Nath, K.A.; Norby, S.M. Reactive Oxygen Species and Acute Renal Failure. Am. J. Med. 2000, 109, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, J.; Chen, X.; Xie, J.; Yang, Q.; Wang, J.; Deng, X. Senkyunolide I Alleviates Renal Ischemia-Reperfusion Injury by Inhibiting Oxidative Stress, Endoplasmic Reticulum Stress and Apoptosis. Int. Immunopharmacol. 2022, 102, 108393. [Google Scholar] [CrossRef]
- Ishimoto, Y.; Tanaka, T.; Yoshida, Y.; Inagi, R. Physiological and Pathophysiological Role of Reactive Oxygen Species and Reactive Nitrogen Species in the Kidney. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1097–1105. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Lei, C.-T.; Zhang, C. Interleukin-6 Signaling Pathway and Its Role in Kidney Disease: An Update. Front. Immunol. 2017, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- Magno, A.; Herat, L.; Carnagarin, R.; Schlaich, M.; Matthews, V. Current Knowledge of IL-6 Cytokine Family Members in Acute and Chronic Kidney Disease. Biomedicines 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Bonventre, J.V.; Yang, L. Cellular Pathophysiology of Ischemic Acute Kidney Injury. J. Clin. Investig. 2011, 121, 4210–4221. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chang, A.; Hack, B.K.; Eadon, M.T.; Alper, S.L.; Cunningham, P.N. TNF-Mediated Damage to Glomerular Endothelium Is an Important Determinant of Acute Kidney Injury in Sepsis. Kidney Int. 2014, 85, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, M.A.; Vaidya, V.S.; Bonventre, J.V. Biomarkers of Nephrotoxic Acute Kidney Injury. Toxicology 2008, 245, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinke, H.; Masuda, S.; Togashi, Y.; Ikemi, Y.; Ozawa, A.; Sato, T.; Kim, Y.H.; Mishima, M.; Ichimura, T.; Bonventre, J.V.; et al. Urinary Kidney Injury Molecule-1 and Monocyte Chemotactic Protein-1 Are Noninvasive Biomarkers of Cisplatin-Induced Nephrotoxicity in Lung Cancer Patients. Cancer Chemother. Pharmacol. 2015, 76, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Vaidya, V.S.; Brown, R.P.; Zhang, J.; Rosenzweig, B.A.; Thompson, K.L.; Miller, T.J.; Bonventre, J.V.; Goering, P.L. Comparison of Kidney Injury Molecule-1 and Other Nephrotoxicity Biomarkers in Urine and Kidney Following Acute Exposure to Gentamicin, Mercury, and Chromium. Toxicol. Sci. 2008, 101, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Choucry, M.A.; Khalil, M.N.A.; El Awdan, S.A. Protective Action of Crateva Nurvala Buch. Ham Extracts against Renal Ischaemia Reperfusion Injury in Rats via Antioxidant and Anti-Inflammatory Activities. J. Ethnopharmacol. 2018, 214, 47–57. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, B.; Li, Y.; Zhang, H. Protective Effect of Luteolin Against Renal Ischemia/Reperfusion Injury via Modulation of Pro-Inflammatory Cytokines, Oxidative Stress and Apoptosis for Possible Benefit in Kidney Transplant. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2017, 23, 5720–5727. [Google Scholar] [CrossRef] [Green Version]
- Punuru, P.; Sujatha, D.; Kumari, B.P.; Charisma, V.V.L. Evaluation of Aqueous Extract of Murraya Koenigii in Unilateral Renal Ischemia Reperfusion Injury in Rats. Indian J. Pharmacol. 2014, 46, 171–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, C.; Yang, J.; Yang, H.; Li, X.; Wang, G. Attenuation of Renal Ischemia/Reperfusion Injury by Oleanolic Acid Preconditioning via Its Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Activities. Mol. Med. Rep. 2016, 13, 4697–4704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Liu, H.; Wang, J. Polydatin Ameliorates Renal Ischemia/Reperfusion Injury by Decreasing Apoptosis and Oxidative Stress through Activating Sonic Hedgehog Signaling Pathway. Food Chem. Toxicol. 2016, 96, 215–225. [Google Scholar] [CrossRef]
- Ren, Y.; Frank, T.; Meyer, G.; Lei, J.; Grebenc, J.R.; Slaughter, R.; Gao, Y.G.; Kinghorn, A.D. Potential Benefits of Black Chokeberry (Aronia melanocarpa) Fruits and Their Constituents in Improving Human Health. Mol. Basel Switz. 2022, 27, 7823. [Google Scholar] [CrossRef]
- Kang, H.G.; Lee, H.K.; Cho, K.B.; Park, S.I. A Review of Natural Products for Prevention of Acute Kidney Injury. Medicina (Mex.) 2021, 57, 1266. [Google Scholar] [CrossRef]
- Mestry, S.N.; Gawali, N.B.; Pai, S.A.; Gursahani, M.S.; Dhodi, J.B.; Munshi, R.; Juvekar, A.R. Punica Granatum Improves Renal Function in Gentamicin-Induced Nephropathy in Rats via Attenuation of Oxidative Stress. J. Ayurveda Integr. Med. 2020, 11, 16–23. [Google Scholar] [CrossRef]
- El Bohi, K.M.; Abdel-Motal, S.M.; Khalil, S.R.; Abd-Elaal, M.M.; Metwally, M.M.M.; ELhady, W.M. The Efficiency of Pomegranate (Punica granatum) Peel Ethanolic Extract in Attenuating the Vancomycin-Triggered Liver and Kidney Tissues Injury in Rats. Environ. Sci. Pollut. Res. 2021, 28, 7134–7150. [Google Scholar] [CrossRef] [PubMed]
- Nerdy, N.; Ritarwan, K. Hepatoprotective Activity and Nephroprotective Activity of Peel Extract from Three Varieties of the Passion Fruit (Passiflora sp.) in the Albino Rat. Open Access Maced. J. Med. Sci. 2019, 7, 536–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Yousef, H.M.; Alqahtani, A.S.; Ghani, A.S.A.; El-Toumy, S.A.; El-Dougdoug, W.I.A.; Hassan, W.H.B.; Hassan, H.M. Nephroprotective, Cytotoxic and Antioxidant Activities of Euphorbia Paralias. Saudi J. Biol. Sci. 2021, 28, 785–792. [Google Scholar] [CrossRef]
- Heidarian, E.; Jafari-Dehkordi, E.; Valipour, P.; Ghatreh-Samani, K.; Ashrafi-Eshkaftaki, L. Nephroprotective and Anti-Inflammatory Effects of Pistacia atlantica Leaf Hydroethanolic Extract Against Gentamicin-Induced Nephrotoxicity in Rats. J. Diet. Suppl. 2017, 14, 489–502. [Google Scholar] [CrossRef]
- Chinnappan, S.M.; George, A.; Thaggikuppe, P.; Choudhary, Y.; Choudhary, V.K.; Ramani, Y.; Dewangan, R. Nephroprotective Effect of Herbal Extract Eurycoma longifolia on Paracetamol-Induced Nephrotoxicity in Rats. Evid. Based Complement. Alternat. Med. 2019, 2019, 4916519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyasor, G.N.; Onajobi, F.D.; Osilesi, O.; Adebawo, O. Proximate Composition, Mineral Content and in Vitro Antioxidant Activity of Leaf and Stem of Costus afer (Ginger Lily). J. Intercult. Ethnopharmacol. 2014, 3, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Ezejiofor, A.N.; Udowelle, N.A.; Orisakwe, O.E. Nephroprotective and Antioxidant Effect of Aqueous Leaf Extract of Costus Afer Ker Gawl on Cyclosporin-a (Csa) Induced Nephrotoxicity. Clin. Phytoscience 2017, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Anyanwu, B.O.; Orish, C.N.; Ezejiofor, A.N.; Nwaogazie, I.L.; Orisakwe, O.E. Neuroprotective Effect of Costus Afer on Low Dose Heavy Metal Mixture (Lead, Cadmium and Mercury) Induced Neurotoxicity via Antioxidant, Anti-Inflammatory Activities. Toxicol. Rep. 2020, 7, 1032–1038. [Google Scholar] [CrossRef]
- Genfi, A.K.A.; Larbie, C.; Emikpe, B.O.; Oyagbemi, A.A.; Firempong, C.K.; Adjei, C.O. Modulation of Oxidative Stress and Inflammatory Cytokines as Therapeutic Mechanisms of Ocimum Americanum L Extract in Carbon Tetrachloride and Acetaminophen-Induced Toxicity in Rats. J. Evid.-Based Integr. Med. 2020, 25, 2515690X2093800. [Google Scholar] [CrossRef]
- Nyarko, A.K.; Asare-Anane, H.; Ofosuhene, M.; Addy, M.E. Extract of Ocimum Canum Lowers Blood Glucose and Facilitates Insulin Release by Isolated Pancreatic β-Islet Cells. Phytomedicine 2002, 9, 346–351. [Google Scholar] [CrossRef]
- Ranfaing, J.; Dunyach-Remy, C.; Louis, L.; Lavigne, J.-P.; Sotto, A. Propolis Potentiates the Effect of Cranberry (Vaccinium macrocarpon) against the Virulence of Uropathogenic Escherichia Coli. Sci. Rep. 2018, 8, 10706. [Google Scholar] [CrossRef] [Green Version]
- Amin, R.; Thalluri, C.; Docea, A.O.; Sharifi-Rad, J.; Calina, D. Therapeutic Potential of Cranberry for Kidney Health and Diseases. eFood 2022, 3, e33. [Google Scholar] [CrossRef]
- Csikós, E.; Horváth, A.; Ács, K.; Papp, N.; Balázs, V.L.; Dolenc, M.S.; Kenda, M.; Kočevar Glavač, N.; Nagy, M.; Protti, M.; et al. Treatment of Benign Prostatic Hyperplasia by Natural Drugs. Molecules 2021, 26, 7141. [Google Scholar] [CrossRef]
- Carneiro, D.M.; Freire, R.C.; Honório, T.C.D.D.; Zoghaib, I.; Cardoso, F.F.D.S.E.S.; Tresvenzol, L.M.F.; De Paula, J.R.; Sousa, A.L.L.; Jardim, P.C.B.V.; Cunha, L.C.D. Randomized, Double-Blind Clinical Trial to Assess the Acute Diuretic Effect of Equisetum arvense (Field Horsetail) in Healthy Volunteers. Evid. Based Complement. Alternat. Med. 2014, 2014, 760683. [Google Scholar] [CrossRef] [Green Version]
- Pallag, A.; Filip, G.A.; Olteanu, D.; Clichici, S.; Baldea, I.; Jurca, T.; Micle, O.; Vicaş, L.; Marian, E.; Soriţău, O.; et al. Equisetum arvense L. Extract Induces Antibacterial Activity and Modulates Oxidative Stress, Inflammation, and Apoptosis in Endothelial Vascular Cells Exposed to Hyperosmotic Stress. Oxid. Med. Cell. Longev. 2018, 2018, 3060525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabi, T.D.; Brooks, N.L.; Oguntibeju, O.O. Leaf Extracts of Anchomanes Difformis Ameliorated Kidney and Pancreatic Damage in Type 2 Diabetes. Plants 2021, 10, 300. [Google Scholar] [CrossRef]
- Rodríguez-Fierros, F.L.; Guarner-Lans, V.; Soto, M.E.; Manzano-Pech, L.; Díaz-Díaz, E.; Soria-Castro, E.; Rubio-Ruiz, M.E.; Jiménez-Trejo, F.; Pérez-Torres, I. Modulation of Renal Function in a Metabolic Syndrome Rat Model by Antioxidants in Hibiscus sabdariffa L. Molecules 2021, 26, 2074. [Google Scholar] [CrossRef]
- Benzer, F.; Kandemir, F.M.; Kucukler, S.; Comaklı, S.; Caglayan, C. Chemoprotective Effects of Curcumin on Doxorubicin-Induced Nephrotoxicity in Wistar Rats: By Modulating Inflammatory Cytokines, Apoptosis, Oxidative Stress and Oxidative DNA Damage. Arch. Physiol. Biochem. 2018, 124, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Akhondzadeh, S. Melissa Officinalis Extract in the Treatment of Patients with Mild to Moderate Alzheimer’s Disease: A Double Blind, Randomised, Placebo Controlled Trial. J. Neurol. Neurosurg. Psychiatry 2003, 74, 863–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarei, A.; Changizi-Ashtiyani, S.; Taheri, S.; Hosseini, N. A Brief Overview of the Effects of Melissa Officinalis L. Extract on the Function of Various Body Organs. Zahedan J. Res. Med. Sci. 2015, 17. [Google Scholar] [CrossRef] [Green Version]
- Haji, S.A.; Movahed, A. Update on Digoxin Therapy in Congestive Heart Failure. Am. Fam. Physician 2000, 62, 409–416. [Google Scholar]
- Sesso, H.D.; Liu, S.; Gaziano, J.M.; Buring, J.E. Dietary Lycopene, Tomato-Based Food Products and Cardiovascular Disease in Women. J. Nutr. 2003, 133, 2336–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viuda-Martos, M.; Sanchez-Zapata, E.; Sayas-Barberá, E.; Sendra, E.; Pérez-Álvarez, J.A.; Fernández-López, J. Tomato and Tomato Byproducts. Human Health Benefits of Lycopene and Its Application to Meat Products: A Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1032–1049. [Google Scholar] [CrossRef]
- Chaudhuri, K.N.; Ghosh, B.; Tepfer, D.; Jha, S. Genetic Transformation of Tylophora Indica with Agrobacterium rhizogenes A4: Growth and Tylophorine Productivity in Different Transformed Root Clones. Plant Cell Rep. 2005, 24, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Harrath, A.H. Phytoestrogens and Their Effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Bhathena, S.J. Dietary Phytoestrogens: A Possible Role in Renal Disease Protection. Am. J. Kidney Dis. 2001, 37, 1056–1068. [Google Scholar] [CrossRef]
- Ríos-Silva, M.; Santos-Álvarez, R.; Trujillo, X.; Cárdenas-María, R.; López-Zamudio, M.; Bricio-Barrios, J.; Leal, C.; Saavedra-Molina, A.; Huerta-Trujillo, M.; Espinoza-Mejía, K.; et al. Effects of Chronic Administration of Capsaicin on Biomarkers of Kidney Injury in Male Wistar Rats with Experimental Diabetes. Molecules 2018, 24, 36. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and Clinical Uses of Capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef]
- Hewlings, S.; Kalman, D. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkara, P.J.; Sabina, E.P. Pre-Treatment with Beta Carotene Gives Protection Against Nephrotoxicity Induced by Bromobenzene via Modulation of Antioxidant System, Pro-Inflammatory Cytokines and Pro-Apoptotic Factors. Appl. Biochem. Biotechnol. 2020, 190, 616–633. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A.; Stahl, W.; Rock, C.L. Beta Carotene: From Biochemistry to Clinical Trials. Nutr. Rev. 2009, 58, 39–53. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea Catechins and Polyphenols: Health Effects, Metabolism, and Antioxidant Functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Wongmekiat, O.; Peerapanyasut, W.; Kobroob, A. Catechin Supplementation Prevents Kidney Damage in Rats Repeatedly Exposed to Cadmium through Mitochondrial Protection. Naunyn. Schmiedebergs Arch. Pharmacol. 2018, 391, 385–394. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential of Resveratrol: The In Vivo Evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Schempp, C.M.; Pelz, K.; Wittmer, A.; Schöpf, E.; Simon, J.C. Antibacterial Activity of Hyperforin from St John’s Wort, against Multiresistant Staphylococcus Aureus and Gram-Positive Bacteria. Lancet 1999, 353, 2129. [Google Scholar] [CrossRef]
- Prassas, I.; Diamandis, E.P. Novel Therapeutic Applications of Cardiac Glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef]
- Harvey, A. Natural Products in Drug Discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Thomford, N.; Senthebane, D.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Alice, C.B.; Vargas, V.M.F.; Silva, G.A.A.B.; De Siqueira, N.C.S.; Schapoval, E.E.S.; Gleye, J.; Henriques, J.A.P.; Henriques, A.T. Screening of Plants Used in South Brazilian Folk Medicine. J. Ethnopharmacol. 1991, 35, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, J.O.; Krettli, A.U. Potential Antimalarials from Nigerian Plants: A Review. J. Ethnopharmacol. 2011, 133, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ud-Din, S.; Wilgus, T.A.; McGeorge, D.D.; Bayat, A. Pre-Emptive Priming of Human Skin Improves Cutaneous Scarring and Is Superior to Immediate and Delayed Topical Anti-Scarring Treatment Post-Wounding: A Double-Blind Randomised Placebo-Controlled Clinical Trial. Pharmaceutics 2021, 13, 510. [Google Scholar] [CrossRef] [PubMed]
- Kehrmann, J.; Tatura, R.; Zeschnigk, M.; Probst-Kepper, M.; Geffers, R.; Steinmann, J.; Buer, J. Impact of 5-Aza-2′-Deoxycytidine and Epigallocatechin-3-Gallate for Induction of Human Regulatory T Cells. Immunology 2014, 142, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.P.G.; Cooney, J.M.; Dommels, Y.E.M.; Nones, K.; Brewster, D.T.; Park, Z.; Butts, C.A.; McNabb, W.C.; Laing, W.A.; Roy, N.C. Modulation of Colonic Inflammation in Mdr1a(−/−) Mice by Green Tea Polyphenols and Their Effects on the Colon Transcriptome and Proteome. J. Nutr. Biochem. 2013, 24, 1678–1690. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 3. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Falcon, S.; Gentleman, R. Using GOstats to Test Gene Lists for GO Term Association. Bioinforma. Oxf. Engl. 2007, 23, 257–258. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Use R!; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING V10: Protein–Protein Interaction Networks, Integrated over the Tree of Life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.; Wruck, W.; Erichsen, L.; Graffmann, N.; Adjaye, J. The Nephrotoxin Puromycin Aminonucleoside Induces Injury in Kidney Organoids Differentiated from Induced Pluripotent Stem Cells. Cells 2022, 11, 635. [Google Scholar] [CrossRef] [PubMed]
- Erichsen, L.; Thimm, C.; Bohndorf, M.; Rahman, M.S.; Wruck, W.; Adjaye, J. Activation of the Renin–Angiotensin System Disrupts the Cytoskeletal Architecture of Human Urine-Derived Podocytes. Cells 2022, 11, 1095. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-L.; Fu, V.; Liu, G.; Tang, T.; Konradi, A.W.; Peng, X.; Kemper, E.; Cravatt, B.F.; Franklin, J.M.; Wu, Z.; et al. Hippo Pathway Regulation by Phosphatidylinositol Transfer Protein and Phosphoinositides. Nat. Chem. Biol. 2022, 18, 1076–1086. [Google Scholar] [CrossRef]
- Romero-Córdoba, S.L.; Salido-Guadarrama, I.; Meneses, M.E.; Cosentino, G.; Iorio, M.V.; Tagliabue, E.; Torres, N.; Sánchez-Tapia, M.; Bonilla, M.; Castillo, I.; et al. Mexican Ganoderma Lucidum Extracts Decrease Lipogenesis Modulating Transcriptional Metabolic Networks and Gut Microbiota in C57BL/6 Mice Fed with a High-Cholesterol Diet. Nutrients 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Kürbitz, C.; Heise, D.; Redmer, T.; Goumas, F.; Arlt, A.; Lemke, J.; Rimbach, G.; Kalthoff, H.; Trauzold, A. Epicatechin Gallate and Catechin Gallate Are Superior to Epigallocatechin Gallate in Growth Suppression and Anti-Inflammatory Activities in Pancreatic Tumor Cells. Cancer Sci. 2011, 102, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Yang, C.S. Mechanisms of Cancer Prevention by Tea Constituents. J. Nutr. 2003, 133, 3262S–3267S. [Google Scholar] [CrossRef] [Green Version]
- Kanlaya, R.; Khamchun, S.; Kapincharanon, C.; Thongboonkerd, V. Protective Effect of Epigallocatechin-3-Gallate (EGCG) via Nrf2 Pathway against Oxalate-Induced Epithelial Mesenchymal Transition (EMT) of Renal Tubular Cells. Sci. Rep. 2016, 6, 30233. [Google Scholar] [CrossRef]
- Ermak, G.; Davies, K.J.A. Calcium and Oxidative Stress: From Cell Signaling to Cell Death. Mol. Immunol. 2002, 38, 713–721. [Google Scholar] [CrossRef]
- Peng, T.-I.; Jou, M.-J. Oxidative Stress Caused by Mitochondrial Calcium Overload. Ann. N. Y. Acad. Sci. 2010, 1201, 183–188. [Google Scholar] [CrossRef]
- Vandekerckhove, L.; Vermeulen, Z.; Liu, Z.Z.; Boimvaser, S.; Patzak, A.; Segers, V.F.M.; De Keulenaer, G.W. Neuregulin-1 Attenuates Development of Nephropathy in a Type 1 Diabetes Mouse Model with High Cardiovascular Risk. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E495–E504. [Google Scholar] [CrossRef] [Green Version]
- Denechaud, P.-D.; Fajas, L.; Giralt, A. E2F1, a Novel Regulator of Metabolism. Front. Endocrinol. 2017, 8, 311. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, R.; Zhou, J. E2F1 and NF-ΚB: Key Mediators of Inflammation-Associated Cancers and Potential Therapeutic Targets. Curr. Cancer Drug Targets 2016, 16, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. Regulation of NF-ΚB Activation by MAP Kinase Cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Liang, Y.-J.; Jian, J.-H.; Liu, Y.-C.; Juang, S.-J.; Shyu, K.-G.; Lai, L.-P.; Wang, B.-W.; Leu, J.-G. Advanced Glycation End Products-Induced Apoptosis Attenuated by PPARδ Activation and Epigallocatechin Gallate through NF-ΚB Pathway in Human Embryonic Kidney Cells and Human Mesangial Cells. Diabetes Metab. Res. Rev. 2010, 26, 406–416. [Google Scholar] [CrossRef]
- Boima, V.; Tannor, E.K.; Osafo, C.; Awuku, Y.A.; Mate-Kole, M.; Davids, M.R.; Adu, D. The Ghana Renal Registry—A First Annual Report. Afr. J. Nephrol. 2021, 24, 19–24. [Google Scholar] [CrossRef]
- Osafo, C.; Mate-Kole, M.; Affram, K.; Adu, D. Prevalence of Chronic Kidney Disease in Hypertensive Patients in Ghana. Ren. Fail. 2011, 33, 388–392. [Google Scholar] [CrossRef]
Figure 1.
Gene signature upregulated by EGCG. (A) Venn diagram comparison of genes upregulated in EGCG-treated human skin cells, human T cells and mouse colon cells. The orange circle marks the 11 genes upregulated in common in all 3 datasets, and the red ellipse marks the 23 (11 + 12) genes upregulated in common in the two human datasets. (B) Overrepresented GO terms in the common signatures of the 11 genes. (C) Overrepresented GO terms in the common human EGCG signatures of the 23 genes. The large overlap between both GO analysis results shows that most of these biological processes are conserved between men and mice. The results can be grouped into metabolic (amine and phenol metabolism), ion transport, membrane and secretory categories. (D) The EGCG gene signatures from the marked subsets of the Venn diagram.
Figure 1.
Gene signature upregulated by EGCG. (A) Venn diagram comparison of genes upregulated in EGCG-treated human skin cells, human T cells and mouse colon cells. The orange circle marks the 11 genes upregulated in common in all 3 datasets, and the red ellipse marks the 23 (11 + 12) genes upregulated in common in the two human datasets. (B) Overrepresented GO terms in the common signatures of the 11 genes. (C) Overrepresented GO terms in the common human EGCG signatures of the 23 genes. The large overlap between both GO analysis results shows that most of these biological processes are conserved between men and mice. The results can be grouped into metabolic (amine and phenol metabolism), ion transport, membrane and secretory categories. (D) The EGCG gene signatures from the marked subsets of the Venn diagram.
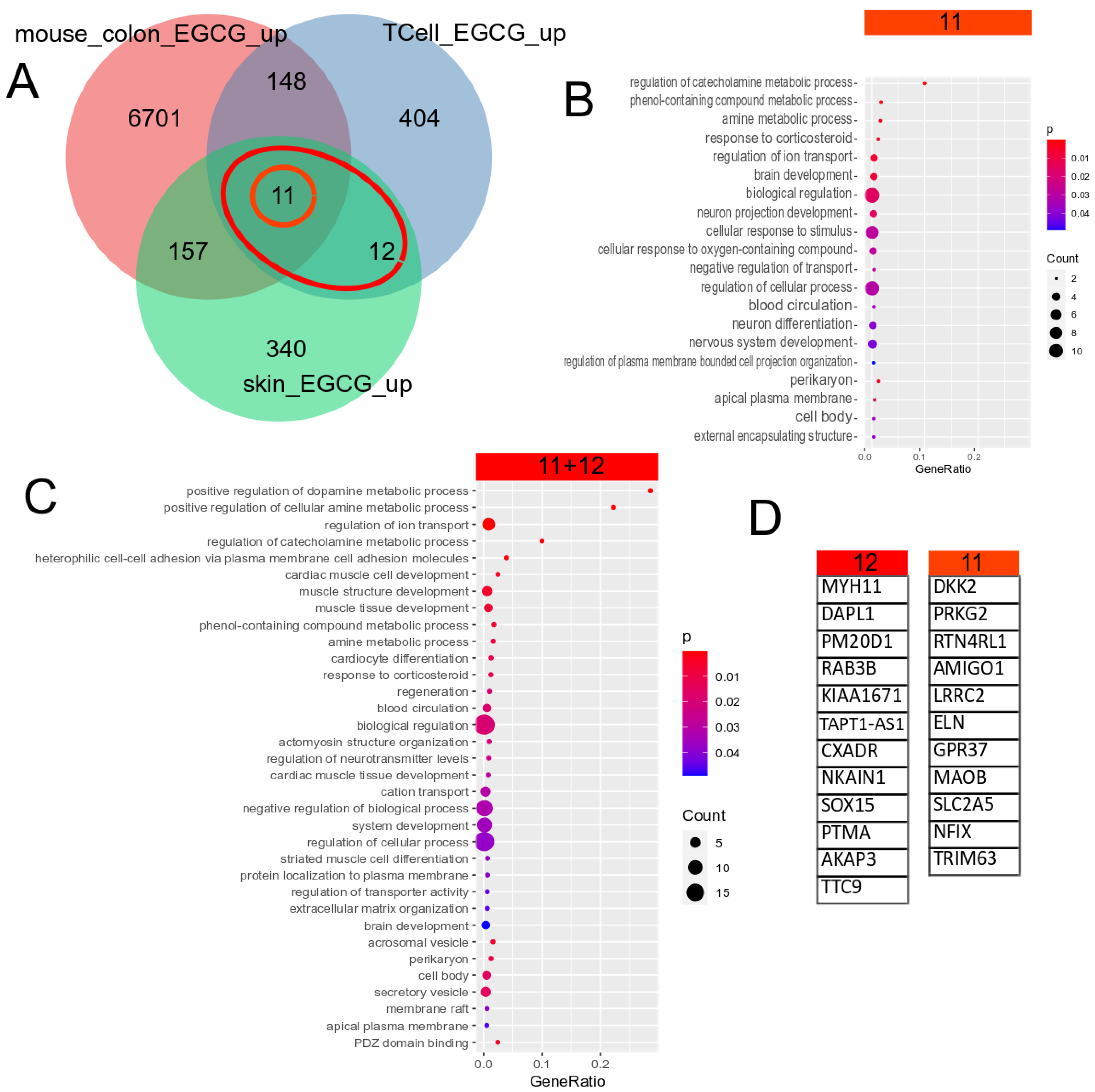
Figure 2.
Gene signature downregulated by EGCG. (A) Venn diagram comparison of the genes downregulated in EGCG-treated human skin cells, human T cells and mouse colon cells. The orange circle marks the 13 genes downregulated in common in all 3 datasets, and the red ellipse marks the 50 (13 + 37) genes downregulated in common in the two human datasets. (B) Overrepresented GO terms in the common signatures of the 13 genes. (C) Overrepresented GO terms in the common human EGCG signatures of the 50 genes. The large overlap between both GO analysis results shows that most of these biological processes are conserved between men and mice. The results can be grouped into calcium signalling, neuronal and synaptic, inflammatory (response to wounding), connective tissue, ion transport and taxis categories. (D) The EGCG gene signatures from the marked subsets of the Venn diagram.
Figure 2.
Gene signature downregulated by EGCG. (A) Venn diagram comparison of the genes downregulated in EGCG-treated human skin cells, human T cells and mouse colon cells. The orange circle marks the 13 genes downregulated in common in all 3 datasets, and the red ellipse marks the 50 (13 + 37) genes downregulated in common in the two human datasets. (B) Overrepresented GO terms in the common signatures of the 13 genes. (C) Overrepresented GO terms in the common human EGCG signatures of the 50 genes. The large overlap between both GO analysis results shows that most of these biological processes are conserved between men and mice. The results can be grouped into calcium signalling, neuronal and synaptic, inflammatory (response to wounding), connective tissue, ion transport and taxis categories. (D) The EGCG gene signatures from the marked subsets of the Venn diagram.
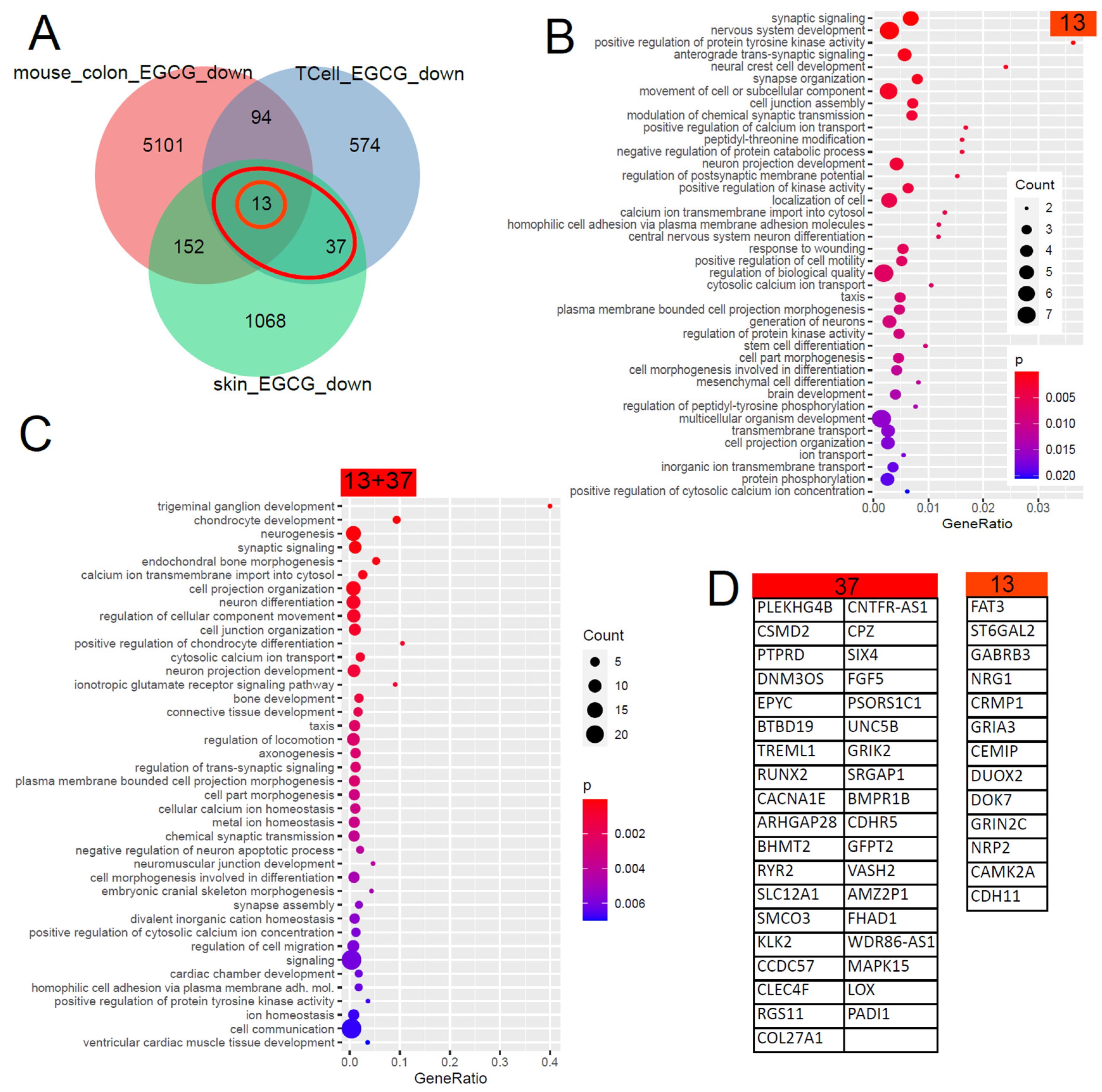
Figure 3.
EGCG-associated gene signatures may have an impact on the kidneys and are associated with inflammatory processes regulated by the transcription factors NFKB1, JUN and E2F1. (A) The enrichment analysis results of the EGCG-downregulated 50 (13 + 37) genes in the UK_Biobank_GWAS_v1 dataset collection and (B) the “Jensen Diseases” dataset collection. (C) Most EGCG-downregulated genes are regulated by transcription factor E2F1. (D) Most EGCG-upregulated genes are regulated by transcription factors JUN and NFKB1. (E) Protein interaction network of E2F1, NFKB1 and JUN expanded by five interacting proteins made via STRING-DB. (F) Top 20 Reactome pathways found by STRING-SB in the network of (E) reveal inflammatory processes. (G) Top 20 WikiPathway pathways found by STRING-SB in the network of (E) neuroinflammation and other inflammatory processes.
Figure 3.
EGCG-associated gene signatures may have an impact on the kidneys and are associated with inflammatory processes regulated by the transcription factors NFKB1, JUN and E2F1. (A) The enrichment analysis results of the EGCG-downregulated 50 (13 + 37) genes in the UK_Biobank_GWAS_v1 dataset collection and (B) the “Jensen Diseases” dataset collection. (C) Most EGCG-downregulated genes are regulated by transcription factor E2F1. (D) Most EGCG-upregulated genes are regulated by transcription factors JUN and NFKB1. (E) Protein interaction network of E2F1, NFKB1 and JUN expanded by five interacting proteins made via STRING-DB. (F) Top 20 Reactome pathways found by STRING-SB in the network of (E) reveal inflammatory processes. (G) Top 20 WikiPathway pathways found by STRING-SB in the network of (E) neuroinflammation and other inflammatory processes.
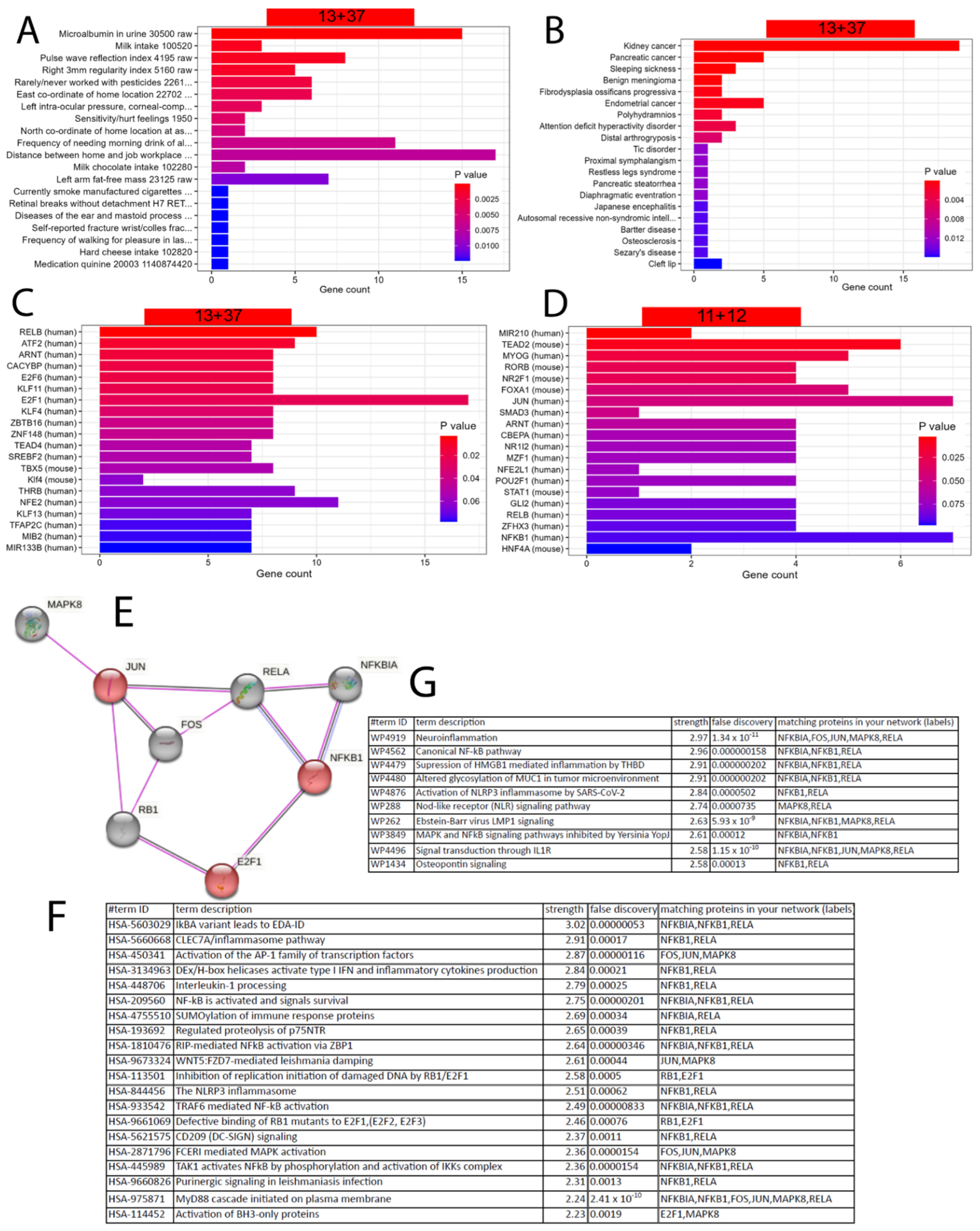
Figure 4.
EGCG-associated gene signatures are transferred to kidney datasets with other antioxidant compounds. (A) Venn diagram comparison of the human skin and T-cell up- and downregulated EGCG gene signatures to dataset GSE198890 of HEK cells treated with VT01454 shows only 1+1 genes overlapping. (B) In dataset GSE159656 of Galoderma lucidum-treated mouse kidney cells, there is more overlap of the indicated six genes in the downregulated EGCG signature and two genes in the upregulated EGCG signature. (C) The six EGCG-downregulated genes overlapping can be further characterised by the overrepresented renal impairment processes found by the R package EnrichR; however, they are only associated with single genes: SLC12A1 or LOX.
Figure 4.
EGCG-associated gene signatures are transferred to kidney datasets with other antioxidant compounds. (A) Venn diagram comparison of the human skin and T-cell up- and downregulated EGCG gene signatures to dataset GSE198890 of HEK cells treated with VT01454 shows only 1+1 genes overlapping. (B) In dataset GSE159656 of Galoderma lucidum-treated mouse kidney cells, there is more overlap of the indicated six genes in the downregulated EGCG signature and two genes in the upregulated EGCG signature. (C) The six EGCG-downregulated genes overlapping can be further characterised by the overrepresented renal impairment processes found by the R package EnrichR; however, they are only associated with single genes: SLC12A1 or LOX.

Figure 5.
Ganoderma lucidum-associated gene signatures are compared to kidney datasets with PAN and ANGII stimulation. (A) Venn diagram comparison of up- and downregulated genes in Ganoderma lucidum (GL)-treated mouse kidney and PAN-treated human kidney organoids. (B) Venn diagram comparison of up- and downregulated genes in GL-treated mouse kidney and ANGII-treated human urine-derived podocytes. The dot plots in (C–F) show the most significant overrepresented GO terms in the Venn diagram subsets with matching colours. (C) The most significant GO terms in the 100 genes upregulated in GL and downregulated in PAN include VEGF signalling. (D) The most significant GO terms in the 36 genes downregulated in GL and downregulated in PAN include sterol synthesis and blood circulation/pressure. (E) The most significant GO terms in the 90 genes upregulated in GL and downregulated in ANGII include epithelial cell development, inflammatory processes such as interferon-alpha and leukaemia inhibitory factor responses and ERBB and BDNF signalling. (F) The most significant GO terms in the 76 genes upregulated in GL and upregulated in ANGII include ADP synthesis and metabolism and several other metabolic processes, as well as the positive regulation of acute inflammatory processes.
Figure 5.
Ganoderma lucidum-associated gene signatures are compared to kidney datasets with PAN and ANGII stimulation. (A) Venn diagram comparison of up- and downregulated genes in Ganoderma lucidum (GL)-treated mouse kidney and PAN-treated human kidney organoids. (B) Venn diagram comparison of up- and downregulated genes in GL-treated mouse kidney and ANGII-treated human urine-derived podocytes. The dot plots in (C–F) show the most significant overrepresented GO terms in the Venn diagram subsets with matching colours. (C) The most significant GO terms in the 100 genes upregulated in GL and downregulated in PAN include VEGF signalling. (D) The most significant GO terms in the 36 genes downregulated in GL and downregulated in PAN include sterol synthesis and blood circulation/pressure. (E) The most significant GO terms in the 90 genes upregulated in GL and downregulated in ANGII include epithelial cell development, inflammatory processes such as interferon-alpha and leukaemia inhibitory factor responses and ERBB and BDNF signalling. (F) The most significant GO terms in the 76 genes upregulated in GL and upregulated in ANGII include ADP synthesis and metabolism and several other metabolic processes, as well as the positive regulation of acute inflammatory processes.
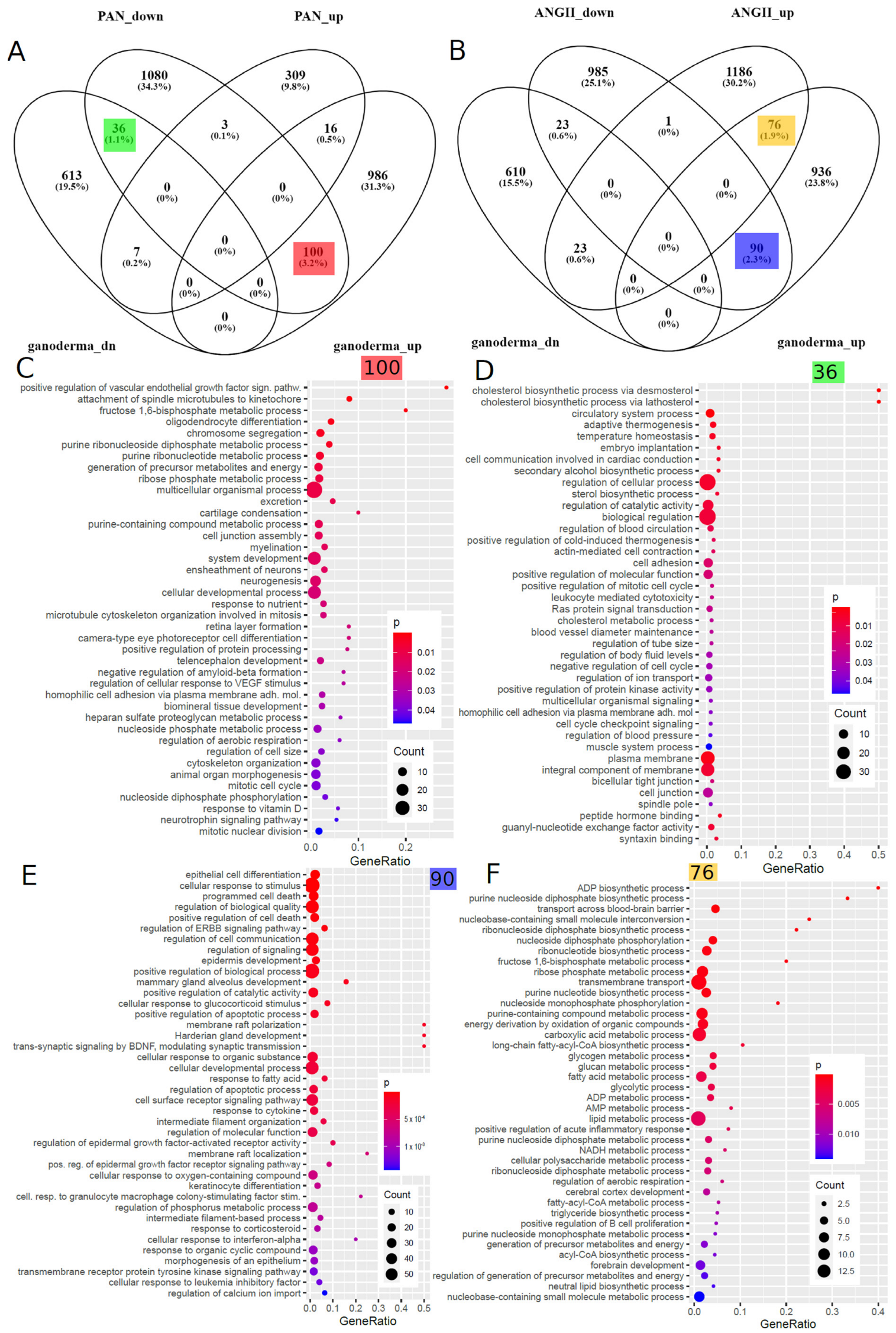
Table 2.
Inflammatory and wounding-related GO terms overrepresented in the genes overlapping between the GL and ANGII signatures.
Table 2.
Inflammatory and wounding-related GO terms overrepresented in the genes overlapping between the GL and ANGII signatures.
| Term | Ganoderma Lucidum | ANGII | p-Value | Odds Ratio | Genes |
|---|---|---|---|---|---|
| positive regulation of acute inflammatory response | up | up | 0.0043 | 22.34 | C3, OSMR |
| response to wounding | up | down | 0.0207 | 5.41 | DDR1, DSP, HMOX1, ID, PLAT, PLEC, USF1, ZFP36, ZFP36L2 |
| wound healing | up | down | 0.0473 | 2.65 | DDR1, DSP, HMOX1, PLAT, PLEC, USF1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wruck, W.; Genfi, A.K.A.; Adjaye, J. Natural Products in Renal-Associated Drug Discovery. Antioxidants 2023, 12, 1599. https://doi.org/10.3390/antiox12081599
AMA Style
Wruck W, Genfi AKA, Adjaye J. Natural Products in Renal-Associated Drug Discovery. Antioxidants. 2023; 12(8):1599. https://doi.org/10.3390/antiox12081599
Chicago/Turabian StyleWruck, Wasco, Afua Kobi Ampem Genfi, and James Adjaye. 2023. "Natural Products in Renal-Associated Drug Discovery" Antioxidants 12, no. 8: 1599. https://doi.org/10.3390/antiox12081599
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.