
Se Alleviated Pb-Caused Neurotoxicity in Chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO Pathway, Selenoprotein Suppression, Oxidative Stress, and Inflammatory Injury

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Sample Collection
2.3. The Observation of Microstructure
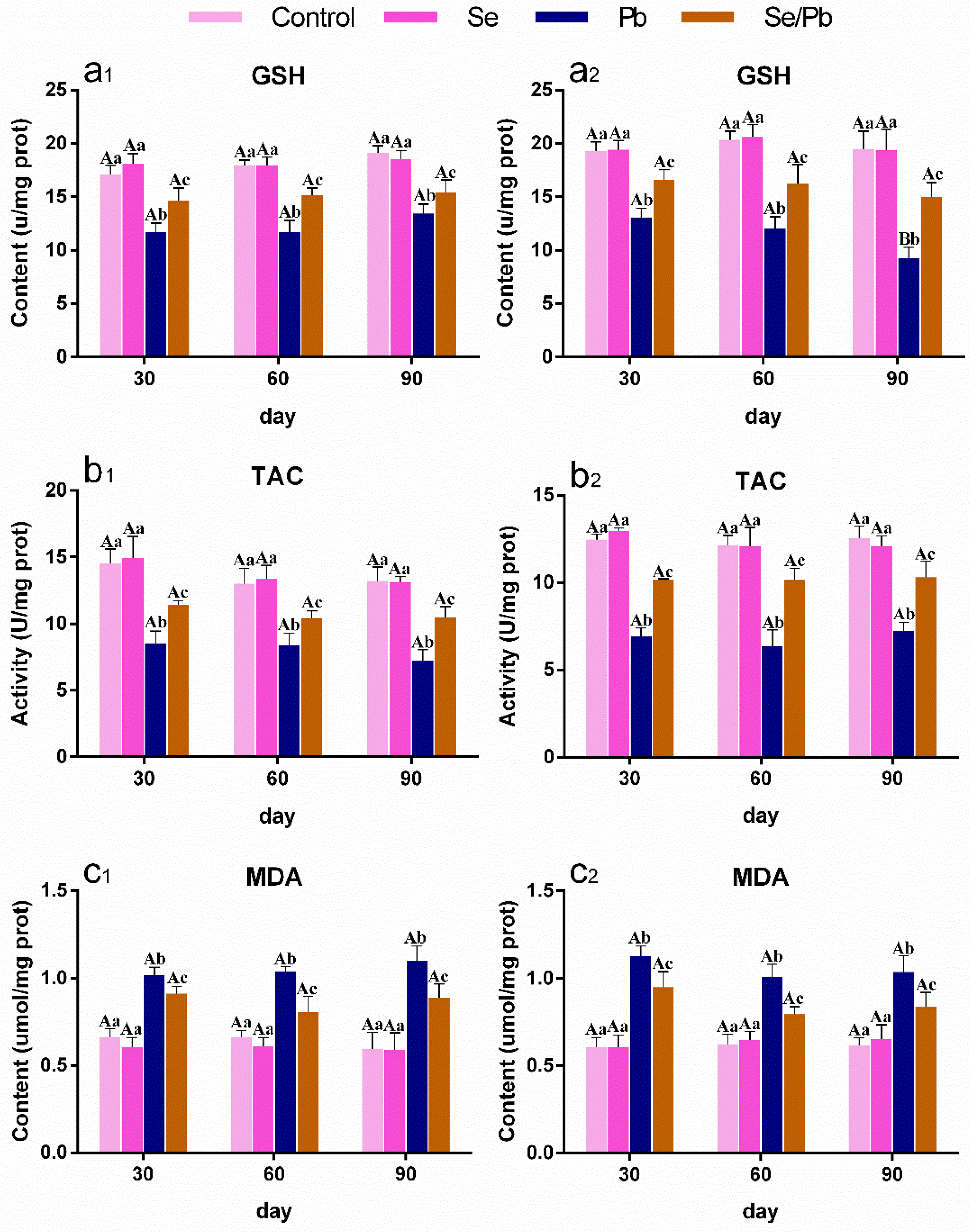
2.4. The Determination of GSH, TAC, MDA, and NO
2.5. The Detection of Selenoproteins and Inflammation-Related Genes
2.5.1. Primer Acquisition and Total RNA Extraction
2.5.2. cDNA Synthesis and PCR Amplification
2.6. Statistical Analysis and IBR Value Calculation
3. Results
3.1. The Symptoms of Chickens
3.2. The Changes in Microstructures in Chicken Ces and Thi
3.3. The Changes in Oxidative Stress-Related Indexes in Chicken Ces and Thi
3.4. The Changes in Inflammatory-Related Factors in Chicken Ces and Thi
3.5. The Changes in Selenoprotein in Chicken Ces and Thi
3.6. The Changes in IBR Values of Selenoproteins, Oxidative Stress Indexes, and Inflammatory-Related Factors in Chicken Ces and Thi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO). Lead Poisoning. Available online: https://www.who.int/news-room/fact-sheets/detail/lead-poisoning-and-health (accessed on 15 January 2023).
- Laubner, G.; Stražnickaitė, I. Case series of chronic occupational lead exposure in shooting ranges. J. Trace. Elem. Med. Biol. 2022, 69, 126886. [Google Scholar] [CrossRef] [PubMed]
- Ansari, J.A.; Mahdi, A.A.; Malik, P.S.; Jafar, T. Blood Lead Levels in Children Living Near an Informal Lead Battery Recycling Workshop in Patna, Bihar. J. Health Pollut. 2020, 10, 200308. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Ma, L.; Yi, Y.; Lin, T. Assessment of heavy metal pollution and exposure risk for migratory birds—A case study of Caohai wetland in Guizhou Plateau (China). Environ. Pollut. 2021, 275, 116564. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yang, W.; Wang, S.; Zhang, H.; Zhang, Y. Does environmental metal pollution affect bird morphometry? A case study on the tree sparrow Passer montanus. Chemosphere 2022, 295, 133947. [Google Scholar] [CrossRef] [PubMed]
- Leão, L.K.R.; Bittencourt, L.O.; Oliveira, A.C.; Nascimento, P.C.; Miranda, G.H.N.; Ferreira, R.O.; Nabiça, M.; Dantas, K.; Dionizio, A.; Cartágenes, S.; et al. Long-Term Lead Exposure Since Adolescence Causes Proteomic and Morphological Alterations in the Cerebellum Associated with Motor Deficits in Adult Rats. Int. J. Mol. Sci. 2020, 21, 3571. [Google Scholar] [CrossRef]
- Albores-Garcia, D.; McGlothan, J.L.; Bursac, Z.; Guilarte, T.R. Chronic developmental lead exposure increases µ-opiate receptor levels in the adolescent rat brain. NeuroToxicology 2021, 82, 119–129. [Google Scholar] [CrossRef]
- Sun, W.; Lei, Y.; Jiang, Z.; Wang, K.; Liu, H.; Xu, T. BPA and low-Se exacerbate apoptosis and mitophagy in chicken pancreatic cells by regulating the PTEN/PI3K/AKT/mTOR pathway. J. Adv. Res. 2024, in press. [CrossRef]
- Wang, D.; Zong, C.; Cheng, K. Chicken thalamic injury induced by copper (II) or/and arsenite exposure involves oxidative stress and inflammation-induced apoptosis. Ecotoxicol. Environ. Saf. 2020, 197, 110554. [Google Scholar] [CrossRef]
- Wang, S.; Hou, L.; Wang, M.; Feng, R.; Lin, X.; Pan, S.; Zhao, Q.; Huang, H. Selenium-Alleviated Testicular Toxicity by Modulating Inflammation; Heat Shock Response; and Autophagy under Oxidative Stress in Lead-Treated Chickens. Biol. Trace Elem. Res. 2021, 199, 4700–4712. [Google Scholar] [CrossRef]
- Chen, X.; Bi, M.; Yang, J.; Cai, J.; Zhang, H.; Zhu, Y.; Zheng, Y.; Liu, Q.; Shi, G.; Zhang, Z. Cadmium exposure triggers oxidative stress; necroptosis; Th1/Th2 imbalance and promotes inflammation through the TNF-α/NF-κB pathway in swine small intestine. J. Hazard. Mater. 2022, 421, 126704. [Google Scholar] [CrossRef]
- Tucovic, D.; Kulas, J.; Mirkov, I.; Popovic, D.; Zolotarevski, L.; Despotovic, M.; Kataranovski, M.; Aleksandra, P.A. Oral Cadmium Intake Enhances Contact Allergen-induced Skin Reaction in Rats. Biomed. Environ. Sci. 2022, 35, 1038–1050. [Google Scholar] [CrossRef]
- Wu, J.; Ding, J.; Shi, Y.; Fang, Y.; Li, P.; Fan, F.; Zhao, E.; Sun, X.; Shen, X.; Hu, Q. Inhibition of immunotoxicity of Pb2+-induced RAW264.7 macrophages by selenium species in selenium-enriched rice. Food Chem. Toxicol. 2021, 148, 111943. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jiao, X.; Xu, Y.; Han, Q.; Jiao, W.; Liu, Y.; Li, S.; Teng, X. Dietary selenium supplementation alleviates immune toxicity in the hearts of chickens with lead-added drinking water. Avian. Pathol. 2019, 48, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Huang, J.; Hu, T.; Zhang, Y.; Zhang, L.; Zhang, J.; Cui, D.; Li, L.; Wang, J.; Wu, Q. Protective Effects of Selenium Nanoparticles against Bisphenol A-Induced Toxicity in Porcine Intestinal Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 7242. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, J.; Zhao, H.; Wang, Y.; Liu, J.; Shao, Y.; Xue, Y.; Xing, M. Synergistic effect of copper and arsenic upon oxidative stress, inflammation and autophagy alterations in brain tissues of Gallus gallus. J. Inorg. Biochem. 2018, 178, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.S.; Talukder, M.; Jin, H.T.; Lv, M.W.; Ge, J.; Zhang, C.; Li, J.L. Cadmium through Disturbing MTF1-Mediated Metal Response Induced Cerebellar Injury. Neurotox. Res. 2022, 40, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Salaramoli, S.; Amiri, H.; Joshaghani, H.R.; Hosseini, M.; Hashemy, S.I. Bio-synthesized selenium nanoparticles ameliorate Brain oxidative stress in Parkinson disease rat models. Metab. Brain Dis. 2023, 38, 2055–2064. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Cao, H.; Xing, C.; Feng, J.; Zhang, L.; Zhang, C.; Hu, G.; Yang, F. Selenium triggers Nrf2-AMPK crosstalk to alleviate cadmium-induced autophagy in rabbit cerebrum. Toxicology 2021, 459, 152855. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, C.; Liu, C.; Teng, X.; Li, S. Selenium Deficiency Mainly Influences Antioxidant Selenoproteins Expression in Broiler Immune Organs. Biol. Trace Elem. Res. 2016, 172, 209–221. [Google Scholar] [CrossRef]
- Bi, M.; Zhang, H.; Yuan, L.; Zhao, L.; Liu, R. Molecular mechanisms of lead-induced changes of selenium status in mice livers through interacting with selenoprotein P. Ecotoxicol. Environ. Saf. 2019, 175, 282–288. [Google Scholar] [CrossRef]
- Chen, D.; Yu, W.; Hao, Z.; Qiu, M.; Cui, J.; Tang, Y.; Teng, X.; Liu, Y.; Liu, H. Molecular mechanism of selenium against lead-induced apoptosis in chicken brainstem relating to heat shock protein, selenoproteins, and inflammatory cytokines. Ecotoxicol. Environ. Saf. 2024, 272, 116028. [Google Scholar] [CrossRef]
- Meng, Z. Basic Environmental Toxicology; Higher Education Press: Beijing, China, 2003; p. 79. [Google Scholar]
- Vengris, V.E.; Mare, C.J. Lead poisoning in chickens and the effect of lead on interferon and antibody production. Can. J. Comp. Med. 1974, 38, 328–335. [Google Scholar] [PubMed]
- Li, X.; Xing, M.; Chen, M.; Zhao, J.; Fan, R.; Zhao, X.; Cao, C.; Yang, J.; Zhang, Z.; Xu, S. Effects of selenium-lead interaction on the gene expression of inflammatory factors and selenoproteins in chicken neutrophils. Ecotoxicol. Environ. Saf. 2017, 139, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Zhu, Q.; Cao, Q.; Bai, Y.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. Quercetin and Allicin Can Alleviate the Hepatotoxicity of Lead (Pb) through the PI3K Signaling Pathway. J. Agric. Food Chem. 2021, 69, 9451–9460. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Xu, W.; Zhang, Y.; Sun, Q.; Li, Z.; Geng, L.; Teng, X. Transcriptome analysis revealed the mechanism of Luciobarbus capito (L. capito) adapting high salinity: Antioxidant capacity, heat shock proteins, immunity. Mar. Pollut. Bull. 2023, 192, 115017. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Luo, D.; Xu, A.; Zhao, B.; Lin, H.; Yao, H.; Li, S. Insight into the mechanism of melatonin in attenuating PCB126-induced liver injury: Resistance to ROS-dependent NETs formation to alleviate inflammation and lipid metabolism dysfunction. Ecotoxicol. Environ. Saf. 2024, 270, 115923. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Guan, H.; Li, D.; Shi, B.; Jiang, Y.; Qiao, S.; Liu, Q.; Fang, C.; Zhang, Z. New insights into Microalgal astaxanthin’s effect on Lambda-cyhalothrin-induced lymphocytes immunotoxicity in Cyprinus carpio: Involving miRNA-194-5p-FoxO1-mediated-mitophagy and pyroptosis. Fish. Shellfish. Immunol. 2023, 141, 109046. [Google Scholar] [CrossRef] [PubMed]
- Pattee, O.H.; Carpenter, J.W.; Fritts, S.H.; Rattner, B.A.; Wiemeyer, S.N.; Royle, J.A.; Smith, M.R. Lead poisoning in captive Andean condors (Vultur gryphus). J. Wildl. Dis. 2006, 42, 772–779. [Google Scholar] [CrossRef]
- Wang, Y.; Weng, Y.; Lv, L.; Wang, D.; Yang, G.; Jin, Y.; Wang, Q. Transgenerational effects of co-exposure to cadmium and carbofuran on zebrafish based on biochemical and transcriptomic analyses. J. Hazard. Mater. 2022, 439, 129644. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, C.; Wang, D.; Zhou, J.; Yang, G.; Shao, K.; Wang, Q.; Jin, Y. Effects of chlorothalonil; prochloraz and the combination on intestinal barrier function and glucolipid metabolism in the liver of mice. J. Hazard. Mater. 2021, 410, 124639. [Google Scholar] [CrossRef]
- Abubakar, K.; Muhammad Mailafiya, M.; Danmaigoro, A.; Musa Chiroma, S.; Abdul Rahim, E.B.; Abu Bakar Zakaria, M.Z. Curcumin Attenuates Lead-Induced Cerebellar Toxicity in Rats via Chelating Activity and Inhibition of Oxidative Stress. Biomolecules 2019, 9, 453. [Google Scholar] [CrossRef]
- Huang, H.; Li, X.; Wang, Z.; Lin, X.; Tian, Y.; Zhao, Q.; Zheng, P. Anti-inflammatory effect of selenium on lead-induced testicular inflammation by inhibiting NLRP3 inflammasome activation in chickens. Theriogenology 2020, 155, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, J.; Wang, Y.L.; Luo, Z.; Zheng, C.; Yu, G.; Wu, S.; Zheng, F.; Li, H. NOX2 activation contributes to cobalt nanoparticles-induced inflammatory responses and Tau phosphorylation in mice and microglia. Ecotoxicol. Environ. Saf. 2021, 225, 112725. [Google Scholar] [CrossRef] [PubMed]
- Hosseinirad, H.; Shahrestanaki, J.K.; Moosazadeh Moghaddam, M.; Mousazadeh, A.; Yadegari, P.; Afsharzadeh, N. Protective Effect of Vitamin D3 against Pb-Induced Neurotoxicity by Regulating the Nrf2 and NF-κB Pathways. Neurotox. Res. 2021, 39, 687–696. [Google Scholar] [CrossRef]
- Albrakati, A.; Alsharif, K.F.; Al Omairi, N.E.; Alsanie, W.F.; Almalki, A.S.A.; Abd Elmageed, Z.Y.; Elshopakey, G.E.; Lokman, M.S.; Bauomy, A.A.; Abdel Moneim, A.E.; et al. Neuroprotective Efficiency of Prodigiosins Conjugated with Selenium Nanoparticles in Rats Exposed to Chronic Unpredictable Mild Stress is Mediated Through Antioxidative, Anti-Inflammatory, Anti-Apoptotic, and Neuromodulatory Activities. Int. J. Nanomed. 2021, 16, 8447–8464. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Tong, J.; Sun, Q.; Teng, X.; Zhang, H.; Teng, X. The involvement of miR-6615-5p/Smad7 axis and immune imbalance in ammonia-caused inflammatory injury via NF-κB pathway in broiler kidneys. Poult. Sci. 2020, 99, 5378–5388. [Google Scholar] [CrossRef] [PubMed]
- Laurence, A.; Tato, C.M.; Davidson, T.S.; Kanno, Y.; Chen, Z.; Yao, Z.; Blank, R.B.; Meylan, F.; Siegel, R.; Hennighausen, L.; et al. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity 2007, 26, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, D.; Cvetkovic, I.; Momcilovic, M.; Maksimovic-Ivanic, D.; Stosic-Grujicic, S.; Trajkovic, V. Interleukin-17 stimulates inducible nitric oxide synthase-dependent toxicity in mouse beta cells. Cell. Mol. Life. Sci. 2005, 62, 2658–2668. [Google Scholar] [CrossRef]
- Wang, H.; Li, S.; Teng, X. The Antagonistic Effect of Selenium on Lead-Induced Inflammatory Factors and Heat Shock Proteins mRNA Expression in Chicken Livers. Biol. Trace Elem. Res. 2016, 171, 437–444. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, F.; Lu, P.; Zhao, R.; Zhang, H.; Song, B.; Li, L.; Wu, Z.; Wu, R. Selenium-Yeast Alleviated Inflammatory Damage Caused by Lead via Inhibiting Ras/ERK Pathway and Inflammatory Factors in Chicken Skeletal Muscles. Biol. Trace Elem. Res. 2019, 190, 493–500. [Google Scholar] [CrossRef]
- Huang, J.; Bao, D.; Lei, C.T.; Tang, H.; Zhang, C.Y.; Su, H.; Zhang, C. Selenoprotein T protects against cisplatin-induced acute kidney injury through suppression of oxidative stress and apoptosis. FASEB J. 2020, 34, 11983–11996. [Google Scholar] [CrossRef]
- Luo, D.; Tang, X.; Wang, Y.; Ying, S.; He, Y.; Lin, H.; Khoso, P.A.; Li, S. Selenium deficiency exacerbated Bisphenol A-induced intestinal toxicity in chickens: Apoptosis and cell cycle arrest mediated by ROS/P53. Sci. Total Environ. 2024, 913, 169730. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lin, H.; Xu, T.; Shi, X.; Yao, Y.; Khoso, P.A.; Jiang, Z.; Xu, S. New insights into brain injury in chickens induced by bisphenol A and selenium deficiency-Mitochondrial reactive oxygen species and mitophagy-apoptosis crosstalk homeostasis. Sci. Total Environ. 2023, 905, 166890. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, S.; Wang, X.; Zhao, B.; Chen, S.; Jiang, Q.; Xu, S.; Li, S. Astilbin targeted Sirt1 to inhibit acetylation of Nrf2 to alleviate grass carp hepatocyte apoptosis caused by PCB126-induced mitochondrial kinetic and metabolism dysfunctions. Fish Shellfish Immunol. 2023, 141, 109000. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, M.; Wang, Y.; Gong, K.; Yan, T.; Wang, D.; Meng, X.; Yang, X.; Chen, Y.; Han, J.; et al. Daidzein alleviates doxorubicin-induced heart failure via the SIRT3/FOXO3a signaling pathway. Food Funct. 2022, 13, 9576–9588. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shen, F.; Liu, J.; Tang, H.; Teng, X.; Yang, F.; Liu, H. Luteolin enhanced antioxidant capability and induced pyroptosis through NF-κB/NLRP3/Caspase-1 in splenic lymphocytes exposure to ammonia. Sci. Total Environ. 2024, 919, 170699. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Hao, Z.; Zhou, Q.; Qiu, M.; Liu, Y.; Liu, Y.; Teng, X.; Kang, L. Chlorpyrifos induced autophagy and mitophagy in common carp livers through AMPK pathway activated by energy metabolism disorder. Ecotoxicol. Environ. Saf. 2023, 258, 114983. [Google Scholar] [CrossRef] [PubMed]
- Hadzic, T.; Li, L.; Cheng, N.; Walsh, S.A.; Spitz, D.R.; Knudson, C.M. The role of low molecular weight thiols in T lymphocyte proliferation and IL-2 secretion. J. Immunol. 2005, 175, 7965–7972. [Google Scholar] [CrossRef]
- Zhang, C.; Ge, J.; Lv, M.; Zhang, Q.; Talukder, M.; Li, J.L. Selenium prevent cadmium-induced hepatotoxicity through modulation of endoplasmic reticulum-resident selenoproteins and attenuation of endoplasmic reticulum stress. Environ. Pollut. 2020, 260, 113873. [Google Scholar] [CrossRef]
- Talukder, M.; Bi, S.S.; Jin, H.T.; Ge, J.; Zhang, C.; Lv, M.W.; Li, J.L. Cadmium induced cerebral toxicity via modulating MTF1-MTs regulatory axis. Environ. Pollut. 2021, 285, 117083. [Google Scholar] [CrossRef]
- Xu, X.M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef]
- Fan, R.; Yao, H.; Zhao, X.; Cao, C.; Yang, T.; Luan, Y.; Zhang, Z.; Xu, S. Gene expression of selenoproteins can be regulated by selenoprotein K silencing in chicken myoblasts. BioMetals 2016, 29, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Sagliocchi, S.; Cicatiello, A.G.; Di Cicco, E.; Ambrosio, R.; Miro, C.; Di Girolamo, D.; Nappi, A.; Mancino, G.; De Stefano, M.A.; Luongo, C.; et al. The thyroid hormone activating enzyme, type 2 deiodinase, induces myogenic differentiation by regulating mitochondrial metabolism and reducing oxidative stress. Redox Biol. 2019, 24, 101228. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, X.; Liu, Q.; Wang, Y.; Li, S.; Xu, S. Selenoprotein K protects skeletal muscle from damage and is required for satellite cells-mediated myogenic differentiation. Redox Biol. 2022, 50, 102255. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.V.; Shin, E.J.; Jeong, J.H.; Lee, J.W.; Lee, Y.; Jang, C.G.; Nah, S.Y.; Lei, X.G.; Toriumi, K.; Yamada, K.; et al. Protective Potential of the Glutathione Peroxidase-1 Gene in Abnormal Behaviors Induced by Phencyclidine in Mice. Mol. Neurobiol. 2017, 54, 7042–7062. [Google Scholar] [CrossRef]
- Chen, H.; Li, P.; Shen, Z.; Wang, J.; Diao, L. Protective effects of selenium yeast against cadmium-induced necroptosis through miR-26a-5p/PTEN/PI3K/AKT signaling pathway in chicken kidney. Ecotoxicol. Environ. Saf. 2021, 220, 112387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Hao, D.; Wang, J.; Zhu, R.; Liu, W.; Liu, C. Selenium regulates the mitogen-activated protein kinase pathway to protect broilers from hexavalent chromium-induced kidney dysfunction and apoptosis. Ecotoxicol. Environ. Saf. 2022, 239, 113629. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Wang, J.; Wang, Y.; Liu, H.; Bao, J. Selenium Alleviates Ammonia-Induced Splenic Cell Apoptosis and Inflammation by Regulating the Interleukin Family/Death Receptor Axis and Nrf2 Signaling Pathway. Biol. Trace Elem. Res. 2023, 201, 1748–1760. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Jiang, Y.; Shi, B.; Zhang, Z.; Zhang, Z. Selenomethionine alleviates decabromodiphenyl ether-induced oxidative stress and ferroptosis via the NRF2/GPx4 pathway in the chicken brain. J. Hazard. Mater. 2024, 465, 133307. [Google Scholar] [CrossRef]
- Jing, J.; Zeng, H.; Shao, Q.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Selenomethionine alleviates environmental heat stress induced hepatic lipid accumulation and glycogen infiltration of broilers via maintaining mitochondrial and endoplasmic reticulum homeostasis. Redox Biol. 2023, 67, 102912. [Google Scholar] [CrossRef]
- Eid, Y.Z.; Omara, Y.; Ragab, A.; Ismail, A.; Zommara, M.; Dawood, M.A.O. Mitigation of Imidacloprid Toxicity in Poultry Chicken by Selenium Nanoparticles: Growth Performance, Lipid Peroxidation, and Blood Traits. Biol. Trace Elem. Res. 2023, 201, 5379–5388. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Serial Numbers | Primer Sequences | |
|---|---|---|---|
| β-actin | L08165 | F: 5’-CCGCTCTATGAAGGCTACGC-3’ | R: 5’-CTC TCG GCT GTG GTGGTG AA-3’ |
| GPx1 | NM_001277853.1 | F: 5’-ACGGCGCATCTTCCAAAG-3’ | R: 5’-TGTTCCCCCAACCATTTCTC-3’ |
| GPx2 | NM_001277854.1 | F: 5’-ATCGCCAAGTCCTTCTACGA-3’ | R: 5’-ACGTTCTCGATGAGGACCAC-3’ |
| GPx3 | NM_001163232.2 | F: 5’-CCTGCAGTACCTCGAACTGA-3’ | R: 5’-CTTCAGTGCAGGGAGGATCT-3’ |
| GPx4 | AF498316.2 | F: 5’-CTTCGTCTGCATCATCACCAA-3’ | R: 5’-TCGACGAGCTGAGTGTAATTCAC-3’ |
| Dio1 | NM_001097614.1 | F: 5’-GCGCTATACCACAGGCAGTA-3’ | R: 5’-GGTCTTGCAAATGTCACCAC-3’ |
| Dio2 | NM_204114.3 | F: 5’-ATTTGCTGATCACGCTTCAG-3’ | R: 5’-GCTCAGAAACAGCACCATGT-3’ |
| Dio3 | NM_001122648.1 | F: 5’-CTGTGCATTCGCAAGAAGAT-3’ | R: 5’-GCCGACTTGAAGAAGTCCAG-3’ |
| SelH | NM_001277865.1 | F: 5’- CATCGAGCACTGCCGTAG-3’ | R: 5’- GACACCTCGAAGCTGTTCCT-3’ |
| SelI | NM_001031528.2 | F: 5’-TGCCAGCCTCTGAACTGGAT-3’ | R: 5’-TGCAAACCCAGACATCACCAT-3’ |
| SelK | NM_001025441.2 | F: 5’- ATGACGACCACCCTCACGAT-3’ | R: 5’- CCAGCGTTAACCGGAATGAT-3 |
| SelM | NM_001277859.1 | F: 5’-AAGAAGGACCACCCAGACCT-3’ | R: 5’-GCTGTCCTGTCTCCCTCATC-3’ |
| SelO | NM_001115017.1 | F: 5’-CCAGCGTTAACCGGAATGAT-3’ | R: 5’-GCCTACAGAATGGATCCAACTGA-3’ |
| SelS | NM_173120.2 | F: 5’-GCCTGCGTCGCCATCTATCTCA-3’ | R: 5’-TTCTGCCTTCGCTTCTGTTCTTCAA-3’ |
| SelT | NM_001006557.3 | F: 5’-AGGAGTACATGCGGGTCATCA-3’ | R: 5’-GACAGACAGGAAGGATGCTATGTG-3’ |
| SelU | NM_001193518.1 | F: 5’-GATGCTTTCAGGCTTCTTCC-3’ | R: 5’-CTGTCTTCCTGCTCCAATCA-3’ |
| SelPb | XM_003641687.2 | F: 5’-AGGCCAACAGTACCATGGAG-3’ | R: 5’-GTGGTGAGGATGGAGATGGT-3’ |
| Sepn1 | NM_001114972.1 | F: 5’-CAGGATCCATGCTGAGTTCCA-3’ | R: 5’-GAGAGGACGATGTAACCCGTAAAC-3’ |
| SelP1 | NM_001031609 | F: 5’-CCAAGTGGTCAGCATTCACATC-3’ | R: 5’-ATGACGACCACCCTCACGAT-3’ |
| Sepx1 | NM_001135558.2 | F: 5’-TGGCAAGTGTGGCAATGG-3’ | R: 5’-GAATTTGAGCGAGCTGCTGAAT-3’ |
| Sepw1 | NM_001166327.1 | F: 5’-TGGTGTGGGTCTGCTTTACG-3’ | R: 5’-CCAAAGCTGGAAGGTGCAA-3’ |
| Sep15 | NM_001012926.2 | F: 5’-ACTTGGCTTCTCCAGTAACTTGCT-3’ | R: 5’-GCCTACAGAATGGATCCAACTGA-3’ |
| SPS2 | BM489698.1 | F: 5’-CGTTGGGTATCGGAACTGAC-3’ | R: 5’-CGTCCACCAGAGGGTAGAAA-3’ |
| IL-2 | AY510091 | F:5′-GAACCTCAAGAGTCTTACGGGTCTA-3′ | R: 5′-ACAAAGTTGGTCAGTTCATGGAGA-3′ |
| IL-4 | AJ621249 | F: 5′-GTGCCCACGCTGTGCTTAC-3′ | R: 5′-AGGAAACCTCTCCCTGGATGTC-3′ |
| IL-6 | NM204628 | F: 5′-AAATCCCTCCTCGCCAATCT-3′ | R: 5′-CCCTCACGGTCTTCTCCATAAA-3′ |
| IL-12β | AJ564201.1 | F: 5′-TGTCTCACCTGCTATTTGCCTTAC-3′ | R:5′-CATACACATTCTCTCTAAGTTTCCACTGT-3′ |
| IL-17 | AY744450 | F: 5′-CATGTTGTCAGCCAGCATTTCT-3′ | R: 5′-CATCTTTTTGGGTTAGGCATCC-3′ |
| INF-γ | DQ470471 | F: 5′-AAGTCATAGCGGCACATCAAAC-3′ | R: 5′-CTGGAATCTCATGTCGTTCATCG-3′ |
| Organs | Items | Days | The Control Group | The Se Group | The Pb Group | The Se/Pb Group |
|---|---|---|---|---|---|---|
| Cerebellum | GPx1 | 30 d | 0.98 ± 0.04 Aa | 1.05 ± 0.04 Aa | 0.62 ± 0.05 Ab | 0.83 ± 0.01 Ac |
| 60 d | 1.01 ± 0.01 Aa | 0.97 ± 0.04 Aa | 0.45 ± 0.11 ABb | 0.72 ± 0.02 Bc | ||
| 90 d | 1.05 ± 0.05 Aa | 1.00 ± 0.04 Aa | 0.31 ± 0.01 Bb | 0.72 ± 0.02 Bc | ||
| GPx2 | 30 d | 1.00 ± 0.03 Aa | 0.98 ± 0.02 Aa | 0.55 ± 0.05 Ab | 0.65 ± 0.02 Ac | |
| 60 d | 1.01 ± 0.02 Aa | 1.03 ± 0.02 Aa | 0.41 ± 0.04 Bb | 0.68 ± 0.06 Ac | ||
| 90 d | 1.04 ± 0.06 Aa | 0.98 ± 0.03 Aa | 0.43 ± 0.01 Bb | 0.67 ± 0.02 Ac | ||
| GPx3 | 30 d | 0.97 ± 0.02 Aa | 1.01 ± 0.04 Aa | 0.72 ± 0.07 Ab | 0.94 ± 0.05 Aa | |
| 60 d | 1.03 ± 0.02 Aa | 1.02 ± 0.02 Aa | 0.68 ± 0.06 Ab | 0.88 ± 0.02 Ac | ||
| 90 d | 1.00 ± 0.08 Aa | 1.04 ± 0.01 Aa | 0.64 ± 0.05 Ab | 0.77 ± 0.05 Ac | ||
| GPx4 | 30 d | 1.00 ± 0.02 Aa | 1.07 ± 0.03 Aa | 0.79 ± 0.02 Ab | 0.80 ± 0.06 Ab | |
| 60 d | 1.02 ± 0.06 Aa | 1.03 ± 0.01 Aa | 0.67 ± 0.03 Bb | 0.78 ± 0.02 Ac | ||
| 90 d | 1.02 ± 0.01 Aa | 1.07 ± 0.02 Aa | 0.69 ± 0.04 Bb | 0.80 ± 0.05 Ac | ||
| Dio1 | 30 d | 0.99 ± 0.03 Aa | 1.02 ± 0.02 Aa | 0.27 ± 0.03 Ab | 0.71 ± 0.06 Ac | |
| 60 d | 0.98 ± 0.02 Aa | 1.00 ± 0.02 Aa | 0.26 ± 0.02 Ab | 0.75 ± 0.02 Ac | ||
| 90 d | 1.02 ± 0.03 Aa | 1.04 ± 0.02 Aa | 0.26 ± 0.01 Ab | 0.66 ± 0.04 Ac | ||
| Dio2 | 30 d | 1.01 ± 0.02 Aa | 1.04 ± 0.05 Aa | 0.42 ± 0.06 Ab | 0.74 ± 0.03 Ac | |
| 60 d | 0.99 ± 0.02 Aa | 0.98 ± 0.04 Aa | 0.35 ± 0.06 Ab | 0.74 ± 0.08 Ac | ||
| 90 d | 1.02 ± 0.03 Aa | 1.04 ± 0.03 Aa | 0.39 ± 0.07 Ab | 0.73 ± 0.08 Ac | ||
| Dio3 | 30 d | 0.97 ± 0.05 Aa | 1.07 ± 0.02 Aa | 0.41 ± 0.07 Ab | 0.81 ± 0.05 Ac | |
| 60 d | 0.96 ± 0.05 Aa | 1.00 ± 0.06 Aa | 0.36 ± 0.06 Ab | 0.76 ± 0.05 Ac | ||
| 90 d | 1.03 ± 0.01 Aa | 1.02 ± 0.04 Aa | 0.37 ± 0.01 Ab | 0.75 ± 0.03 Ac | ||
| SelH | 30 d | 0.99 ± 0.03 Aa | 1.02 ± 0.05 Aa | 0.64 ± 0.01 Ab | 0.72 ± 0.06 Ac | |
| 60 d | 1.01 ± 0.02 Aa | 0.98 ± 0.09 Aa | 0.61 ± 0.01 Ab | 0.73 ± 0.03 Ac | ||
| 90 d | 1.01 ± 0.06 Aa | 1.05 ± 0.03 Aa | 0.60 ± 0.03 Ab | 0.72 ± 0.02 Ac | ||
| SelI | 30 d | 1.02 ± 0.01 Aa | 1.04 ± 0.03 Aa | 0.51 ± 0.07 Ab | 0.85 ± 0.01 Ac | |
| 60 d | 0.99 ± 0.06 Aa | 1.01 ± 0.05 Aa | 0.48 ± 0.02 Ab | 0.81 ± 0.05 Ac | ||
| 90 d | 1.01 ± 0.03 Aa | 1.05 ± 0.05 Aa | 0.46 ± 0.02 Bb | 0.82 ± 0.08 Ac | ||
| SelK | 30 d | 0.98 ± 0.02 Aa | 1.05 ± 0.04 Aa | 0.64 ± 0.03 Ab | 0.85 ± 0.04 Ac | |
| 60 d | 1.01 ± 0.09 Aa | 0.99 ± 0.05 Aa | 0.65 ± 0.03 Ab | 0.81 ± 0.03 Ac | ||
| 90 d | 1.04 ± 0.06 Aa | 1.06 ± 0.01 Aa | 0.66 ± 0.03 Ab | 0.78 ± 0.01 Ac | ||
| SelM | 30 d | 1.00 ± 0.04 Aa | 1.04 ± 0.01 Aa | 0.54 ± 0.09 Ab | 0.83 ± 0.04 Ac | |
| 60 d | 0.98 ± 0.04 Aa | 1.03 ± 0.01 Aa | 0.50 ± 0.02 Ab | 0.83 ± 0.05 Ac | ||
| 90 d | 1.04 ± 0.04 Aa | 1.06 ± 0.02 Aa | 0.49 ± 0.03 Ab | 0.84 ± 0.02 Ac | ||
| SelO | 30 d | 1.01 ± 0.03 Aa | 1.04 ± 0.02 Aa | 0.62 ± 0.05 Ab | 0.85 ± 0.04 Ac | |
| 60 d | 0.99 ± 0.05 Aa | 1.03 ± 0.05 Aa | 0.65 ± 0.05 Ab | 0.83 ± 0.03 Ac | ||
| 90 d | 1.05 ± 0.04 Aa | 1.01 ± 0.05 Aa | 0.56 ± 0.04 Ab | 0.80 ± 0.01 Ac | ||
| SelS | 30 d | 1.01 ± 0.04 Aa | 1.01 ± 0.06 Aa | 0.45 ± 0.02 Ab | 0.85 ± 0.05 Ac | |
| 60 d | 1.00 ± 0.02 Aa | 1.02 ± 0.01 Aa | 0.43 ± 0.05 Ab | 0.86 ± 0.20 Ac | ||
| 90 d | 1.01 ± 0.05 Aa | 1.01 ± 0.05 Aa | 0.40 ± 0.08 Ab | 0.84 ± 0.04 Ac | ||
| SelT | 30 d | 1.00 ± 0.03 Aa | 1.02 ± 0.01 Aa | 0.61 ± 0.03 Ab | 0.82 ± 0.07 Ac | |
| 60 d | 0.99 ± 0.04 Aa | 1.04 ± 0.05 Aa | 0.56 ± 0.05 Ab | 0.82 ± 0.02 Ac | ||
| 90 d | 1.03 ± 0.05 Aa | 0.98 ± 0.03 Aa | 0.55 ± 0.05 Ab | 0.70 ± 0.06 Ac | ||
| SelU | 30 d | 0.97 ± 0.06 Aa | 0.99 ± 0.06 Aa | 0.61 ± 0.07 Ab | 0.79 ± 0.04 Ac | |
| 60 d | 0.99 ± 0.06 Aa | 1.03 ± 0.05 Aa | 0.56 ± 0.02 Ab | 0.72 ± 0.03 ABc | ||
| 90 d | 1.00 ± 0.06 Aa | 1.04 ± 0.02 Aa | 0.51 ± 0.04 Ab | 0.67 ± 0.03 Bc | ||
| SelPb | 30 d | 0.96 ± 0.02 Aa | 1.08 ± 0.02 Ab | 0.60 ± 0.06 Ac | 0.70 ± 0.03 Ad | |
| 60 d | 0.97 ± 0.05 Aa | 1.06 ± 0.03 Aa | 0.62 ± 0.05 Ab | 0.72 ± 0.02 Ac | ||
| 90 d | 1.00 ± 0.03 Aa | 1.06 ± 0.01 Aa | 0.58 ± 0.04 Ab | 0.76 ± 0.02 Ac | ||
| Sepn1 | 30 d | 0.98 ± 0.04 Aa | 0.99 ± 0.03 Aa | 0.49 ± 0.05 Ab | 0.84 ± 0.01 Ac | |
| 60 d | 1.00 ± 0.01 Aa | 0.99 ± 0.02 Aa | 0.43 ± 0.05 Ab | 0.84 ± 0.03 Ac | ||
| 90 d | 0.99 ± 0.01 Aa | 1.03 ± 0.01 Aa | 0.42 ± 0.07 Ab | 0.85 ± 0.02 Ac | ||
| SelP1 | 30 d | 1.01 ± 0.03 Aa | 1.02 ± 0.03 Aa | 0.54 ± 0.02 Ab | 0.77 ± 0.02 Ac | |
| 60 d | 0.96 ± 0.10 Aa | 1.05 ± 0.02 Aa | 0.50 ± 0.07 Ab | 0.74 ± 0.04 Ac | ||
| 90 d | 1.01 ± 0.03 Aa | 1.04 ± 0.03 Aa | 0.46 ± 0.02 Ab | 0.66 ± 0.04 Bc | ||
| Sepx1 | 30 d | 1.02 ± 0.05 Aa | 1.07 ± 0.02 Aa | 0.73 ± 0.08 Ab | 0.79 ± 0.05 Ab | |
| 60 d | 0.98 ± 0.02 Aa | 1.03 ± 0.05 Aa | 0.53 ± 0.02 Ab | 0.79 ± 0.03 Ac | ||
| 90 d | 1.03 ± 0.04 Aa | 1.05 ± 0.03 Aa | 0.44 ± 0.04 Bb | 0.83 ± 0.05 Ac | ||
| Sepw1 | 30 d | 0.98 ± 0.03 Aa | 1.04 ± 0.03 Aa | 0.74 ± 0.04 Ab | 0.86 ± 0.02 Ac | |
| 60 d | 1.06 ± 0.05 Aa | 1.07 ± 0.02 Aa | 0.63 ± 0.03 Bb | 0.88 ± 0.07 Ac | ||
| 90 d | 1.01 ± 0.02 Aa | 1.03 ± 0.05 Aa | 0.54 ± 0.04 Bb | 0.90 ± 0.09 Ac | ||
| Sep15 | 30 d | 1.00 ± 0.04 Aa | 1.00 ± 0.05 Aa | 0.47 ± 0.03 Ab | 0.71 ± 0.04 Ac | |
| 60 d | 1.02 ± 0.02 Aa | 1.06 ± 0.01 Aa | 0.32 ± 0.03 Bb | 0.68 ± 0.03 Ac | ||
| 90 d | 1.01 ± 0.02 Aa | 1.02 ± 0.07 Aa | 0.27 ± 0.04 Bb | 0.63 ± 0.07 Ac | ||
| SPS2 | 30 d | 1.00 ± 0.03 Aa | 0.99 ± 0.04 Aa | 0.75 ± 0.03 Ab | 0.86 ± 0.05 Ac | |
| 60 d | 1.06 ± 0.03 Aa | 1.05 ± 0.03 Aa | 0.66 ± 0.02 Bb | 0.80 ± 0.03 ABc | ||
| 90 d | 1.04 ± 0.04 Aa | 1.00 ± 0.01 Aa | 0.46 ± 0.03 Cb | 0.73 ± 0.01 Bc | ||
| Thalamus | GPx1 | 30 d | 0.98 ± 0.02 Aa | 1.03 ± 0.02 Aa | 0.68 ± 0.04 Ab | 0.82 ± 0.03 Ac |
| 60 d | 1.05 ± 0.03 Aa | 1.04 ± 0.01 Aa | 0.65 ± 0.01 Ab | 0.87 ± 0.02 Ac | ||
| 90 d | 1.00 ± 0.03 Aa | 1.04 ± 0.04 Aa | 0.48 ± 0.05 Ab | 0.86 ± 0.02 Ac | ||
| GPx2 | 30 d | 1.01 ± 0.04 Aa | 1.05 ± 0.02 Aa | 0.61 ± 0.04 Ab | 0.80 ± 0.02 Ac | |
| 60 d | 1.05 ± 0.04 Aa | 1.06 ± 0.03 Aa | 0.57 ± 0.09 Ab | 0.81 ± 0.03 Ac | ||
| 90 d | 1.03 ± 0.03 Aa | 1.05 ± 0.02 Aa | 0.55 ± 0.04 Ab | 0.83 ± 0.03 Ac | ||
| GPx3 | 30 d | 0.99 ± 0.03 Aa | 1.03 ± 0.02 Aa | 0.74 ± 0.08 Ab | 0.81 ± 0.03 Ac | |
| 60 d | 1.01 ± 0.02 Aa | 1.03 ± 0.01 Aa | 0.76 ± 0.03 Ab | 0.85 ± 0.08 Ac | ||
| 90 d | 1.03 ± 0.02 Aa | 1.01 ± 0.04 Aa | 0.77 ± 0.08 Ab | 0.83 ± 0.04 Ac | ||
| GPx4 | 30 d | 1.03 ± 0.05 Aa | 1.05 ± 0.04 Aa | 0.52 ± 0.08 Ab | 0.85 ± 0.02 Ac | |
| 60 d | 1.03 ± 0.01 Aa | 1.03 ± 0.01 Aa | 0.73 ± 0.03 Ab | 0.85 ± 0.03 Ac | ||
| 90 d | 1.02 ± 0.04 Aa | 1.02 ± 0.02 Aa | 0.72 ± 0.04 Ab | 0.81 ± 0.02 Ac | ||
| Dio1 | 30 d | 0.97 ± 0.02 Aa | 1.02 ± 0.04 Aa | 0.38 ± 0.04 Ab | 0.85 ± 0.01 Ac | |
| 60 d | 1.03 ± 0.03 Aa | 1.06 ± 0.03 Aa | 0.34 ± 0.02 Ab | 0.85 ± 0.03 Ac | ||
| 90 d | 1.00 ± 0.03 Aa | 1.04 ± 0.03 Aa | 0.34 ± 0.01 Ab | 0.87 ± 0.03 Ac | ||
| Dio2 | 30 d | 1.00 ± 0.06 Aa | 1.07 ± 0.02 Aa | 0.59 ± 0.05 Ab | 0.90 ± 0.03 Ac | |
| 60 d | 1.04 ± 0.03 Aa | 1.06 ± 0.02 Aa | 0.51 ± 0.04 ABb | 0.92 ± 0.04 Ac | ||
| 90 d | 1.01 ± 0.03 Aa | 1.04 ± 0.01 Aa | 0.47 ± 0.02 Bb | 0.95 ± 0.02 Ac | ||
| Dio3 | 30 d | 1.02 ± 0.01 Aa | 1.06 ± 0.02 Aa | 0.55 ± 0.07 Ab | 0.83 ± 0.05 Ac | |
| 60 d | 1.04 ± 0.04 Aa | 1.02 ± 0.08 Aa | 0.47 ± 0.04 ABb | 0.76 ± 0.06 Ac | ||
| 90 d | 1.04 ± 0.03 Aa | 1.01 ± 0.03 Aa | 0.36 ± 0.01 Bb | 0.76 ± 0.06 Ac | ||
| SelH | 30 d | 0.99 ± 0.03 Aa | 1.06 ± 0.03 Aa | 0.56 ± 0.03 Ab | 0.91 ± 0.05 Ac | |
| 60 d | 1.03 ± 0.04 Aa | 1.04 ± 0.01 Aa | 0.46 ± 0.03 Bb | 0.87 ± 0.06 Ac | ||
| 90 d | 1.01 ± 0.02 Aa | 1.04 ± 0.02 Aa | 0.36 ± 0.03 Cb | 0.85 ± 0.02 Ac | ||
| SelI | 30 d | 1.00 ± 0.02 Aa | 1.03 ± 0.03 Aa | 0.37 ± 0.03 Ab | 0.96 ± 0.03 Ac | |
| 60 d | 1.03 ± 0.02 Aa | 1.04 ± 0.03 Aa | 0.36 ± 0.02 Ab | 0.88 ± 0.05 Ac | ||
| 90 d | 1.04 ± 0.04 Aa | 1.00 ± 0.02 Aa | 0.30 ± 0.05 Ab | 0.88 ± 0.06 Ac | ||
| SelK | 30 d | 0.98 ± 0.06 Aa | 1.05 ± 0.02 Aa | 0.47 ± 0.04 Ab | 0.87 ± 0.04 Ac | |
| 60 d | 1.02 ± 0.03 Aa | 1.04 ± 0.04 Aa | 0.48 ± 0.03 Ab | 0.85 ± 0.02 Ac | ||
| 90 d | 0.98 ± 0.03 Aa | 1.06 ± 0.03 Aa | 0.46 ± 0.03 Ab | 0.87 ± 0.02 Ac | ||
| SelM | 30 d | 0.98 ± 0.05 Aa | 1.06 ± 0.04 Aa | 0.54 ± 0.01 Ab | 0.89 ± 0.02 Ac | |
| 60 d | 1.04 ± 0.03 Aa | 1.05 ± 0.02 Aa | 0.34 ± 0.04 Bb | 0.87 ± 0.04 ABc | ||
| 90 d | 1.03 ± 0.03 Aa | 1.05 ± 0.05 Aa | 0.36 ± 0.03 Bb | 0.75 ± 0.08 Bc | ||
| SelO | 30 d | 0.99 ± 0.04 Aa | 1.05 ± 0.03 Aa | 0.50 ± 0.02 Ab | 0.75 ± 0.02 Ac | |
| 60 d | 0.99 ± 0.03 Aa | 1.03 ± 0.02 Aa | 0.44 ± 0.02 Bb | 0.73 ± 0.03 Ac | ||
| 90 d | 1.04 ± 0.05 Aa | 1.03 ± 0.02 Aa | 0.37 ± 0.02 Cb | 0.77 ± 0.02 Ac | ||
| SelS | 30 d | 1.02 ± 0.02 Aa | 1.04 ± 0.01 Aa | 0.51 ± 0.06 Ab | 0.85 ± 0.02 Ac | |
| 60 d | 1.01 ± 0.03 Aa | 1.05 ± 0.01 Aa | 0.34 ± 0.09 Bb | 0.87 ± 0.03 Ac | ||
| 90 d | 1.03 ± 0.01 Aa | 1.02 ± 0.05 Aa | 0.25 ± 0.02 Bb | 0.88 ± 0.06 Ac | ||
| SelT | 30 d | 1.00 ± 0.02 Aa | 1.01 ± 0.06 Aa | 0.44 ± 0.01 Ab | 0.78 ± 0.02 Ac | |
| 60 d | 1.06 ± 0.03 Aa | 1.03 ± 0.01 Aa | 0.41 ± 0.00 ABb | 0.77 ± 0.02 Ac | ||
| 90 d | 1.03 ± 0.05 Aa | 1.03 ± 0.02 Aa | 0.35 ± 0.04 Bb | 0.77 ± 0.01 Ac | ||
| SelU | 30 d | 1.02 ± 0.04 Aa | 1.04 ± 0.02 Aa | 0.75 ± 0.03 Ab | 0.92 ± 0.04 Ac | |
| 60 d | 1.01 ± 0.03 Aa | 1.04 ± 0.03 Aa | 0.60 ± 0.02 Bb | 0.94 ± 0.02 Ac | ||
| 90 d | 1.01 ± 0.01 Aa | 1.01 ± 0.03 Aa | 0.54 ± 0.03 Cb | 0.89 ± 0.01 Ac | ||
| SelPb | 30 d | 1.00 ± 0.02 Aa | 1.02 ± 0.02 Aa | 0.48 ± 0.01 Ab | 0.85 ± 0.02 Ac | |
| 60 d | 1.01 ± 0.01 Aa | 1.04 ± 0.02 Aa | 0.38 ± 0.02 Bb | 0.84 ± 0.01 Ac | ||
| 90 d | 1.00 ± 0.03 Aa | 1.01 ± 0.03 Aa | 0.25 ± 0.03 Cb | 0.88 ± 0.02 Ac | ||
| Sepn1 | 30 d | 1.02 ± 0.04 Aa | 1.04 ± 0.05 Aa | 0.34 ± 0.03 Ab | 0.83 ± 0.04 Ac | |
| 60 d | 1.00 ± 0.02 Aa | 1.02 ± 0.03 Aa | 0.32 ± 0.02 Ab | 0.84 ± 0.03 Ac | ||
| 90 d | 1.01 ± 0.03 Aa | 1.04 ± 0.01 Aa | 0.22 ± 0.01 Bb | 0.80 ± 0.02 Ac | ||
| SelP1 | 30 d | 0.97 ± 0.06 Aa | 1.03 ± 0.01 Aa | 0.40 ± 0.04 Ab | 0.88 ± 0.03 Ac | |
| 60 d | 1.02 ± 0.03 Aa | 1.05 ± 0.04 Aa | 0.26 ± 0.03 Bb | 0.87 ± 0.02 Ac | ||
| 90 d | 1.03 ± 0.02 Aa | 1.04 ± 0.03 Aa | 0.26 ± 0.03 Bb | 0.89 ± 0.04 Ac | ||
| Sepx1 | 30 d | 1.04 ± 0.05 Aa | 1.01 ± 0.02 Aa | 0.48 ± 0.01 Ab | 0.71 ± 0.04 Ac | |
| 60 d | 0.99 ± 0.04 Aa | 1.05 ± 0.02 Aa | 0.52 ± 0.14 Ab | 0.92 ± 0.03 Ac | ||
| 90 d | 1.02 ± 0.06 Aa | 1.01 ± 0.08 Aa | 0.43 ± 0.05 Ab | 0.85 ± 0.03 Ac | ||
| Sepw1 | 30 d | 0.98 ± 0.04 Aa | 1.03 ± 0.02 Aa | 0.47 ± 0.02 Ab | 0.81 ± 0.02 Ac | |
| 60 d | 1.03 ± 0.05 Aa | 1.06 ± 0.04 Aa | 0.36 ± 0.05 Bb | 0.87 ± 0.05 Ac | ||
| 90 d | 1.05 ± 0.01 Aa | 1.03 ± 0.02 Aa | 0.37 ± 0.01 Bb | 0.84 ± 0.03 Ac | ||
| Sep15 | 30 d | 0.99 ± 0.02 Aa | 1.03 ± 0.03 Aa | 0.47 ± 0.03 Ab | 0.77 ± 0.04 Ac | |
| 60 d | 1.00 ± 0.01 Aa | 1.03 ± 0.01 Aa | 0.37 ± 0.06 Ab | 0.77 ± 0.01 Ac | ||
| 90 d | 1.03 ± 0.02 Aa | 1.00 ± 0.01 Aa | 0.23 ± 0.03 Ab | 0.73 ± 0.01 Ac | ||
| SPS2 | 30 d | 0.97 ± 0.07 Aa | 0.99 ± 0.04 Aa | 0.38 ± 0.02 Ab | 0.86 ± 0.05 Ac | |
| 60 d | 1.00 ± 0.06 Aa | 1.04 ± 0.02 Aa | 0.36 ± 0.03 Ab | 0.83 ± 0.03 Ac | ||
| 90 d | 0.96 ± 0.04 Aa | 0.99 ± 0.05 Aa | 0.26 ± 0.09 Ab | 0.78 ± 0.07 Ac |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Liang, J.; Jiang, C.; Cui, J.; Hong, L.; Hao, Z.; Tang, Y.; Liu, Y.; Cui, X.; Teng, X. Se Alleviated Pb-Caused Neurotoxicity in Chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO Pathway, Selenoprotein Suppression, Oxidative Stress, and Inflammatory Injury. Antioxidants 2024, 13, 370. https://doi.org/10.3390/antiox13030370
Li Y, Liang J, Jiang C, Cui J, Hong L, Hao Z, Tang Y, Liu Y, Cui X, Teng X. Se Alleviated Pb-Caused Neurotoxicity in Chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO Pathway, Selenoprotein Suppression, Oxidative Stress, and Inflammatory Injury. Antioxidants. 2024; 13(3):370. https://doi.org/10.3390/antiox13030370
Chicago/Turabian StyleLi, Yansheng, Jiatian Liang, Chunyu Jiang, Jiawen Cui, Lan Hong, Zhiyu Hao, You Tang, Yuhao Liu, Xun Cui, and Xiaohua Teng. 2024. "Se Alleviated Pb-Caused Neurotoxicity in Chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO Pathway, Selenoprotein Suppression, Oxidative Stress, and Inflammatory Injury" Antioxidants 13, no. 3: 370. https://doi.org/10.3390/antiox13030370