
Comparison of the Antioxidant Potency of Four Triterpenes of Centella asiatica against Oxidative Stress




Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Hs68 Cell Culture
2.2.1. Cell Culture and Cytotoxicity
2.2.2. Measurement of Intracellular Reactive Oxygen Species Production
2.2.3. Measurement of Reduced Glutathione and Lipid Peroxidation
2.2.4. Measurement of Matrix Metalloproteinase-1 and Matrix Metalloproteinase-3
2.2.5. Measurement of Collagen Contents
2.3. HepG2 Cell Culture
2.3.1. Cell Culture and Cytotoxicity
2.3.2. Measurement of Intracellular Reactive Oxygen Species Production
2.3.3. Measurement of Hepatic Alanine Aminotransferase and Aspartate Aminotransferase Activities
2.4. EA.hy926 Cell Culture
2.4.1. Cell Culture and Cytotoxicity
2.4.2. Measurement of Intracellular Reactive Oxygen Species Production
2.4.3. Measurement of Reduced Glutathione and Lipid Peroxidation
2.4.4. Nitric Oxide Measurement
2.5. RAW264.7 Cell Culture
2.5.1. Cell Culture and Cytotoxicity
2.5.2. Nitric Oxide Assay
2.5.3. Measurement of Tumor Necrosis Factor-α and Interleukin-6
2.6. Statistical Analysis
3. Results and Discussion
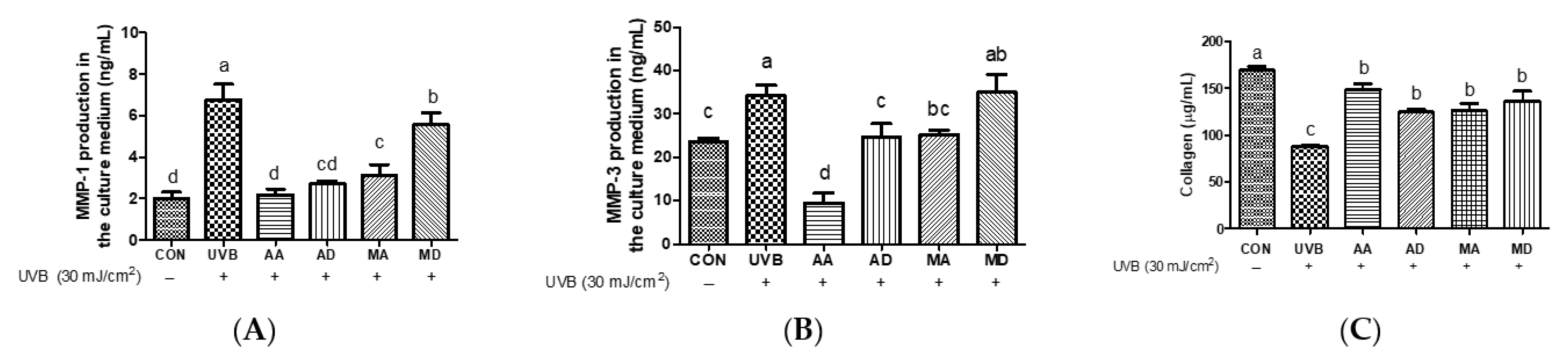
3.1. Effects of Triterpenes on UVB-Exposed Hs68 Cells
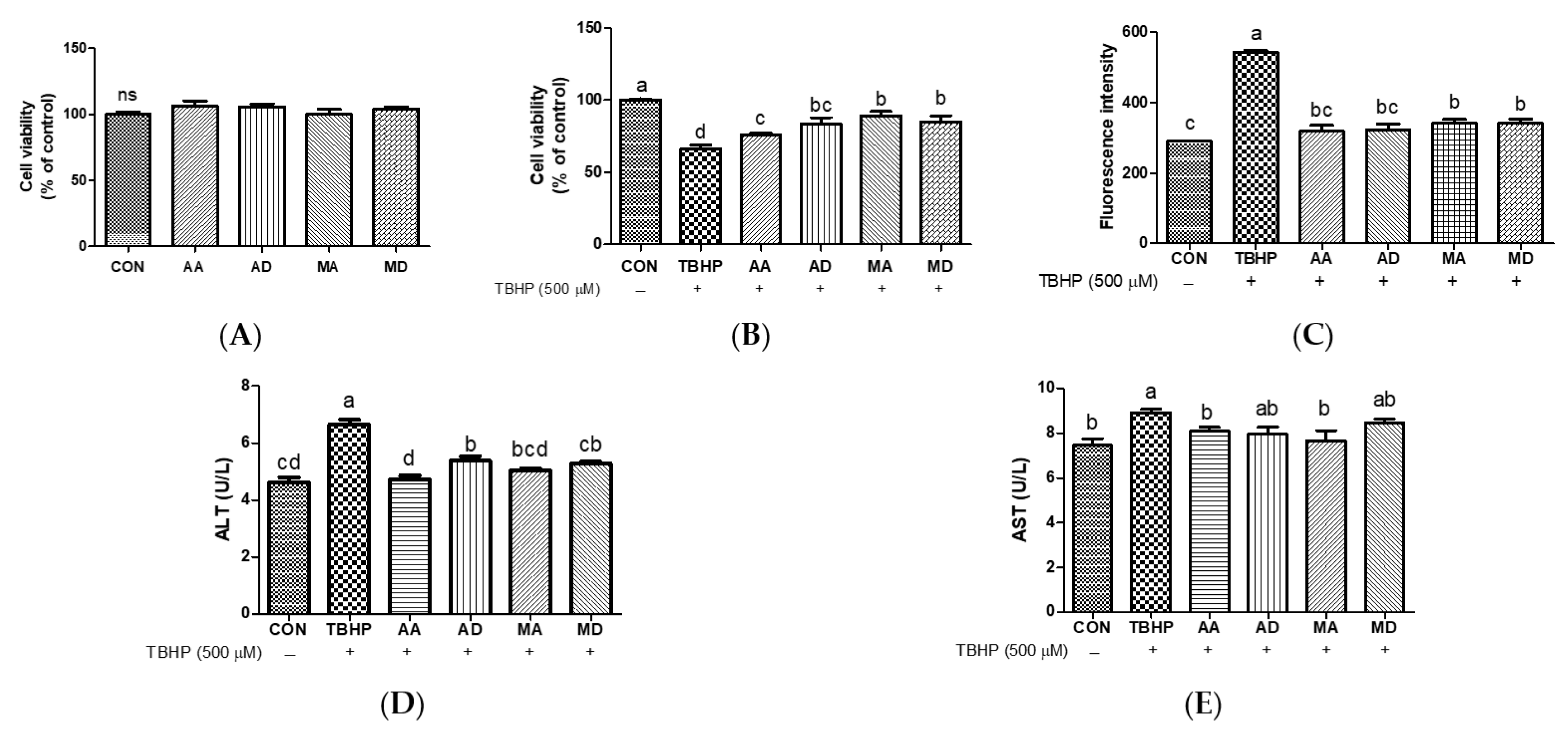
3.2. Effects of Triterpenes against Oxidative Stress in HepG2 Cells
3.3. Effects of Triterpenes on H2O2-Induced EA.hy926 Cells
3.4. Effects of Triterpenes on LPS-Induced RAW264.7 Cells
3.5. Principal Component Analysis (PCA) and Hierarchical Clustering Analysis (HCA) of Triterpenes Based on Biological Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brinkhaus, B.; Lindner, M.; Schuppan, D.; Hahn, E.G. Chemical, pharmacological and clinical profile of the East Asian medical plant Centella asiatica. Phytomedicine 2000, 7, 427–448. [Google Scholar] [CrossRef] [PubMed]
- James, J.T.; Dubery, I.A. Pentacyclic triterpenoids from the medicinal herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef]
- Sun, B.; Wu, L.; Wu, Y.; Zhang, C.; Qin, L.; Hayashi, M.; Kudo, M.; Gao, M.; Liu, T. Therapeutic Potential of Centella asiatica and Its Triterpenes: A Review. Front. Pharmacol. 2020, 11, 568032. [Google Scholar] [CrossRef] [PubMed]
- Zainol, M.K.; Abd-Hamid, A.; Yusof, S.; Muse, R. Antioxidative activity and total phenolic compounds of leaf, root and petiole of four accessions of Centella asiatica (L.) Urban. Food Chem. 2003, 81, 575–581. [Google Scholar] [CrossRef]
- Anukunwithaya, T.; Tantisira, M.H.; Tantisira, B.; Khemawoot, P. Pharmacokinetics of a Standardized Extract of Centella asiatica ECa 233 in Rats. Planta Med. 2017, 83, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Hashim, P.; Sidek, H.; Helan, M.H.; Sabery, A.; Palanisamy, U.D.; Ilham, M. Triterpene composition and bioactivities of Centella asiatica. Molecules 2011, 16, 1310–1322. [Google Scholar] [CrossRef] [PubMed]
- Kulbacka, J.; Saczko, J.; Chwilkowska, A. Oxidative stress in cells damage processes. Pol. Merkur. Lekarski. 2009, 27, 44–47. [Google Scholar] [PubMed]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [PubMed]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012, 936486. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Baker, M.A.; Cerniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]
- Yu, J.; Piao, B.K.; Pei, Y.X.; Qi, X.; Hua, B.J. Protective effects of tetrahydropalmatine against gamma-radiation induced damage to human endothelial cells. Life Sci. 2010, 87, 55–63. [Google Scholar] [CrossRef]
- An, I.S.; An, S.; Kang, S.M.; Choe, T.B.; Lee, S.N.; Jang, H.H.; Bae, S. Titrated extract of Centella asiatica provides a UVB protective effect by altering microRNA expression profiles in human dermal fibroblasts. Int. J. Mol. Med. 2012, 30, 1194–1202. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.H.; Kim, S.Y. Resveratrol-Enriched Rice Attenuates UVB-ROS-Induced Skin Aging via Downregulation of Inflammatory Cascades. Oxid Med. Cell Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef] [PubMed]
- Na, M.; Min, B.S.; An, R.B.; Song, K.S.; Seong, Y.H.; Bae, K. Effect of Astilbe koreana on ultraviolet B (UVB)-induced inflammatory response in human keratinocytes. Biol. Pharm. Bull. 2004, 27, 1301–1304. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, K.S.; Han, C.S.; Yang, H.C.; Park, S.H.; Ko, K.I.; Lee, S.H.; Kim, K.H.; Lee, N.H.; Kim, J.M.; et al. Inhibitory effects of natural plants of Jeju Island on elastase and MMP-1 expression. J. Cosmet. Sci. 2007, 58, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Brenneisen, P.; Wenk, J.; Klotz, L.O.; Wlaschek, M.; Briviba, K.; Krieg, T.; Sies, H.; Scharffetter-Kochanek, K. Central role of Ferrous/Ferric iron in the ultraviolet B irradiation-mediated signaling pathway leading to increased interstitial collagenase (matrix-degrading metalloprotease (MMP)-1) and stromelysin-1 (MMP-3) mRNA levels in cultured human dermal fibroblasts. J. Biol. Chem. 1998, 273, 5279–5287. [Google Scholar] [CrossRef]
- Quan, T.; He, T.; Voorhees, J.J.; Fisher, G.J. Ultraviolet irradiation induces Smad7 via induction of transcription factor AP-1 in human skin fibroblasts. J. Biol. Chem. 2005, 280, 8079–8085. [Google Scholar] [CrossRef]
- Nema, N.K.; Maity, N.; Sarkar, B.K.; Mukherjee, P.K. Matrix metalloproteinase, hyaluronidase and elastase inhibitory potential of standardized extract of Centella asiatica. Pharm. Biol. 2013, 51, 1182–1187. [Google Scholar] [CrossRef]
- Yingngam, B.; Chiangsom, A.; Brantner, A. Modeling and optimization of microwave-assisted extraction of pentacyclic triterpenes from Centella asiatica leaves using response surface methodology. Ind. Crops Prod. 2020, 147, 112231. [Google Scholar] [CrossRef]
- Lee, J.; Jung, E.; Kim, Y.; Park, J.; Park, J.; Hong, S.; Kim, J.; Hyun, C.; Kim, Y.S.; Park, D. Asiaticoside induces human collagen I synthesis through TGFbeta receptor I kinase (TbetaRI kinase)-independent Smad signaling. Planta Med. 2006, 72, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Quan, T.; Purohit, T.; Shao, Y.; Cho, M.K.; He, T.; Varani, J.; Kang, S.; Voorhees, J.J. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am. J. Pathol. 2009, 174, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.O.; Kim, J.H.; Che, D.N.; Kang, H.J.; Shin, J.Y.; Hao, S.; Park, J.H.; Wang, F.; Lee, Y.J.; Jang, S.I.; et al. Prevents Tert-Butyl Hydroperoxide and Acetaminophen-Induced Liver Injury. Molecules 2021, 26, 1635. [Google Scholar] [CrossRef] [PubMed]
- Benatti, U.; Morelli, A.; Damiani, G.; De Flora, A. A methemoglobin dependent and plasma-stimulated experimental model of oxidative hemolysis. Biochem. Biophys. Res. Commun. 1982, 106, 183–1190. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K. A toxicologist guide to the diagnostic interpretation of hepatic biochemical parameters. Food Chem. Toxicol. 2007, 45, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Zheng, H.M.; Kim, J.M.; Lee, K.W.; Park, Y.H.; Lee, D.H. Protective effects of Centella asiatica leaf extract on dimethylnitrosamine-induced liver injury in rats. Mol. Med. Rep. 2016, 14, 4521–4528. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, C.; Gai, R.; Qiu, H.; Wu, Y.; He, Q.; Yang, X. Protective effects and possible mechanisms of Centella asiatica (L.) urban extract against acute and chronic liver injury: Evidence from in vivo and in vitro studies. Phytother. Res. 2021, 35, 2785–2796. [Google Scholar] [CrossRef]
- Yu, S.W. Mediation of poly(ADP-ribose) polymerase-1-dependent cell death by apoptosis-inducing factor. Science 2002, 297, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, H.; Sung, J.; Kim, Y.; Jeong, H.S.; Lee, J. Conversion of Rutin to Quercetin by Acid Treatment in Relation to Biological Activities. Prev. Nutr. Food Sci. 2019, 24, 313–320. [Google Scholar] [CrossRef]
- Han, R.; Tang, F.; Lu, M.; Xu, C.; Hu, J.; Mei, M.; Wang, H. Astragalus polysaccharide ameliorates H2O2-induced human umbilical vein endothelial cell injury. Mol. Med. Rep. 2017, 15, 4027–4034. [Google Scholar] [CrossRef]
- Bian, D.; Liu, M.; Li, Y.; Xia, Y.; Gong, Z.; Dai, Y. Madecassoside, a triterpenoid saponin isolated from Centella asiatica herbs, protects endothelial cells against oxidative stress. J. Biochem. Mol. Toxicol. 2012, 26, 399–406. [Google Scholar] [CrossRef]
- Cardounel, A.J.; Cui, H.; Samouilov, A.; Johnson, W.; Kearns, P.; Tsai, A.L.; Berka, V.; Zweier, J.L. Evidence for the pathophysiological role of endogenous methylarginines in regulation of endothelial NO production and vascular function. J. Biol. Chem. 2007, 282, 879–887. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Wang, W.; Jin, Y.; Chen, M.; Huang, X.; Zhu, X.; Wang, L. Effect of asiaticoside on endothelial cells in hypoxia-induced pulmonary hypertension. Mol. Med. Rep. 2018, 17, 2893–2900. [Google Scholar] [CrossRef] [PubMed]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Uwai, K.; Osanai, Y.; Imaizumi, T.; Kanno, S.; Takeshita, M.; Ishikawa, M. Inhibitory effect of the alkyl side chain of caffeic acid analogues on lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophages. Bioorg. Med. Chem. 2008, 16, 7795–7803. [Google Scholar] [CrossRef] [PubMed]
- Puttarak, P.; Brantner, A.; Panichayupakaranant, P. Biological Activities and Stability of a Standardized Pentacyclic Triterpene Enriched Centella asiatica Extract. Nat. Prod. Sci. 2016, 22, 20. [Google Scholar] [CrossRef]
- Yun, K.J.; Kim, J.Y.; Kim, J.B.; Lee, K.W.; Jeong, S.Y.; Park, H.J.; Jung, H.J.; Cho, Y.W.; Yun, K.; Lee, K.T. Inhibition of LPS-induced NO and PGE2 production by asiatic acid via NF-kappa B inactivation in RAW 264.7 macrophages: Possible involvement of the IKK and MAPK pathways. Int. Immunopharmacol. 2008, 8, 431–441. [Google Scholar] [CrossRef]
- He, Y.; Sun, X.; Huang, C.; Long, X.R.; Lin, X.; Zhang, L.; Lv, X.W.; Li, J. MiR-146a regulates IL-6 production in lipopolysaccharide-induced RAW264.7 macrophage cells by inhibiting Notch1. Inflammation 2014, 37, 71–82. [Google Scholar] [CrossRef]
- Tang, S.; Xie, X.; Wang, M.; Yang, L.; Wei, W. Protective effects of asiaticoside on renal ischemia reperfusion injury in vivo and in vitro. Bioengineered 2022, 13, 10235–10243. [Google Scholar] [CrossRef]
- Granato, D.; Santos, J.S.; Escher, G.B.; Ferreira, B.L.; Maggio, R.M. Use of principal component analysis (PCA) and hierarchical cluster analysis (HCA) for multivariate association between bioactive compounds and functional properties in foods: A critical perspective. Trends Food Sci. Technol. 2018, 72, 83–90. [Google Scholar] [CrossRef]
- Duan, L.; Zhang, C.; Zhao, Y.; Chang, Y.; Guo, L. Comparison of Bioactive Phenolic Compounds and Antioxidant Activities of Different Parts of Taraxacum mongolicum. Molecules 2020, 25, 260. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PC 1 | PC 2 | PC 3 | PC 4 | |
|---|---|---|---|---|
| AA | 0.75 | −0.58 | −0.33 | 0.00 |
| AD | 0.36 | 0.10 | 0.64 | 0.67 |
| MA | 0.41 | 0.25 | 0.49 | −0.72 |
| MD | 0.38 | 0.77 | −0.49 | 0.15 |
| Eigenvalue | 0.89 | 0.14 | 0.04 | 0.00 |
| Variability (%) | 82.83 | 12.89 | 3.88 | 0.04 |
| Cumulative variability (%) | 82.83 | 95.72 | 99.60 | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.; Lee, H.; Hong, S.; Lee, J.; Kim, Y. Comparison of the Antioxidant Potency of Four Triterpenes of Centella asiatica against Oxidative Stress. Antioxidants 2024, 13, 483. https://doi.org/10.3390/antiox13040483
Lim J, Lee H, Hong S, Lee J, Kim Y. Comparison of the Antioxidant Potency of Four Triterpenes of Centella asiatica against Oxidative Stress. Antioxidants. 2024; 13(4):483. https://doi.org/10.3390/antiox13040483
Chicago/Turabian StyleLim, Jinyeong, Hana Lee, Seonghwa Hong, Junsoo Lee, and Younghwa Kim. 2024. "Comparison of the Antioxidant Potency of Four Triterpenes of Centella asiatica against Oxidative Stress" Antioxidants 13, no. 4: 483. https://doi.org/10.3390/antiox13040483
APA StyleLim, J., Lee, H., Hong, S., Lee, J., & Kim, Y. (2024). Comparison of the Antioxidant Potency of Four Triterpenes of Centella asiatica against Oxidative Stress. Antioxidants, 13(4), 483. https://doi.org/10.3390/antiox13040483