

Potential of Natural Phenolic Compounds against Doxorubicin-Induced Chemobrain: Biological and Molecular Mechanisms Involved


Abstract
1. Introduction
2. DOX: Chemotherapeutic Activity and Mechanisms Underlying Its Toxic Effect at Brain Level
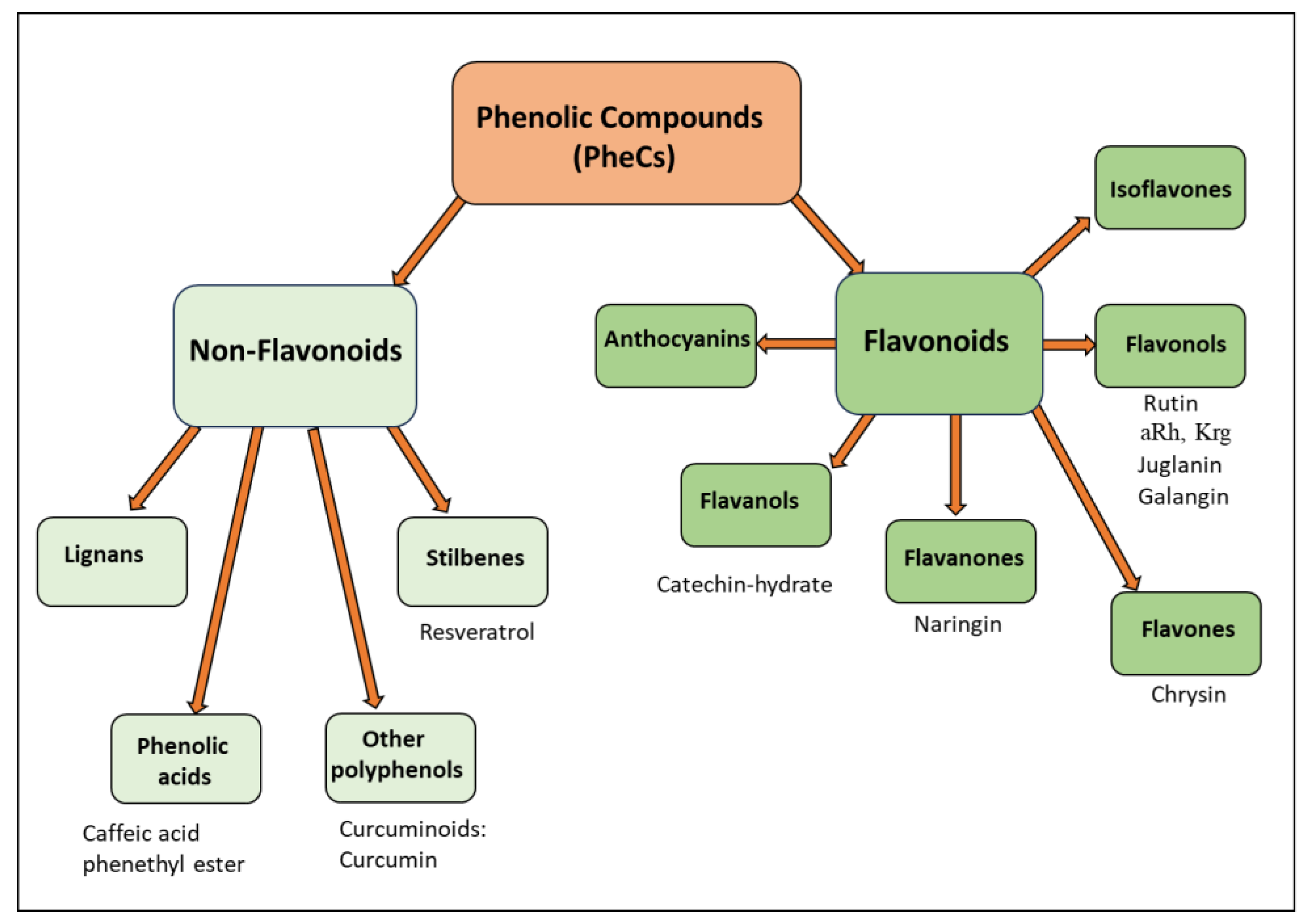
3. PheCs: Natural Products with Antioxidant, Anti-Inflammatory and Antineoplastic Activities
3.1. Effects of PheCs on DOX-Induced Chemobrain

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anthracycline-Induced Chemobrain Model | Anthracyclin Administered | PheC Administered | Protective Effect(s) of the Combination with Respect to the Neurotoxic Effect of DOX Alone | Molecular Factors/Mechanisms Involved in the Protective Effects of PheCs against DOX-Induced Effects | Ref. |
|---|---|---|---|---|---|
| (A) | |||||
| In vitro: human neuroblastoma (IMR32) cells. In vivo: twelve-week-old, healthy female Wistar rats, weighing 180–230 g. | In vitro: 1 μM DOX In vivo: ten cycles with i.p. injection of DOX: 2.5 mg/kg, every 5 days for 50 days. | Rutin (RUT, Flavonol glucoside) In vitro: 100 μM RUT prior to DOX In vivo: RUT: 50 mg/kg (82 μmol/kg), starting one week before first DOX cycle and continuing for further 50 days, per os | In vivo: ↓ Impairment of episodic memory measured by ORT In vitro: ↓ DOX-induced IMR32 cell death, apoptosis, and intracellular ROS generation Restored neurite growth in differentiated IMR32 cells | In vitro: ↓ Cell death (apoptosis) ↓ Intracellular ROS generation ↑ Neurite growth In vivo: In the HIP and frontal cortex: ↓ TNF-α levels ↑ CAT, GSH, total thiols, and SOD levels | [53] |
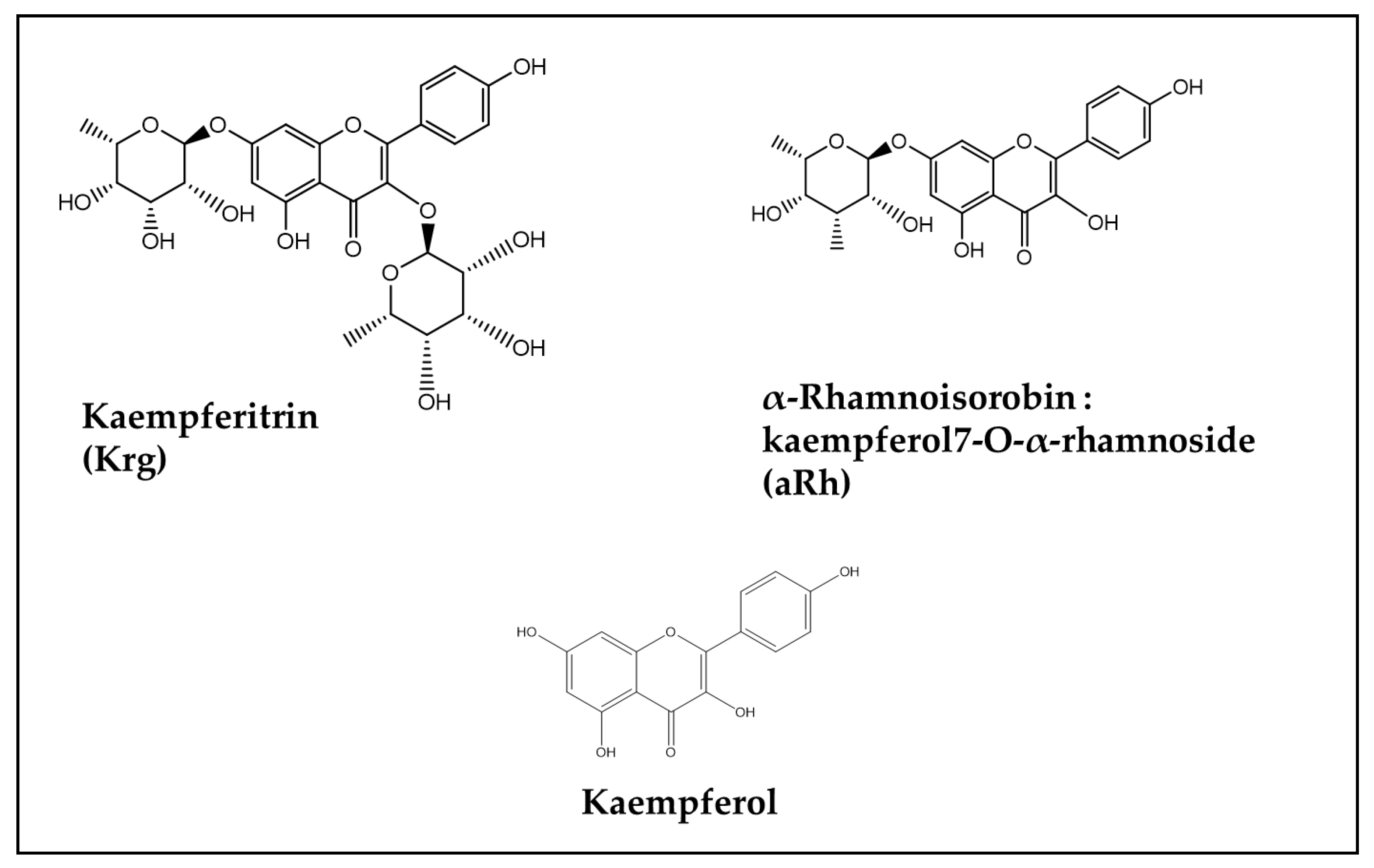
| In vitro: undifferentiated and retinoic acid-differentiated SH-SY5Y neuroblastoma cells | DOX (0.375 and 0.5 μM for UN- and RA-SH-SY5Y, respectively). | Isoquercetin (5–50 μM) used as a reference compound. Kaempferol precursors (Flavonol glycosides) extracted from the leaves of Maesa membranacea (5–50 μM): α-Rhamnoisorobin (Kaempferol 7-O-α-rhamnoside, aRh) and Kaempferitrin (Kaempferol 3,7-di-O-rhamnoside, Krg) | In vitro: No protection by any Flavonoid against DOX-induced cytotoxicity in both UN-SH-SY5Y and RA-SH-SY5Y cells (measured as cell viability modification by WST-1 assay) | Not applicable | [88] |
| In vivo: adult male (8–10 weeks) Wistar rats (180 ± 25 g) | DOX (2 mg/kg, once/week for 4 weeks, i.p.) | Galangin, 50 mg/kg (185 µmol/kg), 5 times/week, for 4 weeks, per os | ↓ Hippocampal neurodegeneration Cognitive and behavioral functions improved HIP antioxidant status ameliorated ↓ HIP DOX-induced inflammation reduced ↓ HIP long-standing and deleterious DOX-induced astrocyte activation | In the HIP: ↑ BDNF expression; ↓ Levels of oxidative indexes (MDA, NO, NOX-1) ↑ NRF2 and HO-1 expression; ↑ GSH tissue level ↓ Expression of inflammatory biomarkers: NF-κB p65, iNOS, TNF-α, IL-6, and IL-1β Modified expression of markers of astrocyte activation: ↑ GFAP expression and ↓ BDNF expression ↓ Expression of necroptosis markers: p-RIPK1, p-RIPK3, and p-MLKL | [89] |
| In vivo: six-week-old male Sprague Dawley rats: 180 ± 40 g | 5 mg/kg DOX i.p. administered once a week for 3 weeks (total dose of 15 mg/kg). | Juglanin, 30 mg/kg/day (71 μmol/kg/day) for 4 weeks. For the first week, alone; for the remaining 3 weeks, in combination with the weekly DOX injection, per os. | ↓ DOX-induced increase in immobility time (DOX-induced sign of anxiety) ↑ DOX-induced decrease in swimming and climbing time (DOX-induced depression-like behaviors). Improved learning and memory (measured by the Y-maze test, the Y-maze test, and the MWM test). ↓ DOX-induced neuroinflammation and oxidative stress. ↓ Histopathologic alterations (pyknosis, congested blood vessels, degenerated and swollen neurons) | Measured brain homogenates: ↓ MDA level ↑ SOD, CAT, and GSH levels ↓ AChE levels TNF-α, IL-1β, IL-6 and NF-κB content ↓ Caspase 3 activity | [90] |
| (B) | |||||
| In vivo: healthy male Wistar rats (150–200 g) | DOX (15 mg/kg, i.p.) administered on the 10th day of the Naringin treatment. | Naringin (Flavanone) 50 and 100 mg/kg (86 and 172 μmol/kg) for 14 days, i.p. injected | Alleviation of anxiety-like behavior (time spent in open arms, closed arms) and depressive-like behavior (immobility time, swimming time) | In vivo: In the HIP: ↓ plasma corticosterone, TNF-α and IL-1β levels ↑ mitochondrial complexes I, II activities and mitochondrial redox activity ↑ levels of serotonin and dopamine | [91] |
| In vitro: undifferentiated and RA-differentiated neuroblastoma IMR-32 cells In vivo: twelve-week-old healthy male rats weighing 200–230 g | In vitro: 1 and 2 μg/mL DOX In vivo: 2.5 mg/kg b.w. i.p. injection (10 cycles every 5 days) | Catechin hydrate (Flavanol) In vitro: 31–250 μg/mL In vivo: 100 mg/kg b.w. (324 μmol/kg) for 57 days including one week prior to the first cycle of DOX, per os | In vitro: ↑ undifferentiated cell viability ↓ degeneration, ↑ neurite length and prevention of DOX-induced cell cycle arrest in differentiated cells In vivo: prevention of DOX-induced memory deficit by the NORT assay | In vivo: ↓ oxidative stress, acetylcholine esterase and neuroinflammation (reduced nitrite and MPO levels) in the HIP and cerebral cortex | [92] |
| In vivo: eight-week-old male Sprague Dawley rats 180–200 g | DOX, 2 mg/kg/week, i.p. for 4 weeks. | Free Chrysin (Flavone), 30 mg/kg (118 μmol/kg), per os or Chrysin formulations (transfersomal and chitosan composite vesicles), 0.5 mg/kg, 5 times/week/4 weeks intranasal delivery | ↓ histological changes and neurodegeneration ↑ cholinergic transmission ↑ memory acquisition and spatial memory (y maze test and Moris water maze test) | In HIP and PFC: ↑ GSH levels and CAT activity ↓ level of hydrogen peroxide and lipid peroxidation Inhibition of TLR4/NF-κB/NLRP3 signaling pathway ↓ caspase-1 and IL-1β protein expression ↓ AchE enzyme | [67] |
| Anthracycline-Induced Chemobrain Model | Anthracycline Administered | PheC Administered | Protective Effect(s) of the Combination with Respect to the Neurotoxic Effect of DOX Alone | Molecular Factors/Mechanisms Involved in the Protective Effects of PheCs against DOX-Induced Effects | Ref. |
|---|---|---|---|---|---|
| Sprague Dawley male rats (200–250 g) | DOX (2 mg/kg, once a week, i.p. for 4 weeks). | CAPE, 10 or 20 μmol/kg/day, 5 days per week, for 4 weeks, i.p. injected. | Counteraction of spatial learning and memory impairment (measured by MWM test; passive avoidance test; assessment of locomotion) ↓ Hippocampal and PFC neurodegenerative changes (nuclear pyknosis and degeneration of the neuronal cells) | In the HIP and cortex: ↓ levels of inflammatory biomarkers (GFAP, COX-2, NF-κB p65, TNF-α) Normalization of GSH and MDA levels ↑ ACh levels ↓ Active caspase-3 levels | [94] |
| Male adult Wistar rats (160 to 180 g) | A single dose of DOX (20 mg/kg, i.p.) on the 10th day | Nanocurcumin, 50 mg/kg/day, for 9 days (before the DOX injection) and for further 4 days, per os | Not determined | Normalization of oxidative stress parameters: in the cortex: (↓ MDA and NO levels) in the HIP: (↑ GSH levels) in the cortex and HIP: no effect on the DOX-induced inhibition of AchE and MAO activities normalization of DOX-induced ↑ levels of dopamine | [97] |
| Male Wistar rats, 3 months of age (290 ± 20 g) | DOX, a weekly dose of 2.5 mg/kg for 4 weeks, i.p. | Curcumin, 10 mg/kg/day (27 μmol/kg/day) for 28 days, started on the same day as the first DOX injection, per os | ↓ Short- and long-term memory impairment (NORT test at 3 and 24 h after habituation) | In frontal cortex, hypothalamus and HIP: ↓ GFAP (in astrocytes) and Iba1 (in microglia) expression (markers of strong microglial and astrocyte response, due to the neuroinflammatory response) | [95] |
| Male Sprague Dawley rats (200–220 g) | DOX (2.5 mg/kg injected i.p. every 2 days) | Curcumin, 30 mg/kg/day (81 μmol/kg/day) for 3 weeks starting 1 week before DOX administration, per os | Improvement of DOX-induced symptoms of depression: (↓ sucrose preference in SPT, ↑ latency time in NSFT, immobility time in FST and number of crossings in OFT ↓ histopathological changes (nuclear pyknosis) and apoptosis (measured by Tunel test) | In the HIP: ↓ oxidative stress biomarkers (↓ HNE-positive cells, ↓ levels of MDA and NO, ↓ CAT and GPx activities ↓ Endoplasmic reticulum (ER) stress biomarkers (↓ CHOP and GRP78 expression) Modulation of autophagy biomarkers (↓ LC3-II/LC3-I ratio, ↓ Atg-5, Atg-7, and Becn1, expression; ↑ p62 expression) Activation of NRF2-ARE pathway (↑ NRF2 and Keap-1 nuclear expression; ↑ NQO-1 and HO-1 expression) | [96] |
| Anthracycline-Induced Chemobrain Model | Anthracycline Administered | PheC Administered | Protective Effect(s) of the Combination with Respect to the Neurotoxic Effect of DOX Alone | Molecular Factors/Mechanisms Involved in the Protective Effects of PheCs against DOX-Induced Effects | Ref. |
|---|---|---|---|---|---|
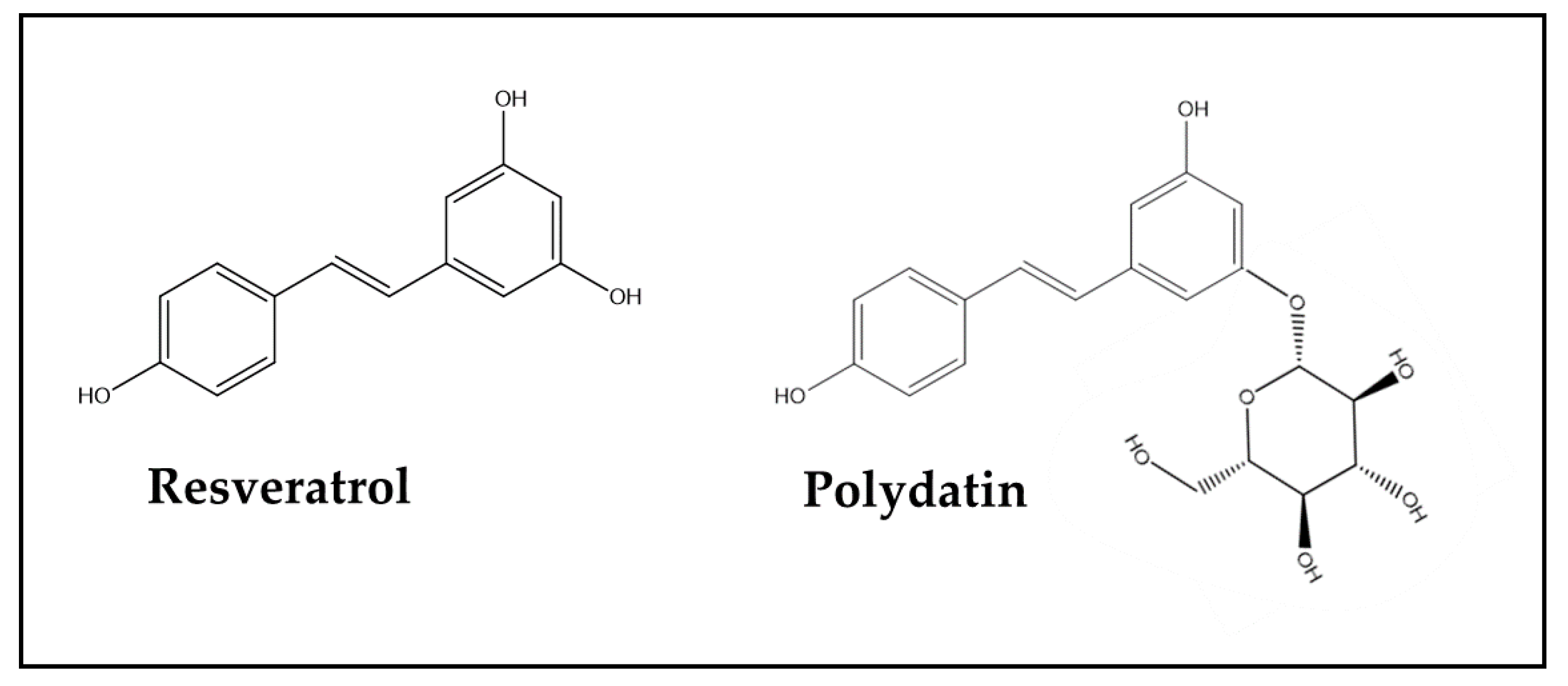
| Female C57/BL6J mice (18–20 g) | DOX in combination with other chemotherapies: DTX + DOX + CP, 10/10/40 mg/kg (DAC) Three i.p. injections at 2-day intervals | Resveratrol, 50 and 100 mg/kg/day (219 or 438 μmol/kg/day) for three weeks, beginning one week before the DAC treatment per os | ↓ Anxiety levels and locomotor activity (open-field test) ↑ Cognitive performance (by Morris water maze test) ↑ PFC and hippocampal neuronal activity (by MEMRI test) | In serum, whole brain, PFC, and HIP: ↓ TNF-α and IL-6 levels ↑ IL-4 and IL-10 levels In PFC and HIP: ↑ Neuroplasticity biomarker expression (BDNF, TrkB, amino acid neurotransmitter receptors, and CaMKII) Regulation of components of PPARγ/NF-κB signaling (↑ PPARγ expression, ↓ p-p65, and p-IκBα expression) | [98] |
| Forty male Wistar rats, 3 months of age and weighing 290 ± 20 g | DOX, 2.5 mg/kg/week for 4 weeks, i.p. | Resveratrol, 10 mg/kg/day (44 μmol/kg/day) for 28 days, started on the same day as the first DOX injection, per os | ↑ Long-term memory impairment (NORT at 24 h after habituation). | In frontal cortex, hypothalamus and HIP: ↓ GFAP (in astrocytes) and Iba1 (in microglia) expression (markers of strong microglial and astrocyte response, due to the neuroinflammatory response) | [95] |
| Six-week-old male Sprague Dawley rats (210–230 g) | DOX, 2 mg/kg, once a week for 4 weeks, i.p. | Polydatin, 50 mg/kg/day (128 μmol/kg/day) for 4 weeks, per os | ↑ Spatial learning and memory ability in rats (Morris water-maze task). Inhibition of nuclear pyknosis and degeneration of neuronal cells in the HIP | In HIP: ↓ Oxidative stress (↓ MDA levels, ↑ GSH levels; ↓ NRF2 expression) ↓ Inflammation (↓ TNF-α, PGE-2 and COX-2 levels; ↓ p-IκB and p-p65 expression) ↓ Apoptosis (↓ cleaved caspase-3 and -9 expression) | [99] |
3.1.1. Flavonoids and DOX-Induced Chemobrain
3.1.2. Non-Flavonoids and DOX-Induced Chemobrain
3.1.3. Extracts of Plants with High Contents of PheCs and DOX-Induced Chemobrain
| Anthracycline-Induced Chemobrain Model | Anthracycline Administered | Administered Sources of PheCs | Protective Effect(s) of the Combination with Respect to the Neurotoxic Effect of DOX Alone or in Combination with CP | Molecular Factors/Mechanisms Involved in the Protective Effects of PheCs against DOX-Induced Effects | Ref. |
|---|---|---|---|---|---|
| Male Wistar rats, weighing 120–160 g | DOX 2.5 mg/kg/week for 4 weeks, i.p. | Carissa macrocarpa leaves polar fraction of hydromethanolic extract containing high % of PheCs (evaluated by UPLC-ESI-MS/MS profiling) 100–500 mg/kg/day Carissa m. extract for 4 weeks, per os | ↓ Short- and long-term memory impairments (measured by Y-maze test and NORT, respectively) in a dose-dependent manner Partial recovery from histopathological alterations in the HIP (↓ perivascular space; ↓ neuropil vacuolation; ↓ neuroglia cell pyknosis and hyperchromasia; ↓ pyramidal cell degeneration) | ↓ ROS and TNF-α serum levels ↓ caspase-3 expression in the HIP ↑ NGF brain levels in a dose-dependent manner | [158] |
| Male Wistar rats (180–200 g, 8 weeks old) | DOX (4 mg/kg/week, i.v.) in combination with CP, (40 mg/kg/week, i.v.) in the rats’ tail vein once per week for 3 weeks | Thunbergia erecta leaf ethyl acetate fraction of alcohol extract (TEAF), (50, 100, or 200 mg/kg) containing 21 characterized phytoconstituents, mostly PheCs (especially Phenolic Acids and Flavonoid glycosides), 5 times per week for 3 weeks, per os | ↓ Learning and memory impairments (measured by NORT, Morris water maze tests, and step-through passive avoidance test) In PFC and HIP: ↓ Histological alterations in a dose-dependent manner Normalization of oxidative stress markers (CAT activity; MDA, GSH, hydrogen peroxide level) | ↓ Protein expression of inflammation-related factors (HMBG1, RAGE, p65 NF-κB, and IL-1β) | [159] |
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ach | acetylcholine |
| AchE | acetylcholinesterase |
| ARE | antioxidant response element |
| ATG | autophagy-related protein |
| BDNF | brain-derived neurotropic factor |
| Becn1 | beclin 1 |
| CaMKII | calmodulin-dependent protein kinase II |
| CAPE | caffeic acid phenethyl ester |
| CAT | catalase |
| CHOP | C/EBP homologous protein |
| COX | cyclooxygenase |
| CP | cyclophosphamide |
| CUR | Curcumin |
| DTX | docetaxel |
| FST | forced swimming test |
| GFAP | anti-glial fibrillary acidic protein |
| GPx | glutathione peroxidase |
| GRP78 | glucose-regulated protein 78 |
| GSH | reduced glutathione |
| HIP | hippocampus |
| HMBG1 | High-Mobility Group Box 1 protein |
| HNE | 4-hydroxynonenal |
| HO-1 | heme oxygenase-1 |
| Iba1 | ionized calcium-binding adaptor molecule 1 |
| IκB | inhibitor of the nuclear factor κ |
| iNOS | inducible nitric oxide synthase |
| MAO | monoamine oxidase |
| MDA | malondialdehyde |
| MEMRI | manganese-enhanced magnetic resonance imaging |
| MLKL | mixed-lineage kinase domain-like protein |
| MPO | mieloperoxidase |
| NF-κB | nuclear factor κB |
| NGF | nerve growth factor |
| NLRP3 | Nod-like receptor pyrin-containing 3 |
| NO | nitric oxide |
| NORT | novel object recognition test |
| NOX | NADPH oxidase-1 |
| NQO-1 | NAD(P)H quinone oxidoreductase |
| NRF2 | nuclear factor (erytheroid-derived-2)-like 2 |
| NSFT | novelty-suppressed feeding test |
| OFT | open-field test |
| PFC | prefrontal cortex |
| PGE2 | prostaglandin E2 |
| PPARγ | peroxisome proliferator-activated receptor |
| RA | retinoic acid |
| RAGE | Receptor for Advanced Glycation End-products |
| RIPK | receptor-interacting serine/threonine protein kinase |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| SPT | sucrose preference test |
| TLR4 | Toll-like receptor 4 |
| TrkB | tropomyosin receptor kinase B |
| UPLC-ESI-MS/MS | ultra-performance liquid chromatography–electrospray ionization–tandem mass spectrometry. |
References
- Kuderer, N.M.; Desai, A.; Lustberg, M.B.; Lyman, G.H. Mitigating acute chemotherapy-associated adverse events in patients with cancer. Nat. Rev. Clin. Oncol. 2022, 19, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Wefel, J.S.; Saleeba, A.K.; Buzdar, A.U.; Meyers, C.A. Acute and late onset cognitive dysfunction associated with chemotherapy in women with breast cancer. Cancer 2010, 116, 3348–3356. [Google Scholar] [CrossRef] [PubMed]
- Janelsins, M.C.; Kesler, S.R.; Ahles, T.A.; Morrow, G.R. Prevalence, mechanisms, and management of cancer-related cognitive impairment. Int. Rev. Psychiatry 2014, 26, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Pinho, R.; Correia, V.; Soares, J.; Bastos, M.L.; Carvalho, F.; Capela, J.P.; Costa, V.M. Mitoxantrone is More Toxic than Doxorubicin in SH-SY5Y Human Cells: A ‘Chemobrain’ In Vitro Study. Pharmaceuticals 2018, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, S.E.; Abdel-Aziz, A.K.; Esmat, A.; Azab, S.S. Chemotherapy and cognition: Comprehensive review on doxorubicin-induced chemobrain. Cancer Chemother. Pharmacol. 2019, 84, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dias-Carvalho, A.; Ferreira, M.; Ferreira, R.; Bastos, M.L.; Sá, S.I.; Capela, J.P.; Carvalho, F.; Costa, V.M. Four decades of chemotherapy-induced cognitive dysfunction: Comprehensive review of clinical, animal and in vitro studies, and insights of key initiating events. Arch. Toxicol. 2022, 96, 11–78. [Google Scholar] [CrossRef]
- Jaiswara, P.K.; Shukla, S.K. Chemotherapy-Mediated Neuronal Aberration. Pharmaceuticals 2023, 16, 1165. [Google Scholar] [CrossRef] [PubMed]
- Onzi, G.R.; D’Agustini, N.; Garcia, S.C.; Guterres, S.S.; Pohlmann, P.R.; Rosa, D.D.; Pohlmann, A.R. Chemobrain in Breast Cancer: Mechanisms, Clinical Manifestations, and Potential Interventions. Drug. Saf. 2022, 45, 601–621. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, X.; Zhong, T.; Zhou, M.; Gao, L.; Chen, L. Prevalence and associated factors of chemotherapy-related cognitive impairment in older breast cancer survivors. J. Adv. Nurs. 2024, 80, 484–499. [Google Scholar] [CrossRef]
- Vega, J.N.; Dumas, J.; Newhouse, P. Cognitive effects of chemotherapy and cancer-related treatments in older adults. Am. J. Geriatr. Psychiatry 2017, 25, 1415–1426. [Google Scholar] [CrossRef]
- Asher, A.; Myers, J.S. The effect of cancer treatment on cognitive function. Clin. Adv. Hematol. Oncol. 2015, 13, 441–450. [Google Scholar] [PubMed]
- Haywood, D.; Wallace, I.N.; Lawrence, B.; Baughman, F.D.; Dauer, E.; O’Connor, M. Oncology healthcare professionals’ perceptions and experiences of ‘chemobrain’ in cancer survivors and persons undergoing cancer treatment. Gen. Hosp. Psychiatry 2023, 84, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, A.; Ahles, T.; Chen, B.T.; Mandelblatt, J.; Janelsins, M.C. Cognitive Function in Older Adults with Cancer: Assessment, Management, and Research Opportunities. J. Clin. Oncol. 2021, 39, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Wefel, J.S.; Lenzi, R.; Theriault, R.L.; Davis, R.N.; Meyers, C.A. The cognitive sequelae of standard-dose adjuvant chemotherapy in women with breast carcinoma: Results of a prospective, randomized, longitudinal trial. Cancer 2004, 100, 2292–2299. [Google Scholar] [CrossRef] [PubMed]
- Schagen, S.B.; Wefel, J.S. Chemotherapy-related changes in cognitive functioning. EJC Suppl. 2013, 11, 225–232. [Google Scholar] [CrossRef]
- Lal, R.; Dharavath, R.N.; Chopra, K. Nrf2 Signaling Pathway: A Potential Therapeutic Target in Combating Oxidative Stress and Neurotoxicity in Chemotherapy-Induced Cognitive Impairment. Mol. Neurobiol. 2024, 61, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.A.; Tang, J.J.; Yoo, K.H.; Corujo-Ramirez, A.; Oliveros, A.; Kim, S.H.; Ullah, F.; Altawell, R.; Hawse, J.R.; Cole, P.D.; et al. The selective cyclooxygenase-2 inhibitor NS398 ameliorates cisplatin-induced impairments in mitochondrial and cognitive function. Front. Mol. Neurosci. 2023, 16, 1295991. [Google Scholar] [CrossRef] [PubMed]
- Rabie, O.; El-Nashar, H.A.S.; George, M.Y.; Majrashi, T.A.; Al-Warhi, T.; Hassan, F.E.; Eldehna, W.M.; Mostafa, N.M. Phytochemical profiling and neuroprotective activity of Callistemon subulatus leaves against cyclophosphamide-induced chemobrain. Biomed. Pharmacother. 2023, 167, 115596. [Google Scholar] [CrossRef]
- Dias-Carvalho, A.; Margarida-Araújo, A.; Reis-Mendes, A.; Sequeira, C.O.; Pereira, S.A.; Guedes de Pinho, P.; Carvalho, F.; Sá, S.I.; Fernandes, E.; Costa, V.M. A Clinically Relevant Dosage of Mitoxantrone Disrupts the Glutathione and Lipid Metabolic Pathways of the CD-1 Mice Brain: A Metabolomics Study. Int. J. Mol. Sci. 2023, 24, 13126. [Google Scholar] [CrossRef]
- Alotayk, L.I.; Aldubayan, M.A.; Alenezi, S.K.; Anwar, M.J.; Alhowail, A.H. Comparative evaluation of doxorubicin, cyclophosphamide, 5-fluorouracil, and cisplatin on cognitive dysfunction in rats: Delineating the role of inflammation of hippocampal neurons and hypothyroidism. Biomed. Pharmacother. 2023, 165, 115245. [Google Scholar] [CrossRef]
- Oliveros, A.; Yoo, K.H.; Rashid, M.A.; Corujo-Ramirez, A.; Hur, B.; Sung, J.; Liu, Y.; Hawse, J.R.; Choi, D.S.; Boison, D.; et al. Adenosine A2A receptor blockade prevents cisplatin-induced impairments in neurogenesis and cognitive function. Proc. Natl. Acad. Sci. USA 2022, 119, e2206415119. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Zhao, S.; Liu, J.X.; Liu, X.; Guo, Y.X.; Wang, G.Y.; Wang, X.L. Paclitaxel induces cognitive impairment via necroptosis, decreased synaptic plasticity and M1 polarisation of microglia. Pharm. Biol. 2022, 60, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Eide, S.; Feng, Z.P. Doxorubicin chemotherapy-induced “chemo-brain”: Meta-analysis. Eur. J. Pharmacol. 2020, 881, 173078. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Mujwar, S.; Kołat, D.; Kałuzińska-Kołat, Ż.; Celik, I.; Kontek, R. Doxorubicin-An Agent with Multiple Mechanisms of Anticancer Activity. Cells 2023, 12, 659. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cui, X.; Zhong, Y.; Ma, R.; Liu, B.; Xia, Y. Phenolic metabolites as therapeutic in inflammation and neoplasms: Molecular pathways explaining their efficacy. Pharmacol. Res. 2023, 193, 106812. [Google Scholar] [CrossRef]
- Farghadani, R.; Naidu, R. The anticancer mechanism of action of selected polyphenols in triple-negative breast cancer (TNBC). Biomed. Pharmacother. 2023, 165, 115170. [Google Scholar] [CrossRef] [PubMed]
- Von Heideman, A.; Sandstrom, M.; Csoka, K.; Tholander, B.; Larsson, R.; Bergh, J.; Nygren, P. Evaluation of drug interactions in the established FEC regimen in primary cultures of tumour cells from patients. Ann. Oncol. 2000, 11, 1301–1307. [Google Scholar] [CrossRef]
- Saharkhiz, S.; Zarepour, A.; Nasri, N.; Cordani, M.; Zarrabi, A. A comparison study between doxorubicin and curcumin co-administration and co-loading in a smart niosomal formulation for MCF-7 breast cancer therapy. Eur. J. Pharm Sci. 2023, 191, 106600. [Google Scholar] [CrossRef]
- Kong, D.; Hong, W.; Yu, M.; Li, Y.; Zheng, Y.; Ying, X. Multifunctional Targeting Liposomes of Epirubicin Plus Resveratrol Improved Therapeutic Effect on Brain Gliomas. Int. J. Nanomed. 2022, 17, 1087–1110. [Google Scholar] [CrossRef]
- Gioti, K.; Papachristodoulou, A.; Benaki, D.; Aligiannis, N.; Skaltsounis, A.L.; Mikros, E.; Tenta, R. Assessment of the Nutraceutical Effects of Oleuropein and the Cytotoxic Effects of Adriamycin, When Administered Alone and in Combination, in MG-63 Human Osteosarcoma Cells. Nutrients 2021, 13, 354. [Google Scholar] [CrossRef]
- Mattioli, R.; Ilari, A.; Colotti, B.; Mosca, L.; Fazi, F.; Colotti, G. Doxorubicin and other anthracyclines in cancers: Activity, chemoresistance and its overcoming. Mol. Asp. Med. 2023, 93, 101205. [Google Scholar] [CrossRef]
- Huang, X.; Ren, Q.; Yang, L.; Cui, D.; Ma, C.; Zheng, Y.; Wu, J. Immunogenic chemotherapy: Great potential for improving response rates. Front. Oncol. 2023, 13, 1308681. [Google Scholar] [CrossRef]
- He, Z.; Feng, D.; Zhang, C.; Chen, Z.; Wang, H.; Hou, J.; Li, S.; Wei, X. Recent strategies for evoking immunogenic Pyroptosis in antitumor immunotherapy. J. Control. Release 2024, 366, 375–394. [Google Scholar] [CrossRef]
- You, X.; Zhu, C.; Yu, P.; Wang, X.; Wang, Y.; Wang, J.; Yu, J.; Wang, K. Emerging strategy for the treatment of urothelial carcinoma: Advances in antibody-drug conjugates combination therapy. Biomed. Pharmacother. 2024, 171, 16152. [Google Scholar] [CrossRef]
- Mei, T.; Wang, T.; Lei, C.; Jiang, D.; Zhou, Q. Pathological complete response to neoadjuvant therapy with serplulimab and chemotherapy in stage IIIB small cell lung cancer: A case report and literature review. Front. Immunol. 2024, 14, 1272450. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, A.; Li, J.; Liu, X.; Wu, S.; Wang, B.; Wang, Y.; Jia, H. Doxorubicin-Induced Cognitive Impairment: The Mechanistic Insights. Front. Oncol. 2021, 11, 673340. [Google Scholar] [CrossRef] [PubMed]
- Arcamone, F.; Cassinelli, G.; Fantini, G.; Grein, A.; Orezzi, P.; Pol, C.; Spalla, C. Adriamycin, 14- hydroxydaunomycin, a new antitumor antibiotic from S. peucetius var. caesius. Biotechnol. Bioeng. 1969, 11, 1101–1110. [Google Scholar] [CrossRef]
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The Good, the Bad and the Ugly Effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An Update on Anticancer Molecular Action, Toxicity and Novel Drug Delivery Systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Van der Zanden, S.Y.; Qiao, X.; Neefjes, J. New insights into the activities and toxicities of the old anticancer drug doxorubicin. FEBS J. 2021, 288, 6095–6111. [Google Scholar] [CrossRef] [PubMed]
- Voorwerk, L.; Slagter, M.; Horlings, H.M.; Sikorska, K.; van de Vijver, K.K.; de Maaker, M.; Nederlof, I.; Kluin, R.J.C.; Warren, S.; Ong, S.; et al. Immune induction strategies in metastatic triple-negative breast cancer to enhance the sensitivity to PD-1 blockade: The TONIC trial. Nat. Med. 2019, 25, 920–928. [Google Scholar] [CrossRef]
- Gilad, Y.; Eliaz, Y.; Yu, Y.; Han, S.J.; O’Malley, B.W.; Lonard, D.M. Drug-induced PD-L1 expression and cell stress response in breast cancer cells can be balanced by drug combination. Sci. Rep. 2019, 9, 15099. [Google Scholar] [CrossRef]
- Kciuk, M.; Kołat, D.; Kałuzińska-Kołat, Ż.; Gawrysiak, M.; Drozda, R.; Celik, I.; Kontek, R. PD-1/PD-L1 and DNA Damage Response in Cancer. Cells 2023, 12, 530. [Google Scholar] [CrossRef]
- Rawat, P.S.; Jaiswal, A.; Khurana, A.; Bhatti, J.S.; Navik, U. Doxorubicin-Induced Cardiotoxicity: An Update on the Molecular Mechanism and Novel Therapeutic Strategies for Effective Management. Biomed. Pharmacother. 2021, 139, 111708. [Google Scholar] [CrossRef]
- Prasanna, P.L.; Renu, K.; Valsala Gopalakrishnan, A. New Molecular and Biochemical Insights of Doxorubicin-Induced Hepatotoxicity. Life Sci. 2020, 250, 117599. [Google Scholar] [CrossRef] [PubMed]
- El-Moselhy, M.A.; El-Sheikh, A.A. Protective mechanisms of atorvastatin against doxorubicin-induced hepato-renal toxicity. Biomed. Pharmacother. 2014, 68, 101–110. [Google Scholar] [CrossRef]
- Alhowail, A.H.; Bloemer, J.; Majrashi, M.; Pinky, P.D.; Bhattacharya, S.; Yongli, Z.; Bhattacharya, D.; Eggert, M.; Woodie, L.; Buabeid, M.A.; et al. Doxorubicin-induced neurotoxicity is associated with acute alterations in synaptic plasticity, apoptosis, and lipid peroxidation. Toxicol. Mech. Methods 2019, 29, 457–466. [Google Scholar] [CrossRef]
- Barry, R.L.; Byun, N.E.; Tantawy, M.N.; Mackey, C.A.; Wilson, G.H., 3rd; Stark, A.J.; Flom, M.P.; Gee, L.C.; Quarles, C.C. In vivo neuroimaging and behavioral correlates in a rat model of chemotherapy-induced cognitive dysfunction. Brain Imaging Behav. 2018, 12, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Steiniger, S.C.; Kreuter, J.; Khalansky, A.S.; Skidan, I.N.; Bobruskin, A.I.; Smirnova, Z.S.; Severin, S.E.; Uhl, R.; Kock, M.; Geiger, K.D.; et al. Chemotherapy of glioblastoma in rats using doxorubicin-loaded nanoparticles. Int. J. Cancer 2004, 109, 759–767. [Google Scholar] [CrossRef]
- Byeon, H.J.; Thao le, Q.; Lee, S.; Min, S.Y.; Lee, E.S.; Shin, B.S.; Choi, H.G.; Youn, Y.S. Doxorubicin-loaded nanoparticles consisted of cationic- and mannose-modified-albumins for dual-targeting in brain tumors. J. Control. Release 2016, 225, 301–313. [Google Scholar] [CrossRef]
- Manchon, J.F.; Dabaghian, Y.; Uzor, N.E.; Kesler, S.R.; Wefel, J.S.; Tsvetkov, A.S. Levetiracetam mitigates doxorubicin-induced DNA and synaptic damage in neurons. Sci. Rep. 2016, 6, 25705. [Google Scholar] [CrossRef] [PubMed]
- Shokoohinia, Y.; Hosseinzadeh, L.; Moieni-Arya, M.; Mostafaie, A.; Mohammadi-Motlagh, H.R. Osthole attenuates doxorubicin-induced apoptosis in PC12 cells through inhibition of mitochondrial dysfunction and ROS production. Biomed. Res. Int. 2014, 2014, 156848. [Google Scholar] [CrossRef] [PubMed]
- Ramalingayya, G.V.; Cheruku, S.P.; Nayak, P.G.; Kishore, A.; Shenoy, R.; Rao, C.M.; Krishnadas, N. Rutin protects against neuronal damage in vitro and ameliorates doxorubicin-induced memory deficits in vivo in Wistar rats. Drug. Des. Devel. Ther. 2017, 11, 1011–1026. [Google Scholar] [CrossRef] [PubMed]
- Imosemi, I.O.; Owumi, S.E.; Arunsi, U.O. Biochemical and histological alterations of doxorubicin-induced neurotoxicity in rats: Protective role of luteolin. J. Biochem. Mol. Toxicol. 2022, 36, e22962. [Google Scholar] [CrossRef]
- Ongnok, B.; Khuanjing, T.; Chunchai, T.; Pantiya, P.; Kerdphoo, S.; Arunsak, B.; Nawara, W.; Jaiwongkam, T.; Apaijai, N.; Chattipakorn, N.; et al. Donepezil Protects Against Doxorubicin-Induced Chemobrain in Rats via Attenuation of Inflammation and Oxidative Stress without Interfering with Doxorubicin Efficacy. Neurotherapeutics 2021, 18, 2107–2125. [Google Scholar] [CrossRef]
- El-Agamy, S.E.; Abdel-Aziz, A.K.; Wahdan, S.; Esmat, A.; Azab, S.S. Astaxanthin Ameliorates Doxorubicin-Induced Cognitive Impairment (Chemobrain) in Experimental Rat Model: Impact on Oxidative, Inflammatory, and Apoptotic Machineries. Mol. Neurobiol. 2018, 55, 5727–5740. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, W.; Zhao, J.; Sun, W.; Yang, Q.; Chen, C.; Xia, P.; Zhu, J.; Zhou, Y.; Huang, G.; et al. Apigenin ameliorates doxorubicin-induced renal injury via inhibition of oxidative stress and inflammation. Biomed. Pharmacother. 2021, 137, 111308. [Google Scholar] [CrossRef]
- Sangweni, N.F.; Gabuza, K.; Huisamen, B.; Mabasa, L.; van Vuuren, D.; Johnson, R. Molecular insights into the pathophysiology of doxorubicin-induced cardiotoxicity: A graphical representation. Arch. Toxicol. 2022, 96, 1541–1550. [Google Scholar] [CrossRef]
- Gomes, A.; Capela, J.P.; Ribeiro, D.; Freitas, M.; Silva, A.M.; Pinto, D.C.; Santos, C.M.; Cavaleiro, J.A.; Lima, J.L.; Fernandes, E. Inhibition of NF-kB activation and cytokines production in THP-1 monocytes by 2-styrylchromones. Med. Chem. 2015, 11, 560–566. [Google Scholar] [CrossRef]
- Hayslip, J.; Dressler, E.V.; Weiss, H.; Taylor, T.J.; Chambers, M.; Noel, T.; Miriyala, S.; Keeney, J.T.; Ren, X.; Sultana, R.; et al. Plasma TNF-α and Soluble TNF Receptor Levels after Doxorubicin with or without Co-Administration of Mesna-A Randomized, Cross-Over Clinical Study. PLoS ONE 2015, 10, e0124988. [Google Scholar] [CrossRef]
- Tangpong, J.; Cole, M.P.; Sultana, R.; Joshi, G.; Estus, S.; Vore, M.; St Clair, W.; Ratanachaiyavong, S.; St Clair, D.K.; Butterfield, D.A. Adriamycin-induced, TNF-alpha-mediated central nervous system toxicity. Neurobiol. Dis. 2006, 23, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Osburg, B.; Peiser, C.; Dömling, D.; Schomburg, L.; Ko, Y.T.; Voigt, K.; Bickel, U. Effect of endotoxin on expression of TNF receptors and transport of TNF-alpha at the blood-brain barrier of the rat. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E899–E908. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Kastin, A.J. TNFalpha transport across the blood-brain barrier is abolished in receptor knockout mice. Exp. Neurol. 2002, 174, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Hyka, N.; Dayer, J.M.; Modoux, C.; Kohno, T.; Edwards, C.K., 3rd; Roux-Lombard, P.; Burger, D. Apolipoprotein A-I inhibits the production of interleukin-1beta and tumor necrosis factor-alpha by blocking contact-mediated activation of monocytes by T lymphocytes. Blood 2001, 97, 2381–2389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fan, C.; Jiao, H.C.; Zhang, Q.; Jiang, Y.H.; Cui, J.; Liu, Y.; Jiang, Y.H.; Zhang, J.; Yang, M.Q.; et al. Calycosin Alleviates Doxorubicin-Induced Cardiotoxicity and Pyroptosis by Inhibiting NLRP3 Inflammasome Activation. Oxid. Med. Cell Longev. 2022, 2022, 1733834. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tian, C.; Zhang, Z.; Qin, Y.; Meng, R.; Dai, X.; Zhong, Y.; Wei, X.; Zhang, J.; Shen, C. Astragaloside IV Inhibits NLRP3 Inflammasome-Mediated Pyroptosis via Activation of Nrf-2/HO-1 Signaling Pathway and Protects against Doxorubicin-Induced Cardiac Dysfunction. Front. Biosci. 2023, 28, 45. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Abo Elseoud, O.G.; Mohamedy, M.H.; Amer, M.M.; Mohamed, Y.Y.; Elmansy, S.A.; Kadry, M.M.; Attia, A.A.; Fanous, R.A.; Kamel, M.S.; et al. Nose-to-brain delivery of chrysin transfersomal and composite vesicles in doxorubicin-induced cognitive impairment in rats: Insights on formulation, oxidative stress and TLR4/NF-kB/NLRP3 pathways. Neuropharmacology 2021, 197, 108738. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zhou, Y.; Ma, L.; Li, W.; Chan, C.; Zhang, S.; Zhao, Y. Inhibition of NLRP3 alleviated chemotherapy-induced cognitive impairment in rats. Neurosci. Lett. 2023, 793, 136975. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Duan, F.; Hu, J.; Luo, B.; Huang, B.; Lou, X.; Sun, X.; Li, H.; Zhang, X.; Yin, S.; et al. NLRP3 inflammasome-mediated pyroptosis contributes to the pathogenesis of non-ischemic dilated cardiomyopathy. Redox Biol. 2020, 34, 101523. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Aleixandre-Tudo, J.L.; Buica, A.; Nieuwoudt, H.; Aleixandre, J.L.; du Toit, W. Spectrophotometric Analysis of Phenolic Compounds in Grapes and Wines. J. Agric. Food Chem. 2017, 65, 4009–4026. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Cheng, M.Y.; Xun, M.H.; Zhao, Z.W.; Zhang, Y.; Tang, W.; Cheng, J.; Ni, J.; Wang, W. Possible Mechanisms of Oxidative Stress-Induced Skin Cellular Senescence, Inflammation, and Cancer and the Therapeutic Potential of Plant Polyphenols. Int. J. Mol. Sci. 2023, 24, 3755. [Google Scholar] [CrossRef]
- Chimento, A.; De Luca, A.; D’Amico, M.; De Amicis, F.; Pezzi, V. The Involvement of Natural Polyphenols in Molecular Mechanisms Inducing Apoptosis in Tumor Cells: A Promising Adjuvant in Cancer Therapy. Int. J. Mol. Sci. 2023, 24, 1680. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The role of polyphenols in overcoming cancer drug resistance: A comprehensive review. Cell Mol. Biol. Lett. 2022, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Rendeiro, C.; Rhodes, J.S.; Spencer, J.P. The mechanisms of action of flavonoids in the brain: Direct versus indirect effects. Neurochem. Int. 2015, 89, 126–139. [Google Scholar] [CrossRef]
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Caffo, M.; Minutoli, L.; Marini, H.; Abbritti, R.V.; Squadrito, F.; Trichilo, V.; Valenti, A.; Barresi, V.; Altavilla, D.; et al. ROS and Brain Gliomas: An Overview of Potential and Innovative Therapeutic Strategies. Int. J. Mol. Sci. 2016, 17, 984. [Google Scholar] [CrossRef]
- Mittal, P.; Dhankhar, S.; Chauhan, S.; Garg, N.; Bhattacharya, T.; Ali, M.; Chaudhary, A.A.; Rudayni, H.A.; Al-Zharani, M.; Ahmad, W.; et al. A Review on Natural Antioxidants for Their Role in the Treatment of Parkinson’s Disease. Pharmaceuticals 2023, 16, 908. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and cancer chemoprevention. Evid. Based Complement Alternat. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef]
- Bastianetto, S.; Ménard, C.; Quirion, R. Neuroprotective action of resveratrol. Biochim. Biophys. Acta 2015, 1852, 1195–1201. [Google Scholar] [CrossRef]
- Asgharian, P.; Tazekand, A.P.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Ranjbar, M.; Hasan, M.; Kumar, M.; Beirami, S.M.; Tarhriz, V.; et al. Potential mechanisms of quercetin in cancer prevention: Focus on cellular and molecular targets. Cancer Cell Int. 2022, 22, 257. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zheng, Y.; Guo, M.; Ares, I.; Martínez, M.; Lopez-Torres, B.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; Martínez, M.A. Oxidative stress, the blood-brain barrier and neurodegenerative diseases: The critical beneficial role of dietary antioxidants. Acta Pharm. Sin. B 2023, 13, 3988–4024. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Colonic metabolites of berry polyphenols: The missing link to biological activity? Br. J. Nutr. 2010, 104 (Suppl. S3), S48–S66. [Google Scholar] [CrossRef]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols journey through blood-brain barrier towards neuronal protection. Sci. Rep. 2017, 7, 11456. [Google Scholar] [CrossRef]
- Youdim, K.A.; Dobbie, M.S.; Kuhnle, G.; Proteggente, A.R.; Abbott, N.J.; Rice-Evans, C. Interaction between flavonoids and the blood-brain barrier: In vitro studies. J. Neurochem. 2003, 85, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid permeability across an in situ model of the blood-brain barrier. Free Radic. Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Trombino, S.; Curcio, F.; Sole, R.; Cassano, R.; Calviello, G. Hyaluronic Acid-Mediated Phenolic Compound Nanodelivery for Cancer Therapy. Pharmaceutics 2023, 15, 1751. [Google Scholar] [CrossRef] [PubMed]
- Jantas, D.; Malarz, J.; Le, T.N.; Stojakowska, A. Neuroprotective Properties of Kempferol Derivatives from Maesa membranacea against Oxidative Stress-Induced Cell Damage: An Association with Cathepsin D Inhibition and PI3K/Akt Activation. Int. J. Mol. Sci. 2021, 22, 10363. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aal, S.A.; AbdElrahman, M.; Reda, A.M.; Afify, H.; Ragab, G.M.; El-Gazar, A.A.; Ibrahim, S.S.A. Galangin mitigates DOX-induced cognitive impairment in rats: Implication of NOX-1/Nrf-2/HMGB1/TLR4 and TNF-α/MAPKs/RIPK/MLKL/BDNF. Neurotoxicology 2022, 92, 77–90. [Google Scholar] [CrossRef]
- Wei, T.; Wang, L.; Tang, J.; Ashaolu, T.J.; Olatunji, O.J. Protective effect of Juglanin against doxorubicin-induced cognitive impairment in rats: Effect on oxidative, inflammatory and apoptotic machineries. Metab. Brain Dis. 2022, 37, 1185–1195. [Google Scholar] [CrossRef]
- Kwatra, M.; Jangra, A.; Mishra, M.; Sharma, Y.; Ahmed, S.; Ghosh, P.; Kumar, V.; Vohora, D.; Khanam, R. Naringin and Sertraline Ameliorate Doxorubicin-Induced Behavioral Deficits Through Modulation of Serotonin Level and Mitochondrial Complexes Protection Pathway in Rat Hippocampus. Neurochem. Res. 2016, 41, 2352–2366. [Google Scholar] [CrossRef]
- Cheruku, S.P.; Ramalingayya, G.V.; Chamallamudi, M.R.; Biswas, S.; Nandakumar, K.; Nampoothiri, M.; Gourishetti, K.; Kumar, N. Catechin ameliorates doxorubicin-induced neuronal cytotoxicity in in vitro and episodic memory deficit in in vivo in Wistar rats. Cytotechnology 2018, 70, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Menze, E.T.; Tadros, M.G.; Tolba, M.F. Caffeic acid phenethyl ester counteracts doxorubicin-induced chemobrain in Sprague-Dawley rats: Emphasis on the modulation of oxidative stress and neuroinflammation. Neuropharmacology 2020, 181, 108334. [Google Scholar] [CrossRef]
- Moretti, R.L.; Dias, E.N.; Kiel, S.G.; Augusto, M.C.M.; Rodrigues, P.S.; Sampaio, A.C.S.; Medeiros, L.S.; Martins, M.F.M.; Suffredini, I.B.; Cardoso, C.V.; et al. Behavioral and morphological effects of resveratrol and curcumin in rats submitted to doxorubicin-induced cognitive impairment. Res. Vet. Sci. 2021, 140, 242–250. [Google Scholar] [CrossRef]
- Liao, D.; Shangguan, D.; Wu, Y.; Chen, Y.; Liu, N.; Tang, J.; Yao, D.; Shi, Y. Curcumin protects against doxorubicin induced oxidative stress by regulating the Keap1-Nrf2-ARE and autophagy signaling pathways. Psychopharmacology 2023, 240, 1179–1190. [Google Scholar] [CrossRef]
- Khadrawy, Y.A.; Hosny, E.N.; Mohammed, H.S. Protective effect of nanocurcumin against neurotoxicity induced by doxorubicin in rat’s brain. Neurotoxicology 2021, 85, 1–9. [Google Scholar] [CrossRef]
- Shi, D.D.; Dong, C.M.; Ho, L.C.; Lam, C.T.W.; Zhou, X.D.; Wu, E.X.; Zhou, Z.J.; Wang, X.M.; Zhang, Z.J. Resveratrol, a natural polyphenol, prevents chemotherapy-induced cognitive impairment: Involvement of cytokine modulation and neuroprotection. Neurobiol. Dis. 2018, 114, 164–173. [Google Scholar] [CrossRef]
- Tong, Y.; Wang, K.; Sheng, S.; Cui, J. Polydatin ameliorates chemotherapy-induced cognitive impairment (chemobrain) by inhibiting oxidative stress, inflammatory response, and apoptosis in rats. Biosci. Biotechnol. Biochem. 2020, 84, 1201–1210. [Google Scholar] [CrossRef]
- Harborne, J.B. Nature, distribution and function of plant flavonoids. Prog. Clin. Biol. Res. 1986, 213, 15–24. [Google Scholar]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Budzynska, B.; Faggio, C.; Kruk-Slomka, M.; Samec, D.; Nabavi, S.F.; Sureda, A.; Devi, K.P.; Nabavi, S.M. Rutin as Neuroprotective Agent: From Bench to Bedside. Curr. Med. Chem. 2019, 26, 5152–5164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J.; Sim, W.; Kim, J.H.; Choi, C.; Jeon, J. Discovering the anti-cancer phytochemical rutin against breast cancer through the methodical platform based on traditional medicinal knowledge. BMB Rep. 2023, 56, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Saleemi, M.A.; Alallam, B.; Yong, Y.K.; Lim, V. Synthesis of Zinc Oxide Nanoparticles with Bioflavonoid Rutin: Characterisation, Antioxidant and Antimicrobial Activities and In Vivo Cytotoxic Effects on Artemia Nauplii. Antioxidants 2022, 11, 1853. [Google Scholar] [CrossRef] [PubMed]
- Almatroudi, A.; Allemailem, K.S.; Alwanian, W.M.; Alharbi, B.F.; Alrumaihi, F.; Khan, A.A.; Almatroodi, S.A.; Rahmani, A.H. Effects and Mechanisms of Kaempferol in the Management of Cancers through Modulation of Inflammation and Signal Transduction Pathways. Int. J. Mol. Sci. 2023, 24, 8630. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef]
- Periferakis, A.; Periferakis, A.T.; Troumpata, L.; Periferakis, K.; Scheau, A.E.; Savulescu-Fiedler, I.; Caruntu, A.; Badarau, I.A.; Caruntu, C.; Scheau, C. Kaempferol: A Review of Current Evidence of Its Antiviral Potential. Int. J. Mol. Sci. 2023, 24, 16299. [Google Scholar] [CrossRef]
- Silva Dos Santos, J.; Gonçalves Cirino, J.P.; de Oliveira Carvalho, P.; Ortega, M.M. The Pharmacological Action of Kaempferol in Central Nervous System Diseases: A Review. Front. Pharmacol. 2021, 11, 565700. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar]
- Yang, J.; Lee, H.; Sung, J.; Kim, Y.; Jeong, H.S.; Lee, J. Conversion of Rutin to Quercetin by Acid Treatment in Relation to Biological Activities. Prev. Nutr. Food Sci. 2019, 24, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xiong, Y.Q.; Xu, J.; Wang, J.P.; Meng, Z.L.; Hong, Y.Q. Juglanin inhibits lung cancer by regulation of apoptosis, ROS and autophagy induction. Oncotarget 2017, 8, 93878–93898. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.R.; Zeng, K.; Lan, H.M.; Wang, Q. Juglanin ameliorates UVB-induced skin carcinogenesis via anti-inflammatory and proapoptotic effects in vivo and in vitro. Int. J. Mol. Med. 2018, 42, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.H.; Xu, S.P. Juglanin administration protects skin against UVB-induced injury by reducing Nrf2-dependent ROS generation. Int. J. Mol. Med. 2020, 46, 67–82. [Google Scholar] [CrossRef]
- Zhang, F.X.; Xu, R.S. Juglanin ameliorates LPS-induced neuroinflammation in animal models of Parkinson’s disease and cell culture via inactivating TLR4/NF-κB pathway. Biomed. Pharmacother. 2018, 97, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Pu, L.; Yu, L.; Xiong, F.; Sun, L.; Yu, Q.; Cao, X.; Chen, Y.; Peng, F.; et al. Galangin: A food-derived flavonoid with therapeutic potential against a wide spectrum of diseases. Phytother. Res. 2023, 37, 5700–5723. [Google Scholar] [CrossRef] [PubMed]
- Pakdeechote, P.; Poasakate, A.; Prasatthong, P.; Potue, P.; Khamseekaew, J.; Maneesai, P. Mitigation effect of galangin against aortic dysfunction and hypertrophy in rats with metabolic syndrome. Heliyon 2023, 9, e16500. [Google Scholar] [CrossRef]
- Huh, J.E.; Jung, I.T.; Choi, J.; Baek, Y.H.; Lee, J.D.; Park, D.S.; Choi, D.Y. The natural flavonoid galangin inhibits osteoclastic bone destruction and osteoclastogenesis by suppressing NF-κB in collagen-induced arthritis and bone marrow-derived macrophages. Eur. J. Pharmacol. 2013, 698, 57–66. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sak, K.; Adhikary, S.; Kaur, G.; Aggarwal, D.; Kaur, J.; Kumar, M.; Parashar, N.C.; Parashar, G.; Sharma, U.; et al. Galangin: A metabolite that suppresses anti-neoplastic activities through modulation of oncogenic targets. Exp. Biol. Med. 2022, 247, 345–359. [Google Scholar] [CrossRef]
- Zhang, F.; Yan, Y.; Zhang, L.M.; Li, D.X.; Li, L.; Lian, W.W.; Xia, C.Y.; He, J.; Xu, J.K.; Zhang, W.K. Pharmacological activities and therapeutic potential of galangin, a promising natural flavone, in age-related diseases. Phytomedicine 2023, 120, 155061. [Google Scholar] [CrossRef]
- Sangaraju, R.; Alavala, S.; Nalban, N.; Jerald, M.K.; Sistla, R. Galangin ameliorates Imiquimod-Induced psoriasis-like skin inflammation in BALB/c mice via down regulating NF-κB and activation of Nrf2 signaling pathways. Int. Immunopharmacol. 2021, 96, 107754. [Google Scholar] [CrossRef] [PubMed]
- Salama, S.A.; Elshafey, M.M. Galangin mitigates iron overload-triggered liver injury: Up-regulation of PPARγ and Nrf2 signaling, and abrogation of the inflammatory responses. Life Sci. 2021, 283, 119856. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Shen, L.; Xue, J.; Zou, J.; Wan, D.; Shi, Z. Therapeutic evaluation of galangin on cartilage protection and analgesic activity in a rat model of osteoarthritis. Electron. J. Biotechnol. 2021, 53, 8–13. [Google Scholar] [CrossRef]
- Gerges, S.H.; Tolba, M.F.; Elsherbiny, D.A.; El-Demerdash, E. The natural flavonoid galangin ameliorates dextran sulphate sodium-induced ulcerative colitis in mice: Effect on Toll-like receptor 4, inflammation and oxidative stress. Basic Clin. Pharmacol. Toxicol. 2020, 127, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, E.; Acar, G.; Onal, U.; Erdogan, E.; Baltaci, A.K.; Mogulkoc, R. Effect of 2-Week Naringin Supplementation on Neurogenesis and BDNF Levels in Ischemia-Reperfusion Model of Rats. Neuromolecular Med. 2024, 26, 4. [Google Scholar] [CrossRef] [PubMed]
- Isemura, M. Catechin in Human Health and Disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef] [PubMed]
- Alshatwi, A.A. Catechin hydrate suppresses MCF-7 proliferation through TP53/Caspase-mediated apoptosis. J. Exp. Clin. Cancer Res. 2010, 29, 167. [Google Scholar] [CrossRef] [PubMed]
- Bragança de Moraes, C.M.; Bitencourt, S.; de Mesquita, F.C.; Mello, D.; de Oliveira, L.P.; da Silva, G.V.; Lorini, V.; Caberlon, E.; de Souza Basso, B.; Schmid, J.; et al. (+)-Catechin attenuates activation of hepatic stellate cells. Cell Biol. Int. 2014, 38, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Chen, L.; Xiao, J.; Wang, C.; Jiang, W.; Zhang, R.; Hao, J. Chrysin protects against focal cerebral ischemia/reperfusion injury in mice through attenuation of oxidative stress and inflammation. Int. J. Mol. Sci. 2014, 15, 20913–20926. [Google Scholar] [CrossRef]
- Lee, B.K.; Lee, W.J.; Jung, Y.S. Chrysin attenuates VCAM-1 expression and monocyte adhesion in lipopolysaccharide-stimulated brain endothelial cells by preventing NF-κB signaling. Int. J. Mol. Sci. 2017, 18, 1424. [Google Scholar] [CrossRef]
- Krishnamoorthy, A.; Upadhyay, R.; Sevanan, M. Chrysin for Neurotrophic and Neurotransmitter Balance in Parkinson’s Disease. Methods Mol. Biol. 2024, 2761, 477–490. [Google Scholar] [PubMed]
- Aboud, H.M.; Ali, A.A.; El-Menshawe, S.F.; Elbary, A.A. Nanotransfersomes of carvedilol for intranasal delivery: Formulation, characterization and in vivo evaluation. Drug Deliv. 2016, 23, 2471–2481. [Google Scholar] [CrossRef] [PubMed]
- Sava, V.; Fihurka, O.; Khvorova, A.; Sanchez-Ramos, J. Enriched chitosan nanoparticles loaded with siRNA are effective in lowering Huntington’s disease gene expression following intranasal administration. Nanomedicine 2020, 24, 102119. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.R.; Pinto, C.S.C.; Santos, E.P.; Mansur, C.R.E. Hybrid Vesicular Nanosystems Based on Lipids and Polymers Applied in Therapy, Theranostics, and Cosmetics. Crit. Rev. Ther. Drug Carrier Syst. 2020, 37, 271–303. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Babu, S.S.; Naroth, S.; Janardanan, D. Antioxidant activity of caffeic acid: Thermodynamic and kinetic aspects on the oxidative degradation pathway. Free Radic. Res. 2022, 56, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Olgierd, B.; Kamila, Ż.; Anna, B.; Emilia, M. The Pluripotent Activities of Caffeic Acid Phenethyl Ester. Molecules 2021, 26, 1335. [Google Scholar] [CrossRef]
- Melo, J.B.; Agostinho, P.; Oliveira, C.R. Involvement of oxidative stress in the enhancement of acetylcholinesterase activity induced by amyloid beta-peptide. Neurosci. Res. 2003, 45, 117–127. [Google Scholar] [CrossRef]
- Philpot, R.M.; Ficken, M.; Johns, B.E.; Engberg, M.E.; Wecker, L. Spatial memory deficits in mice induced by chemotherapeutic agents are prevented by acetylcholinesterase inhibitors. Cancer Chemother. Pharmacol. 2019, 84, 579–589. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef]
- Kocaadam, B.; Şanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Hewlings, S.; Kalman, D. Curcumin: A Review of Its’ Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Khalili, N.; Sahebi, E.; Namazi, S.; Reiner, Ž.; Majeed, M.; Sahebkar, A. Curcuminoids modify lipid profile in type 2 diabetes mellitus: A randomized controlled trial. Complement Ther. Med. 2017, 33, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, S.; Asgary, S.; Askari, G.; Keshvari, M.; Hatamipour, M.; Feizi, A.; Sahebkar, A. Treatment of Non-alcoholic Fatty Liver Disease with Curcumin: A Randomized Placebo-controlled Trial. Phytother. Res. 2016, 30, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Khoi, M.J.; Azizi, M.; Goodarzi, M. Can curcumin and its analogs be a new treatment option in cancer therapy? Cancer Gene Ther. 2016, 23, 410. [Google Scholar] [CrossRef] [PubMed]
- Momtazi-Borojeni, A.A.; Haftcheshmeh, S.M.; Esmaeili, S.A.; Johnston, T.P.; Abdollahi, E.; Sahebkar, A. Curcumin: A natural modulator of immune cells in systemic lupus erythematosus. Autoimmun. Rev. 2018, 17, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Esmaily, H.; Sahebkar, A.; Iranshahi, M.; Ganjali, S.; Mohammadi, A.; Ferns, G.; Ghayour-Mobarhan, M. An investigation of the effects of curcumin on anxiety and depression in obese individuals: A randomized controlled trial. Chin. J. Integr. Med. 2015, 21, 332–338. [Google Scholar] [CrossRef]
- Hassanizadeh, S.; Shojaei, M.; Bagherniya, M.; Orekhov, A.N.; Sahebkar, A. Effect of nano-curcumin on various diseases: A comprehensive review of clinical trials. Biofactors 2023, 49, 512–533. [Google Scholar] [CrossRef]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.; Crozier, A. Plant foods and herbal sources of resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Sales, J.M.; Resurreccion, A.V. Resveratrol in peanuts. Crit. Rev. Food Sci. Nutr. 2014, 54, 734–770. [Google Scholar] [CrossRef]
- Shrikanta, A.; Kumar, A.; Govindaswamy, V. Resveratrol content and antioxidant properties of underutilized fruits. J. Food Sci. Technol. 2015, 52, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Kwah, M.X.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.; Wang, L.; Ong, P.S.; et al. Resveratrol for cancer therapy: Challenges and future perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Breuss, J.M.; Atanasov, A.G.; Uhrin, P. Resveratrol and Its Effects on the Vascular System. Int. J. Mol. Sci. 2019, 20, 1523. [Google Scholar] [CrossRef]
- Abhale, K.; Veeranjaneyulu, A.; Desai, S.; Sanap, A.; Bhonde, R. Effects of Mesenchymal Stem Cell-conditioned Media with Natural Immunomodulatory Agent Resveratrol on Type 1 Diabetes. Curr. Drug Discov. Technol. 2024. online ahead of print. [Google Scholar] [CrossRef]
- Chen, Z.; Xiao, G.; Ao, J. Resveratrol Attenuates Rheumatoid Arthritis Induce Neutrophil Extracellular Traps via TLR-4 Mediated Inflammation in C57BL/6 Mice. Physiol. Res. 2024, 73, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.; McElroy, T.; Simmons, P.; Walters, H.; Ntagwabira, F.; Wang, J.; Byrum, S.D.; Allen, A.R. Cognitive impairment resulting from treatment with docetaxel, doxorubicin, and cyclophosphamide. Brain Res. 2021, 1760, 147397. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Nan, A. Combination drug delivery approaches in metastatic breast cancer. J. Drug Deliv. 2012, 2012, 915375. [Google Scholar] [CrossRef]
- Orabi, M.A.A.; Khalil, H.M.A.; Abouelela, M.E.; Zaafar, D.; Ahmed, Y.H.; Naggar, R.A.; Alyami, H.S.; Abdel-Sattar, E.S.; Matsunami, K.; Hamdan, D.I. Carissa macrocarpa Leaves Polar Fraction Ameliorates Doxorubicin-Induced Neurotoxicity in Rats via Downregulating the Oxidative Stress and Inflammatory Markers. Pharmaceuticals 2021, 14, 1305. [Google Scholar] [CrossRef]
- El-Din, M.I.G.; George, M.Y.; Youssef, F.S. Chemical characterization of the polyphenolic rich fraction of Thunbergia erecta and its therapeutic potential against doxorubicin and cyclophosphamide-induced cognitive impairment in rats. J. Ethnopharmacol. 2023, 307, 116213. [Google Scholar] [CrossRef]
- Serini, S.; Cassano, R.; Bruni, M.; Servidio, C.; Calviello, G.; Trombino, S. Characterization of a hyaluronic acid and folic acid-based hydrogel for cisplatin delivery: Antineoplastic effect in human ovarian cancer cells in vitro. Int. J. Pharm. 2021, 606, 120899. [Google Scholar] [CrossRef]
- Rezaei, T.; Rezaei, M.; Karimifard, S.; Mahmoudi Beram, F.; Dakkali, M.S.; Heydari, M.; Afshari-Behbahanizadeh, S.; Mostafavi, E.; Bokov, D.O.; Ansari, M.J.; et al. Folic Acid-Decorated pH-Responsive Nanoniosomes with Enhanced Endocytosis for Breast Cancer Therapy: In Vitro Studies. Front. Pharmacol. 2022, 13, 851242. [Google Scholar] [CrossRef] [PubMed]
- Sahrayi, H.; Hosseini, E.; Karimifard, S.; Khayam, N.; Meybodi, S.M.; Amiri, S.; Bourbour, M.; Farasati Far, B.; Akbarzadeh, I.; Bhia, M.; et al. Co-Delivery of Letrozole and Cyclophosphamide via Folic Acid-Decorated Nanoniosomes for Breast Cancer Therapy: Synergic Effect, Augmentation of Cytotoxicity, and Apoptosis Gene Expression. Pharmaceuticals 2021, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, J.; Taylor, J.; Mehta, M.; Satija, S.; Paudel, K.R.; Hansbro, P.M.; Chellappan, D.K.; Bebawy, M.; Dua, K. Targeting Cancer using Curcumin Encapsulated Vesicular Drug Delivery Systems. Curr. Pharm. Des. 2021, 27, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Cassano, R.; Facchinetti, E.; Amendola, G.; Trombino, S.; Calviello, G. Anti-Irritant and Anti-Inflammatory Effects of DHA Encapsulated in Resveratrol-Based Solid Lipid Nanoparticles in Human Keratinocytes. Nutrients 2019, 11, 1400. [Google Scholar] [CrossRef] [PubMed]
- Trombino, S.; Serini, S.; Cassano, R.; Calviello, G. Xanthan gum-based materials for omega-3 PUFA delivery: Preparation, characterization and antineoplastic activity evaluation. Carbohydr. Polym. 2019, 208, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Cassano, R.; Corsetto, P.A.; Rizzo, A.M.; Calviello, G.; Trombino, S. Omega-3 PUFA Loaded in Resveratrol-Based Solid Lipid Nanoparticles: Physicochemical Properties and Antineoplastic Activities in Human Colorectal Cancer Cells In Vitro. Int. J. Mol. Sci. 2018, 19, 586. [Google Scholar] [CrossRef]
- Cassano, R.; Serini, S.; Curcio, F.; Trombino, S.; Calviello, G. Preparation and Study of Solid Lipid Nanoparticles Based on Curcumin, Resveratrol and Capsaicin Containing Linolenic Acid. Pharmaceutics 2022, 14, 1593. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, S.; Sun, M.; Shi, J.; Zhang, H.; Yu, H.; Yang, Z. Recent advances and clinical translation of liposomal delivery systems in cancer therapy. Eur. J. Pharm. Sci. 2024, 193, 106688. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, L.; Zhang, B.; Xu, Z.P. MnO2-shelled Doxorubicin/Curcumin nanoformulation for enhanced colorectal cancer chemo-immunotherapy. J. Colloid Interface Sci. 2022, 617, 315–325. [Google Scholar] [CrossRef]
- Mirzaei, S.; Gholami, M.H.; Zabolian, A.; Saleki, H.; Bagherian, M.; Torabi, S.M.; Sharifzadeh, S.O.; Hushmandi, K.; Fives, K.R.; Khan, H.; et al. Resveratrol Augments Doxorubicin and Cisplatin Chemotherapy: A Novel Therapeutic Strategy. Curr. Mol. Pharmacol. 2023, 16, 280–306. [Google Scholar]
- Saharkhiz, S.; Zarepour, A.; Zarrabi, A. A new theranostic pH-responsive niosome formulation for doxorubicin delivery and bio-imaging against breast cancer. Int. J. Pharm. 2023, 637, 122845. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serini, S.; Calviello, G. Potential of Natural Phenolic Compounds against Doxorubicin-Induced Chemobrain: Biological and Molecular Mechanisms Involved. Antioxidants 2024, 13, 486. https://doi.org/10.3390/antiox13040486
Serini S, Calviello G. Potential of Natural Phenolic Compounds against Doxorubicin-Induced Chemobrain: Biological and Molecular Mechanisms Involved. Antioxidants. 2024; 13(4):486. https://doi.org/10.3390/antiox13040486
Chicago/Turabian StyleSerini, Simona, and Gabriella Calviello. 2024. "Potential of Natural Phenolic Compounds against Doxorubicin-Induced Chemobrain: Biological and Molecular Mechanisms Involved" Antioxidants 13, no. 4: 486. https://doi.org/10.3390/antiox13040486
APA StyleSerini, S., & Calviello, G. (2024). Potential of Natural Phenolic Compounds against Doxorubicin-Induced Chemobrain: Biological and Molecular Mechanisms Involved. Antioxidants, 13(4), 486. https://doi.org/10.3390/antiox13040486