Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
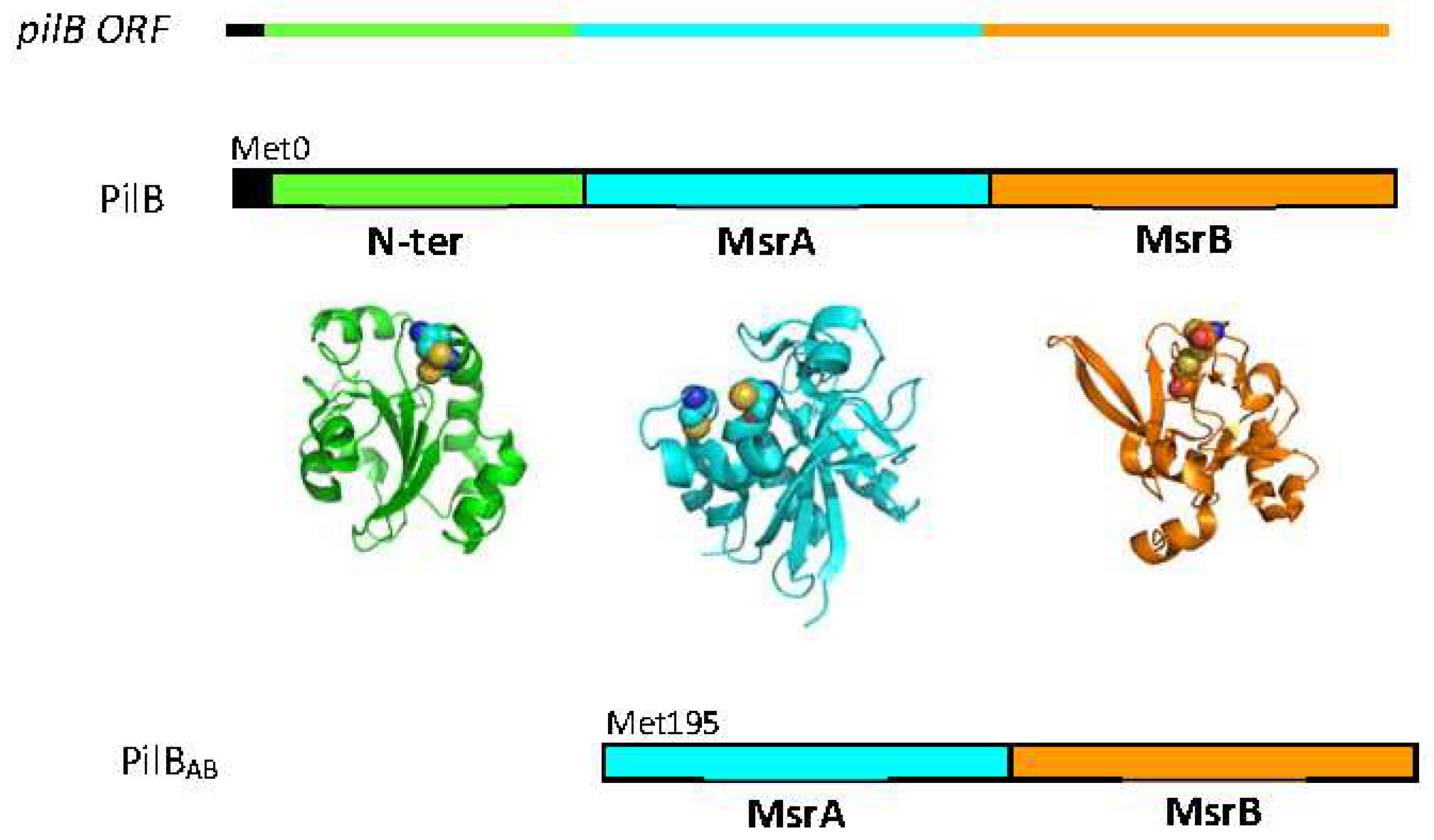
2. Methionine Sulfoxide Reductase Activities of PilB
2.1. MSRs of N. meningitidis
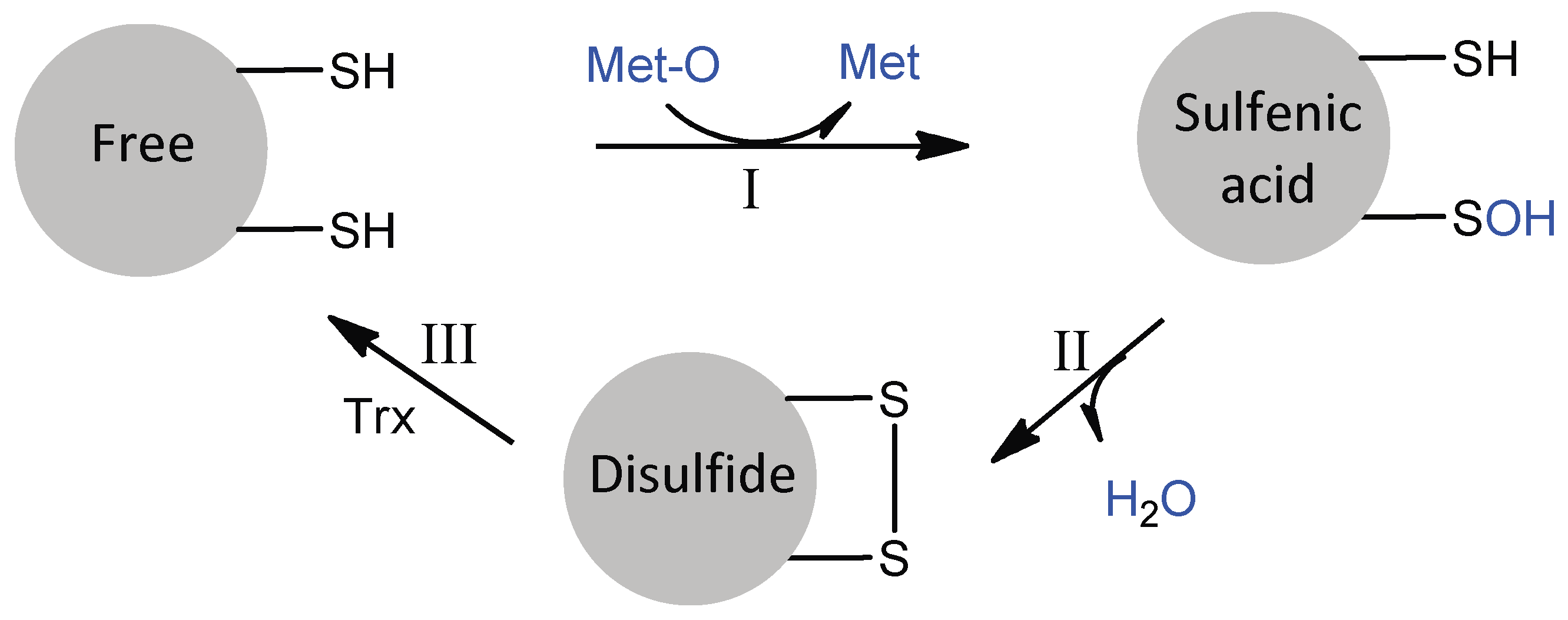
2.2. Catalytic Mechanism of MSR Domains
2.3. Substrate Specificities of MSR Domains
2.4. Catalytic Efficiency of MSRA/MSRB Fusion
3. Recycling of MSR Activities within PilB
3.1. The N-ter Domain
3.2. Recycling of MSRs Activities within PilB
4. Periplasmic Recycling Partners
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Taha, M.K.; Dupuy, B.; Saurin, W.; So, M.; Marchal, C. Control of pilus expression in Neisseria gonorrhoeae as an original system in the family of two-component regulators. Mol. Microbiol. 1991, 5, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.K.; So, M.; Seifert, H.S.; Billyard, E.; Marchal, C. Pilin expression in Neisseria gonorrhoeae is under both positive and negative transcriptional control. EMBO J. 1988, 7, 4367–4378. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.K.; Marchal, C. Conservation of Neisseria gonorrhoeae pilus expression regulatory genes pilA and pilB in the genus Neisseria. Infect. Immun. 1990, 58, 4145–4148. [Google Scholar] [PubMed]
- Skaar, E.P.; Tobiason, D.M.; Quick, J.; Judd, R.C.; Weissbach, H.; Etienne, F.; Brot, N.; Seifert, H.S. The outer membrane localization of the Neisseria gonorrhoeae MSRA/B is involved in survival against reactive oxygen species. Proc. Natl. Acad. Sci. USA 2002, 99, 10108–10113. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Neiers, F.; Boschi-Muller, S.; Branlant, G. The N-terminal domain of PilB from Neisseria meningitidis is a disulfide reductase that can recycle methionine sulfoxide reductases. J. Biol. Chem. 2005, 280, 12344–12350. [Google Scholar] [CrossRef] [PubMed]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Grimaud, R.; Ezraty, B.; Mitchell, J.K.; Lafitte, D.; Briand, C.; Derrick, P.J.; Barras, F. Repair of oxidized proteins. Identification of a new methionine sulfoxide reductase. J. Biol. Chem. 2001, 276, 48915–48920. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Johnson, L.C.; Weissbach, H.; Brot, N.; Lively, M.O.; Lowther, W.T. Free methionine-(R)-sulfoxide reductase from Escherichia coli reveals a new GAF domain function. Proc. Natl. Acad. Sci. USA 2007, 104, 9597–9602. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Poston, J.M.; Berlett, B.S.; Nosworthy, N.J.; Szczepanowski, R.; Stadtman, E.R. Identification and characterization of a putative active site for peptide methionine sulfoxide reductase (MSRA) and its substrate stereospecificity. J. Biol. Chem. 2000, 275, 14167–14172. [Google Scholar] [CrossRef] [PubMed]
- Gruez, A.; Libiad, M.; Boschi-Muller, S.; Branlant, G. Structural and biochemical characterization of free methionine-R-sulfoxide reductase from Neisseria meningitidis. J. Biol. Chem. 2010, 285, 25033–25043. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Gand, A.; Branlant, G. The methionine sulfoxide reductases: Catalysis and substrate specificities. Arch. Biochem. Biophys. 2008, 474, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Olry, A.; Antoine, M.; Branlant, G. The enzymology and biochemistry of methionine sulfoxide reductases. Biochim. Biophys. Acta 2005, 1703, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Branlant, G. Methionine sulfoxide reductase: Chemistry, substrate binding, recycling process and oxidase activity. Bioorg. Chem. 2014, 57, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Kauffmann, B.; Aubry, A.; Favier, F. The three-dimensional structures of peptide methionine sulfoxide reductases: Current knowledge and open questions. Biochim. Biophys. Acta 2005, 1703, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Azza, S.; Sanglier-Cianferani, S.; Talfournier, F.; Van Dorsselear, A.; Branlant, G. A sulfenic acid enzyme intermediate is involved in the catalytic mechanism of peptide methionine sulfoxide reductase from Escherichia coli. J. Biol. Chem. 2000, 275, 35908–35913. [Google Scholar] [CrossRef] [PubMed]
- Olry, A.; Boschi-Muller, S.; Marraud, M.; Sanglier-Cianferani, S.; Van Dorsselear, A.; Branlant, G. Characterization of the methionine sulfoxide reductase activities of PILB, a probable virulence factor from Neisseria meningitidis. J. Biol. Chem. 2002, 277, 12016–12022. [Google Scholar] [CrossRef] [PubMed]
- Lowther, W.T.; Weissbach, H.; Etienne, F.; Brot, N.; Matthews, B.W. The mirrored methionine sulfoxide reductases of Neisseria gonorrhoeae pilB. Nat. Struct. Biol. 2002, 9, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Ranaivoson, F.M.; Neiers, F.; Kauffmann, B.; Boschi-Muller, S.; Branlant, G.; Favier, F. Methionine sulfoxide reductase B displays a high level of flexibility. J. Mol. Biol. 2009, 394, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Ranaivoson, F.M.; Antoine, M.; Kauffmann, B.; Boschi-Muller, S.; Aubry, A.; Branlant, G.; Favier, F. A structural analysis of the catalytic mechanism of methionine sulfoxide reductase A from Neisseria meningitidis. J. Mol. Biol. 2008, 377, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Dokainish, H.M.; Gauld, J.W. A molecular dynamics and quantum mechanics/molecular mechanics study of the catalytic reductase mechanism of methionine sulfoxide reductase A: Formation and reduction of a sulfenic acid. Biochemistry 2013, 52, 1814–1827. [Google Scholar] [CrossRef] [PubMed]
- Robinet, J.J.; Dokainish, H.M.; Paterson, D.J.; Gauld, J.W. A sulfonium cation intermediate in the mechanism of methionine sulfoxide reductase B: A DFT study. J. Phys. Chem. B 2011, 115, 9202–9212. [Google Scholar] [CrossRef] [PubMed]
- Thiriot, E.; Monard, G.; Boschi-Muller, S.; Branlant, G.; Ruiz-López, M.F. Reduction mechanism in class A methionine sulfoxide reductases: A theoretical chemistry investigation. Theor. Chem. Acc. 2011, 129, 93–103. [Google Scholar] [CrossRef]
- Antoine, M.; Boschi-Muller, S.; Branlant, G. Kinetic characterization of the chemical steps involved in the catalytic mechanism of methionine sulfoxide reductase A from Neisseria meningitidis. J. Biol. Chem. 2003, 278, 45352–45357. [Google Scholar] [CrossRef] [PubMed]
- Olry, A.; Boschi-Muller, S.; Branlant, G. Kinetic characterization of the catalytic mechanism of methionine sulfoxide reductase B from Neisseria meningitidis. Biochemistry 2004, 43, 11616–11622. [Google Scholar] [CrossRef] [PubMed]
- Gruez, A.; Branlant, G. Structural diversity in the recognition between reduced thioredoxin and its oxidized enzyme partners. Biomol. Concepts 2012, 3, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Gand, A.; Antoine, M.; Boschi-Muller, S.; Branlant, G. Characterization of the amino acids involved in substrate specificity of methionine sulfoxide reductase A. J. Biol. Chem. 2007, 282, 20484–20491. [Google Scholar] [CrossRef] [PubMed]
- Neiers, F.; Sonkaria, S.; Olry, A.; Boschi-Muller, S.; Branlant, G. Characterization of the amino acids from Neisseria meningitidis methionine sulfoxide reductase B involved in the chemical catalysis and substrate specificity of the reductase step. J. Biol. Chem. 2007, 282, 32397–32405. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Markillie, L.M.; Xiong, Y.; Mayer, M.U.; Squier, T.C. Increased catalytic efficiency following gene fusion of bifunctional methionine sulfoxide reductase enzymes from Shewanella oneidensis. Biochemistry 2007, 46, 14153–14161. [Google Scholar] [CrossRef] [PubMed]
- Han, A.-R.; Kim, M.-J.; Kwak, G.-H.; Son, J.; Hwang, K.Y.; Kim, H.-Y. Essential Role of the Linker Region in the Higher Catalytic Efficiency of a Bifunctional MSRA-MSRB Fusion Protein. Biochemistry 2016, 55, 5117–5127. [Google Scholar] [CrossRef] [PubMed]
- Quinternet, M.; Tsan, P.; Selme, L.; Beaufils, C.; Jacob, C.; Boschi-Muller, S.; Averlant-Petit, M.-C.; Branlant, G.; Cung, M.-T. Solution structure and backbone dynamics of the cysteine 103 to serine mutant of the N-terminal domain of DsbD from Neisseria meningitidis. Biochemistry 2008, 47, 12710–12720. [Google Scholar] [CrossRef] [PubMed]
- Ranaivoson, F.M.; Kauffmann, B.; Neiers, F.; Wu, J.; Boschi-Muller, S.; Panjikar, S.; Aubry, A.; Branlant, G.; Favier, F. The X-ray structure of the N-terminal domain of PILB from Neisseria meningitidis reveals a thioredoxin-fold. J. Mol. Biol. 2006, 358, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Quinternet, M.; Tsan, P.; Selme-Roussel, L.; Jacob, C.; Boschi-Muller, S.; Branlant, G.; Cung, M.-T. Formation of the complex between DsbD and PilB N-terminal domains from Neisseria meningitidis necessitates an adaptability of nDsbD. Structure 2009, 17, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Stirnimann, C.U.; Rozhkova, A.; Grauschopf, U.; Grütter, M.G.; Glockshuber, R.; Capitani, G. Structural basis and kinetics of DsbD-dependent cytochrome c maturation. Structure 2005, 13, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Gand, A.; Selme-Roussel, L.; Collin, S.; Branlant, G.; Jacob, C.; Boschi-Muller, S. Biochemical and functional characterization of a periplasmic disulfide oxidoreductase from Neisseria meningitidis essential for meningococcal viability. Biochem. J. 2015, 468, 271–282. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boschi-Muller, S. Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis. Antioxidants 2018, 7, 131. https://doi.org/10.3390/antiox7100131
Boschi-Muller S. Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis. Antioxidants. 2018; 7(10):131. https://doi.org/10.3390/antiox7100131
Chicago/Turabian StyleBoschi-Muller, Sandrine. 2018. "Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis" Antioxidants 7, no. 10: 131. https://doi.org/10.3390/antiox7100131
APA StyleBoschi-Muller, S. (2018). Molecular Mechanisms of the Methionine Sulfoxide Reductase System from Neisseria meningitidis. Antioxidants, 7(10), 131. https://doi.org/10.3390/antiox7100131