CoQ10 Supplementation in Patients Undergoing IVF-ET: The Relationship with Follicular Fluid Content and Oocyte Maturity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Selection and Study Design
2.2. Follicular Stimulation
2.3. Follicular Fluids CoQ10 Levels
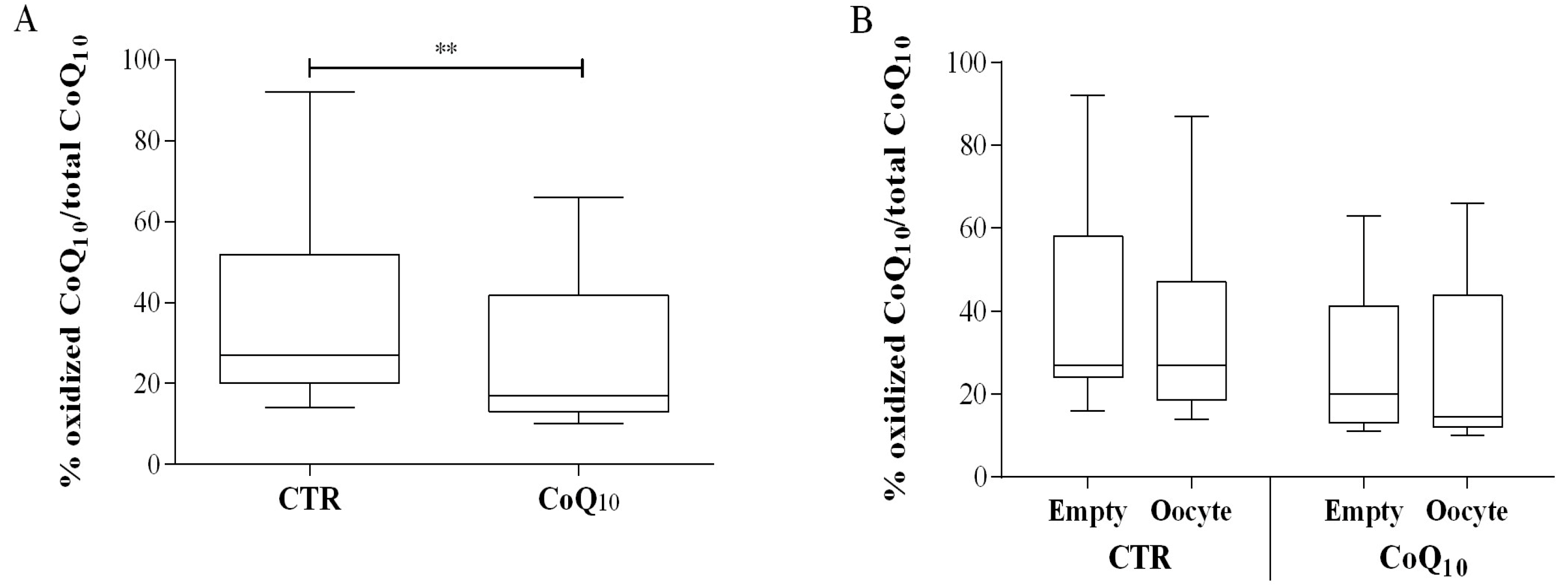
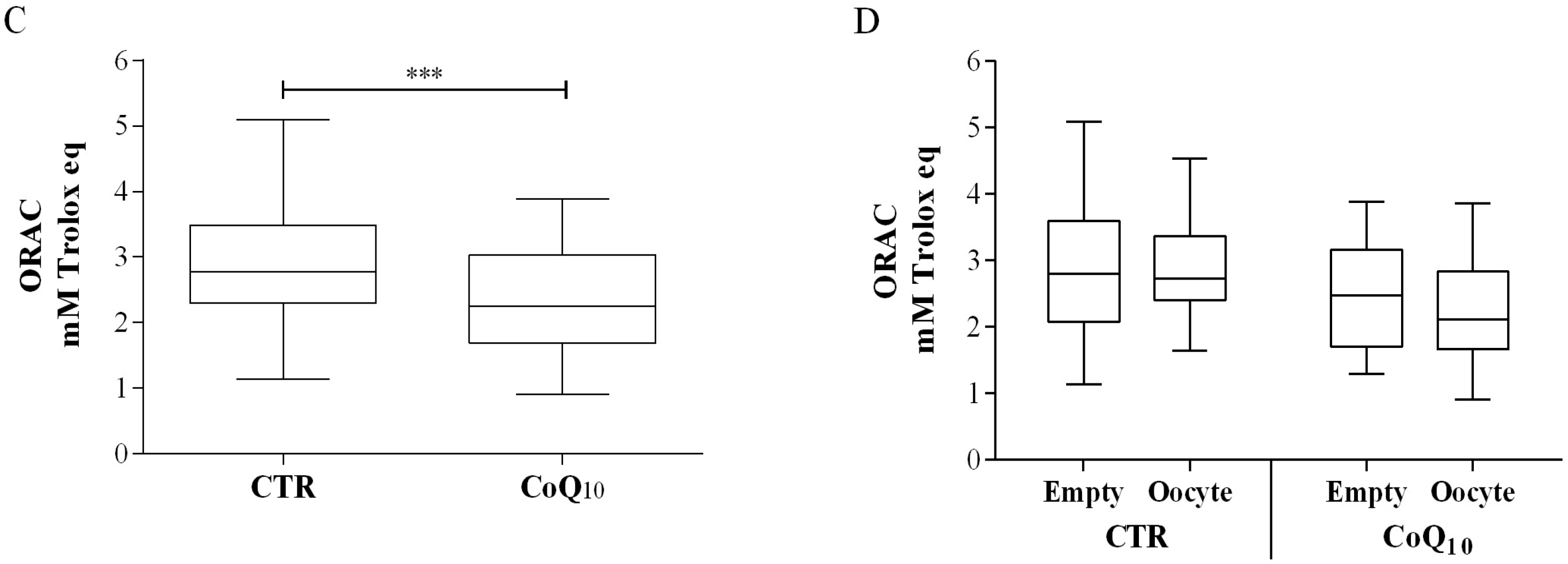
2.4. ORAC Assay of Follicular Fluids
2.5. Statistical Analysis
3. Results
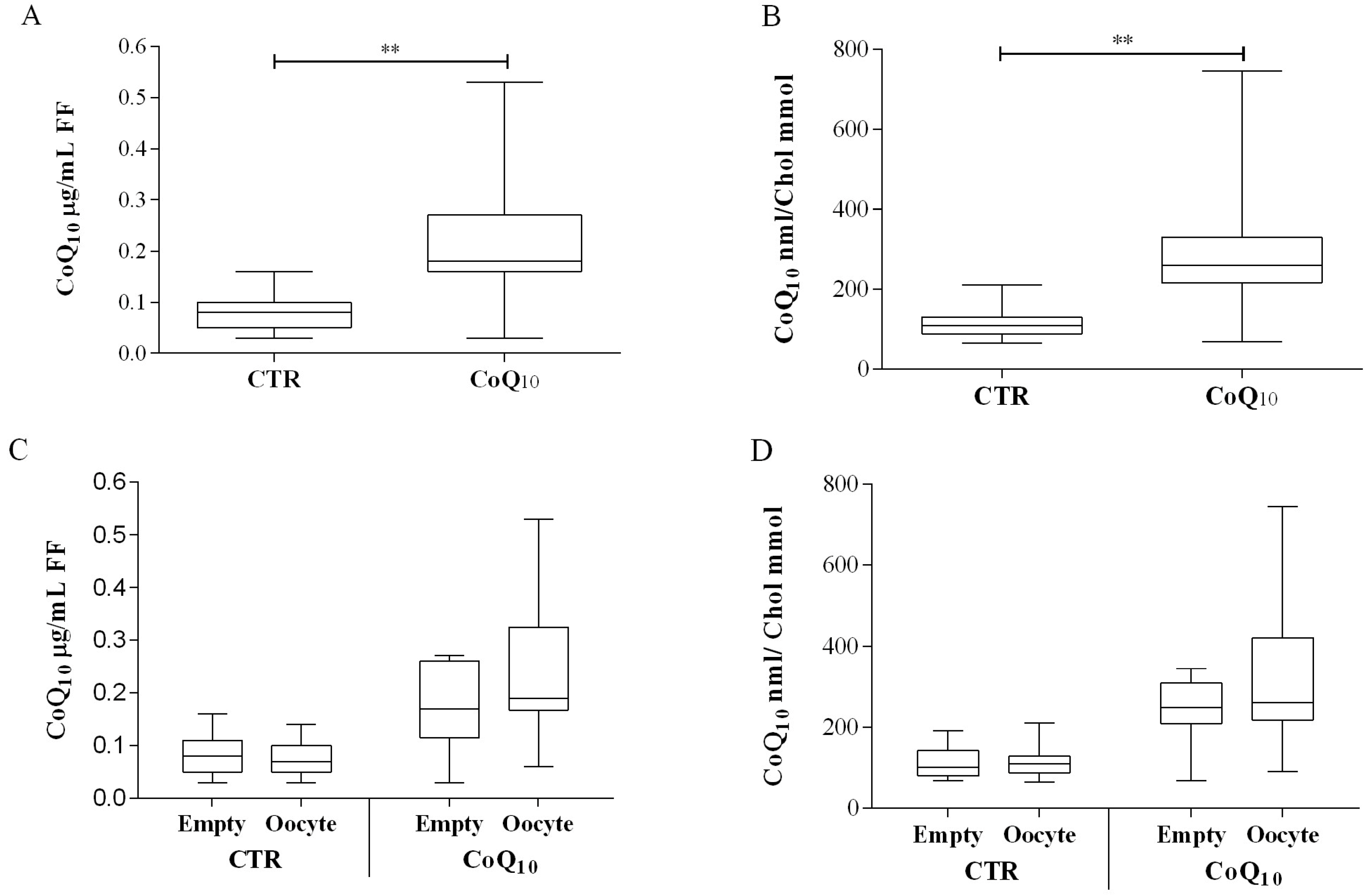
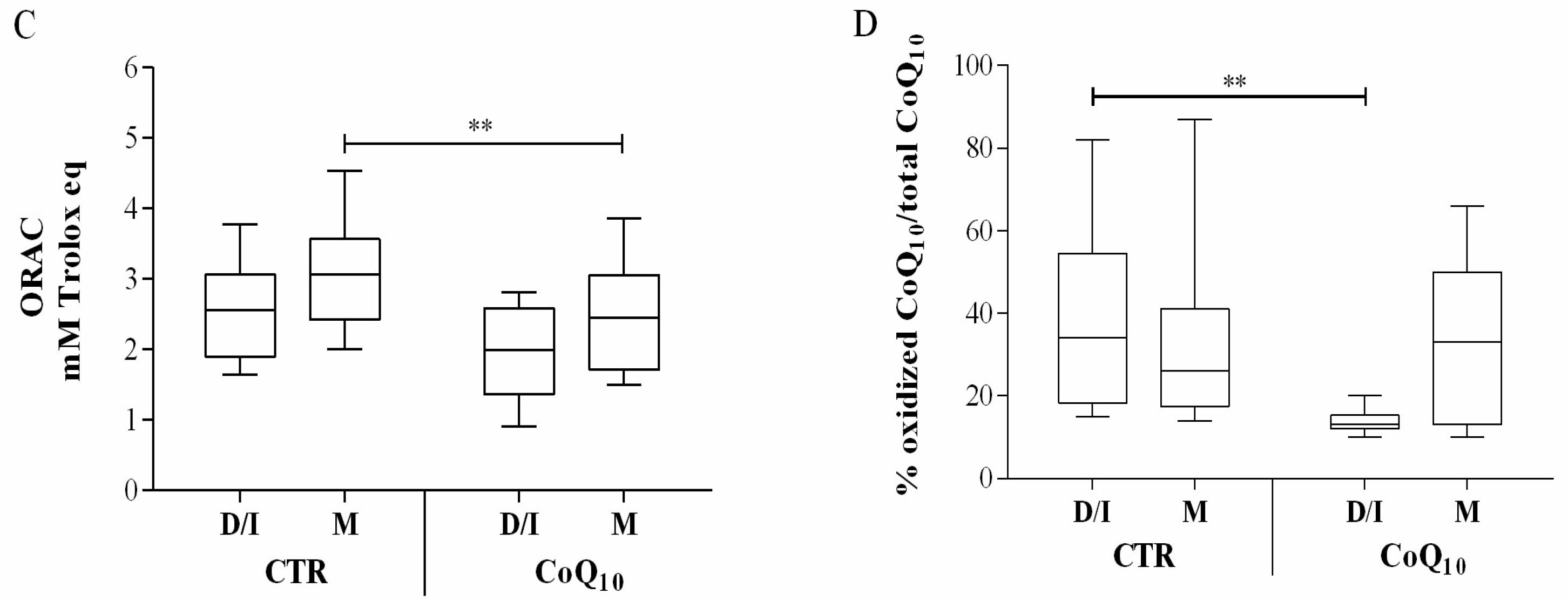
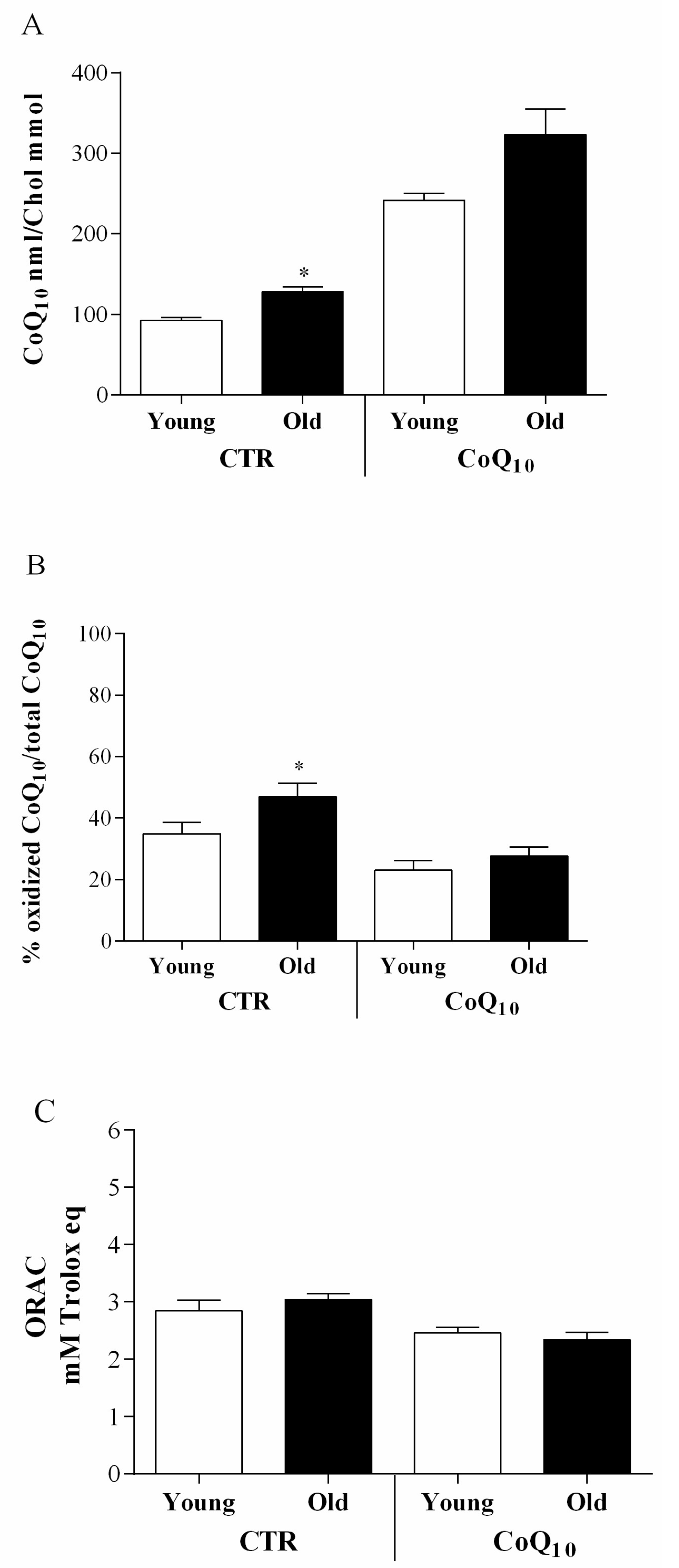
3.1. Follicular Coenzyme Q10 Content and Oxidative Status
3.2. Sub Analysis of Oocyte-Containing Follicles
3.3. Fertilization Rate and Embryo Grading
3.4. Age Related Considerations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carolan, M.; Frankowska, D. Advanced maternal age and adverse perinatal outcome: A review of the evidence. Midwifery 2011, 27, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Kuliev, A.; Cieslak, J.; Verlinsky, Y. Frequency and distribution of chromosome abnormalities in human oocytes. Cytogenet Genome Res. 2005, 111, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Pellestor, F.; Anahory, T.; Hamamah, S. Effect of maternal age on the frequency of cytogenetic abnormalities in human oocytes. Cytogenet Genome Res. 2005, 111, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Von Wald, T.; Monisova, Y.; Hacker, M.R.; Yoo, S.W.; Penzias, A.S.; Reindollar, R.R.; Usheva, A. Age-related variations in follicular apolipoproteins may influence human oocyte maturation and fertility potential. Fertil. Steril. 2010, 93, 2354–2361. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, V.Y.; Kane, J.P.; Ishida, B.Y.; Bloom, M.S.; Browne, R.W. High-density lipoprotein metabolism and the human embryo. Hum. Reprod. Update 2010, 16, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Bausenwein, J.; Serke, H.; Eberle, K.; Hirrlinger, J.; Jogschies, P.; Hmeidan, F.A.; Blumenauer, V.; Spanel-Borowski, K. Elevated levels of oxidized low-density lipoprotein and of catalase activity in follicular fluid of obese women. Mol. Hum. Reprod. 2010, 16, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Hanukoglu, I. Antioxidant protective mechanisms against reactive oxygen species (ROS) generated by mitochondrial P450 systems in steroidogenic cells. Drug. Metab. Rev. 2006, 38, 171–196. [Google Scholar] [CrossRef] [PubMed]
- Bentov, Y.; Esfandiari, N.; Burstein, E.; Casper, R.F. The use of mitochondrial nutrients to improve the outcome of infertility treatment in older patients. Fertil. Steril. 2010, 93, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.Y.; Gao, Y.; Jiao, G.Z.; Sun, M.J.; Wu, X.F.; Wang, T.Y.; Li, H.; Tan, J.H. Antioxidant supplementation overcomes the deleterious effects of maternal restraint stress-induced oxidative stress on mouse oocytes. Reproduction 2013, 146, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, S.; Babu, N.K.; Chattopadhyay, R.; Chakravarty, B.; Chaudhury, K. Upper control limit of reactive oxygen species in follicular fluid beyond which viable embryo formation is not favorable. Reprod. Toxicol. 2010, 29, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Littarru, G.P.; Tiano, L. Clinical aspects of coenzyme Q10: An update. Nutrition 2010, 26, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Littarru, G.P.; Tiano, L. Bioenergetic and antioxidant properties of coenzyme Q10: Recent developments. Mol. Biotechnol. 2007, 37, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Balercia, G.; Gandini, L.; Lenzi, A.; Lombardo, F. Antioxidants in Andrology, 1st ed.; Springer: Cham, Switzerland, 2017; pp. 23–34. ISBN 978-3-319-41747-9. [Google Scholar]
- Balercia, G.; Arnaldi, G.; Fazioli, F.; Serresi, M.; Alleva, R.; Mancini, A.; Mosca, F.; Lamonica, G.R.; Mantero, F.; Littarru, G.P. Coenzyme Q10 levels in idiopathic and varicocele-associated asthenozoospermia. Andrologia 2002, 34, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Balercia, G.; Buldreghini, E.; Vignini, A.; Tiano, L.; Paggi, F.; Amoroso, S.; Ricciardo-Lamonica, G.; Boscaro, M.; Lenzi, A.; Littarru, G. Coenzyme Q10 treatment in infertile men with idiopathic asthenozoospermia: A placebo-controlled, double-blind randomized trial. Fertil. Steril. 2009, 91, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Tirabassi, G.; Vignini, A.; Tiano, L.; Buldreghini, E.; Brugè, F.; Silvestri, S.; Orlando, P.; D’Aniello, A.; Mazzanti, L.; Lenzi, A.; et al. Protective effects of coenzyme Q10 and aspartic acid on oxidative stress and DNA damage in subjects affected by idiopathic asthenozoospermia. Endocrine 2015, 49, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Bentinger, M.; Tekle, M.; Brismar, K.; Chojnacki, T.; Swiezewska, E.; Dallner, G. Stimulation of coenzyme Q synthesis. Biofactors 2008, 32, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, M.; Westesen, K.; Zakhartchenko, V.; Stojkovic, P.; Boxhammer, K.; Wolf, E. Coenzyme Q(10) in submicron-sized dispersion improves development, hatching, cell proliferation, and adenosine triphosphate content of in vitro-produced bovine embryos. Biol. Reprod. 1999, 61, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Turi, A.; Giannubilo, S.R.; Brugè, F.; Principi, F.; Battistoni, S.; Santoni, F.; Tranquilli, A.L.; Littarru, G.; Tiano, L. Coenzyme Q10 content in follicular fluid and its relationship with oocyte fertilization and embryo grading. Arch. Gynecol. Obstet. 2012, 285, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Akarsu, S.; Gode, F.; Isik, A.Z.; Dikmen, Z.G.; Tekindal, M.A. The association between coenzyme Q10 concentrations in follicular fluid withembryo morphokinetics and pregnancy rate in assisted reproductive techniques. J. Assist. Reprod. Genet. 2017, 34, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Gat, I.; Blanco Mejia, S.; Balakier, H.; Librach, C.L.; Claessens, A.; Ryan, E.A. The use of coenzyme Q10 and DHEA during IUI and IVF cycles in patients with decreased ovarian reserve. Gynecol. Endocrinol. 2016, 32, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Nisenblat, V.; Lu, C.; Li, R.; Qiao, J.; Zhen, X.; Wang, S. Pretreatment with coenzyme Q10 improves ovarian response and embryo quality in low-prognosis young women with decreased ovarian reserve: A randomized controlled trial. Reprod. Biol. Endocrinol. 2018, 16, 29. [Google Scholar] [CrossRef] [PubMed]
- Magli, M.C.; Jones, G.M.; Lundin, K.; Van Den Abbeel, E. Atlas of human embryology: From oocytes to preimplantation embryos. Preface. Hum. Reprod. 2012, 27, i2–i21. [Google Scholar] [CrossRef]
- Silvestri, S.; Orlando, P.; Armeni, T.; Padella, L.; Brugè, F.; Seddaiu, G.; Littarru, G.P.; Tiano, L. Coenzyme Q10 and α-lipoic acid: Antioxidant and pro-oxidant effects in plasma and peripheral blood lymphocytes of supplemented subjects. J. Clin. Biochem. Nutr. 2015, 57, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Stouffer, R.L.; Xu, F.; Duffy, D.M. Molecular control of ovulation and luteinization in the primate follicle. Front. Biosci. 2007, 12, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.; Cottell, E.; Gibney, M.J.; McAuliffe, F.M.; Wingfield, M.; Brennan, L. An investigation into the relationship between the metabolic profile of follicular fluid, oocyte developmental potential, and implantation outcome. Fertil. Steril. 2012, 97, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative stress and its implications in female infertility—A clinician’s perspective. Reprod. Biomed. Online 2005, 11, 641–650. [Google Scholar] [CrossRef]
- Combelles, C.M.; Holick, E.A.; Paolella, L.J.; Walker, D.C.; Wu, Q. Profiling of superoxide dismutase isoenzymes in compartments of the developing bovine antral follicles. Reproduction 2010, 139, 871–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appasamy, M.; Jauniaux, E.; Serhal, P.; Al-Qahtani, A.; Groome, N.P.; Muttukrishna, S. Evaluation of the relationship between follicular fluid oxidative stress, ovarian hormones and response to gonadotropin stimulation. Fertil. Steril. 2008, 89, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Serke, H.; Bausenwein, J.; Hirrlinger, J.; Nowicki, M.; Vilser, C.; Jogschies, P.; Hmeidan, F.A.; Blumenauer, V.; Spanel-Borowski, K. Granulosa cell subtypes vary in response to oxidized low-density lipoprotein as regards specific lipoprotein receptors and antioxidant enzyme activity. J. Clin. Endocrinol. Metab. 2010, 95, 3480–3490. [Google Scholar] [CrossRef] [PubMed]
- Bentov, Y.; Yavorska, T.; Esfandiari, N.; Jurisicova, A.; Casper, R.F. The contribution of mitochondrial function to reproductive aging. J. Assist. Reprod. Genet. 2011, 28, 773–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweigert, F.J.; Steinhagen, B.; Raila, J.; Siemann, A.; Peet, D.; Buscher, U. Concentrations of carotenoids, retinol and alpha-tocopherol in plasma and follicular fluid of women undergoing IVF. Hum. Reprod. 2003, 18, 1259–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, R.W.; Bloom, M.S.; Shelly, W.B.; Ocque, A.J.; Huddleston, H.G.; Fujimoto, V.Y. Follicular fluid high density lipoprotein-associated micronutrient levels are associated with embryo fragmentation during IVF. J. Assist. Reprod. Genet. 2009, 26, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Otte, S.; Paletta, J.R.; Becker, S.; König, S.; Fobker, M.; Greb, R.R.; Kiesel, L.; Assmann, G.; Diedrich, K.; Nofer, J.R. Follicular fluid high density lipoprotein-associated sphingosine 1-phosphate is a novel mediator of ovarian angiogenesis. J. Biol. Chem. 2006, 281, 5398–5405. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.W.; Shelly, W.B.; Bloom, M.S.; Ocque, A.J.; Sandler, J.R.; Huddleston, H.G.; Fujimoto, V.Y. Distributions of high-density lipoprotein particle components in human follicular fluid and sera and their associations with embryo morphology parameters during IVF. Hum. Reprod. 2008, 23, 1884–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugè, F.; Bacchetti, T.; Principi, F.; Scarpa, E.S.; Littarru, G.P.; Tiano, L. Olive oil supplemented with Coenzyme Q(10): Effect on plasma and lipoprotein oxidative status. Biofactors 2012, 38, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Gautier, T.; Becker, S.; Drouineaud, V.; Ménétrier, F.; Sagot, P.; Nofer, J.R.; Von Otte, S.; Lagrost, L.; Masson, D.; Tietge, U.J. Human luteinized granulosa cells secrete apoB100-containing lipoproteins. J. Lipid Res. 2010, 51, 2245–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyawoye, O.; Abdel Gadir, A.; Garner, A.; Constantinovici, N.; Perrett, C.; Hardiman, P. Antioxidants and reactive oxygen species in follicular fluid of women undergoing IVF: Relationship to outcome. Hum. Reprod. 2003, 18, 2270–2274. [Google Scholar] [CrossRef] [PubMed]
- Pasqualotto, E.B.; Agarwal, A.; Sharma, R.K.; Izzo, V.M.; Pinotti, J.A.; Joshi, N.J.; Rose, B.I. Effect of oxidative stress in follicular fluid on the outcome of assisted reproductive procedures. Fertil. Steril. 2004, 81, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, L.; Wilson, C.; Lower, A.; Al-Shawaf, T.; Grudzinskas, J.G. Superoxide dismutase activity in human follicular fluid after controlled ovarian hyperstimulation in women undergoing in vitro fertilization. Fertil. Steril. 1999, 72, 1027–1034. [Google Scholar] [CrossRef]
- Gupta, S.; Choi, A.; Yu, H.Y.; Czerniak, S.M.; Holick, E.A.; Paolella, L.J.; Agarwal, A.; Combelles, C.M. Fluctuations in total antioxidant capacity, catalase activity and hydrogen peroxide levels of follicular fluid during bovine folliculogenesis. Reprod. Fertil. Dev. 2011, 23, 673–680. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
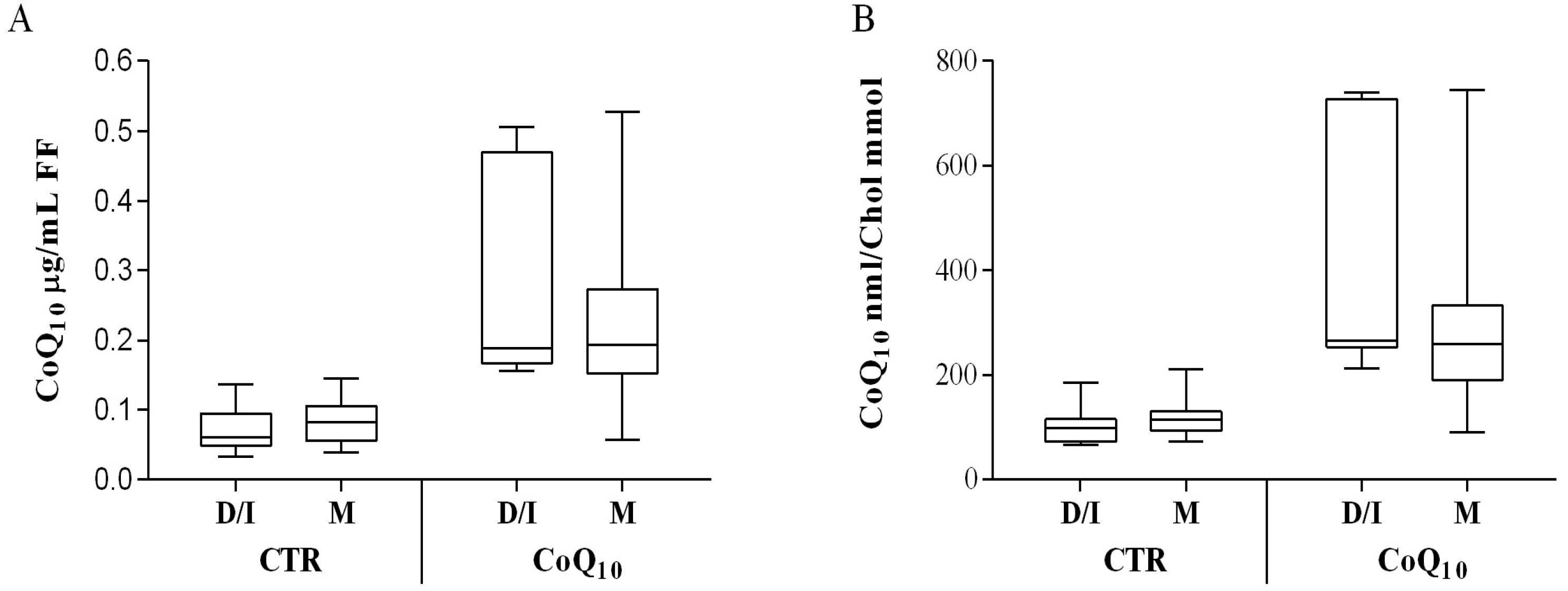{kind=link}
{kind=link}
{kind=link}
| PARAMETERS | CoQ10 Group | CTR Group |
|---|---|---|
| AGE (years) | 31–41 (32.7 mean) | 32–46 (34.6 mean) |
| BMI | 21.1 | 20.9 |
| INFERTILITY (months) | 24–48 | 24–48 |
| N° CYCLES | 15 | 15 |
| FSH | 1600–2100 | 1750–2300 |
| GROUPS | Fertilization rate | EG-I | EG-I | EG-III |
|---|---|---|---|---|
| CoQ10 | 22/25 (88%) | 18 (82%) | 4 (18%) | - |
| CTR | 20/27 (74%) | 12 (60%) | 5 (25%) | 3 (15%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannubilo, S.R.; Orlando, P.; Silvestri, S.; Cirilli, I.; Marcheggiani, F.; Ciavattini, A.; Tiano, L. CoQ10 Supplementation in Patients Undergoing IVF-ET: The Relationship with Follicular Fluid Content and Oocyte Maturity. Antioxidants 2018, 7, 141. https://doi.org/10.3390/antiox7100141
Giannubilo SR, Orlando P, Silvestri S, Cirilli I, Marcheggiani F, Ciavattini A, Tiano L. CoQ10 Supplementation in Patients Undergoing IVF-ET: The Relationship with Follicular Fluid Content and Oocyte Maturity. Antioxidants. 2018; 7(10):141. https://doi.org/10.3390/antiox7100141
Chicago/Turabian StyleGiannubilo, Stefano Raffaele, Patrick Orlando, Sonia Silvestri, Ilenia Cirilli, Fabio Marcheggiani, Andrea Ciavattini, and Luca Tiano. 2018. "CoQ10 Supplementation in Patients Undergoing IVF-ET: The Relationship with Follicular Fluid Content and Oocyte Maturity" Antioxidants 7, no. 10: 141. https://doi.org/10.3390/antiox7100141