In Vivo Effects of Methionine Sulfoxide Reductase Deficiency in Drosophila melanogaster

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Stocks
2.2. Generation of Deletion Mutations in MSR Loci
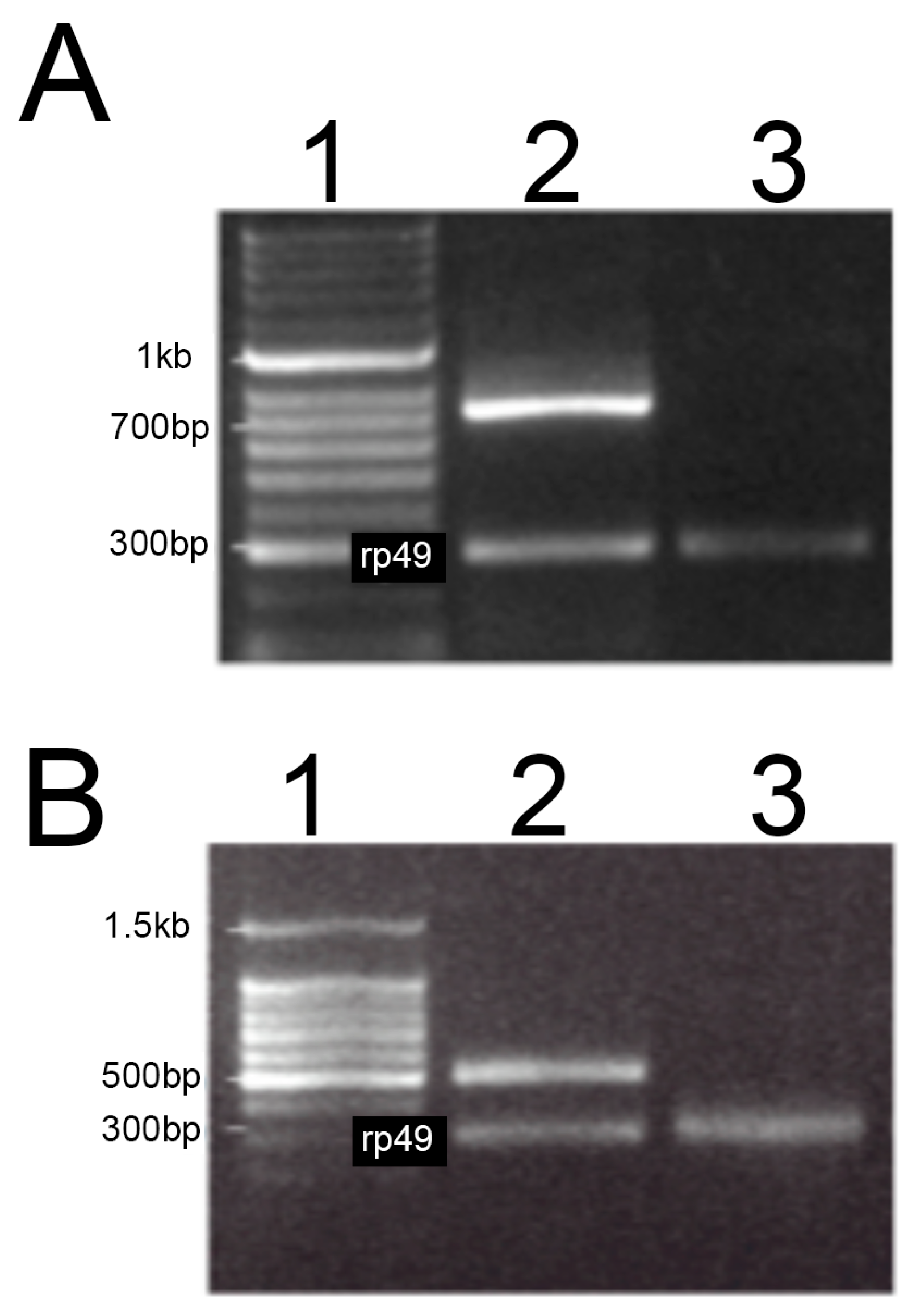
2.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.4. Western Blotting
2.5. Lifespan
2.6. Oxidative Stress Assay
2.7. Protein Carbonyl Assays
2.8. Larval Development
2.9. Determination of Wet and Dry Larval Mass
2.10. Mouth Hook Contraction Assay
2.11. Measurement of 32P-Labeled Food Intake
2.12. Statistics
3. Results
3.1. Characterization of Lines Having Complete Loss-Of-Function (Null) Alleles of MSRA and MSRB
3.2. MSR Deficiency Shortens Lifespan
3.3. Protein Oxidation
3.4. MSR Mutants Are Not More Sensitive to Oxidative Stress
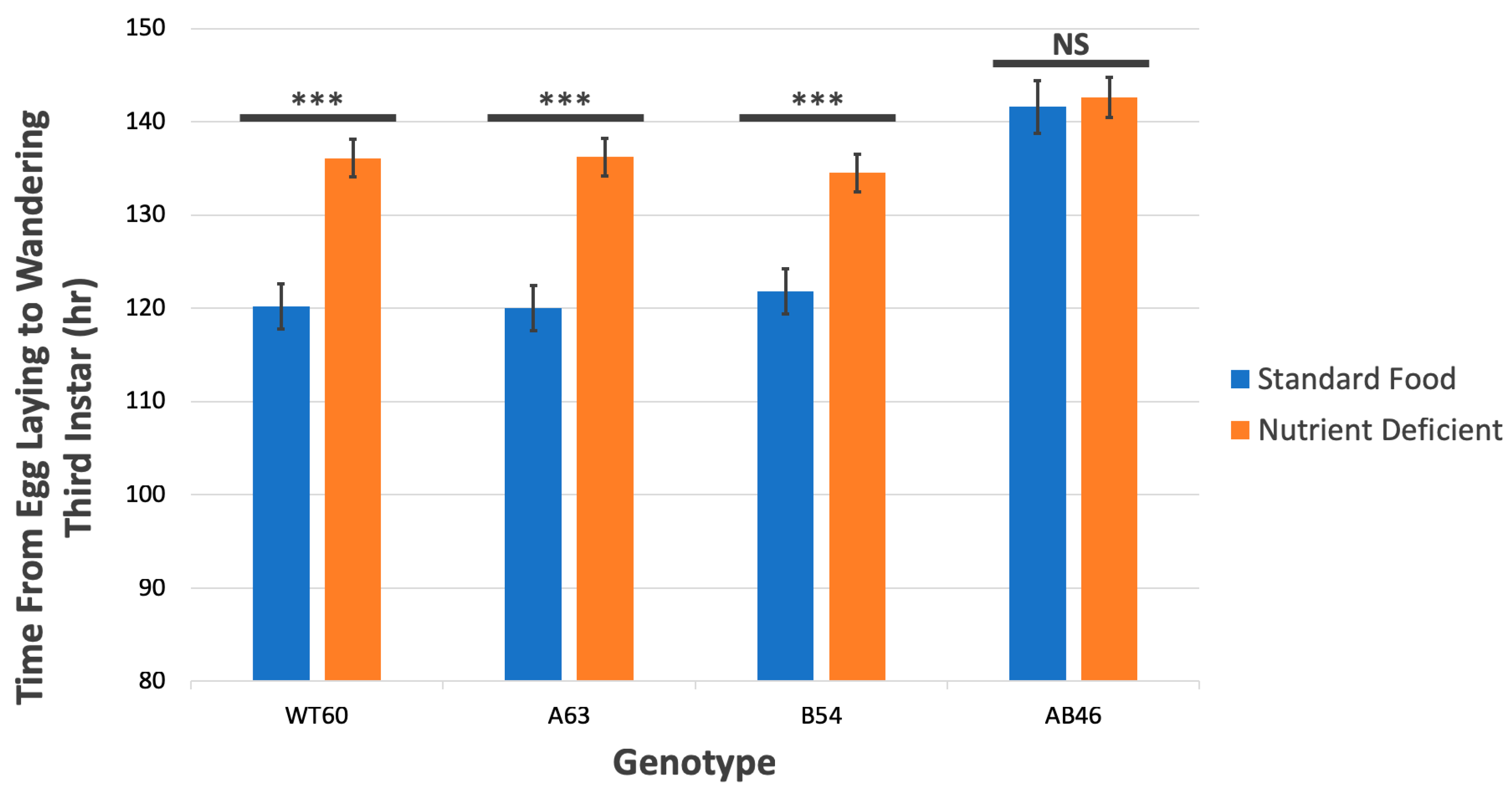
3.5. MSR Deficiency Delays Third Instar Larval Development
3.6. MSR-Deficient Larvae Have Significantly Less Mass at 96 h after Egg Laying
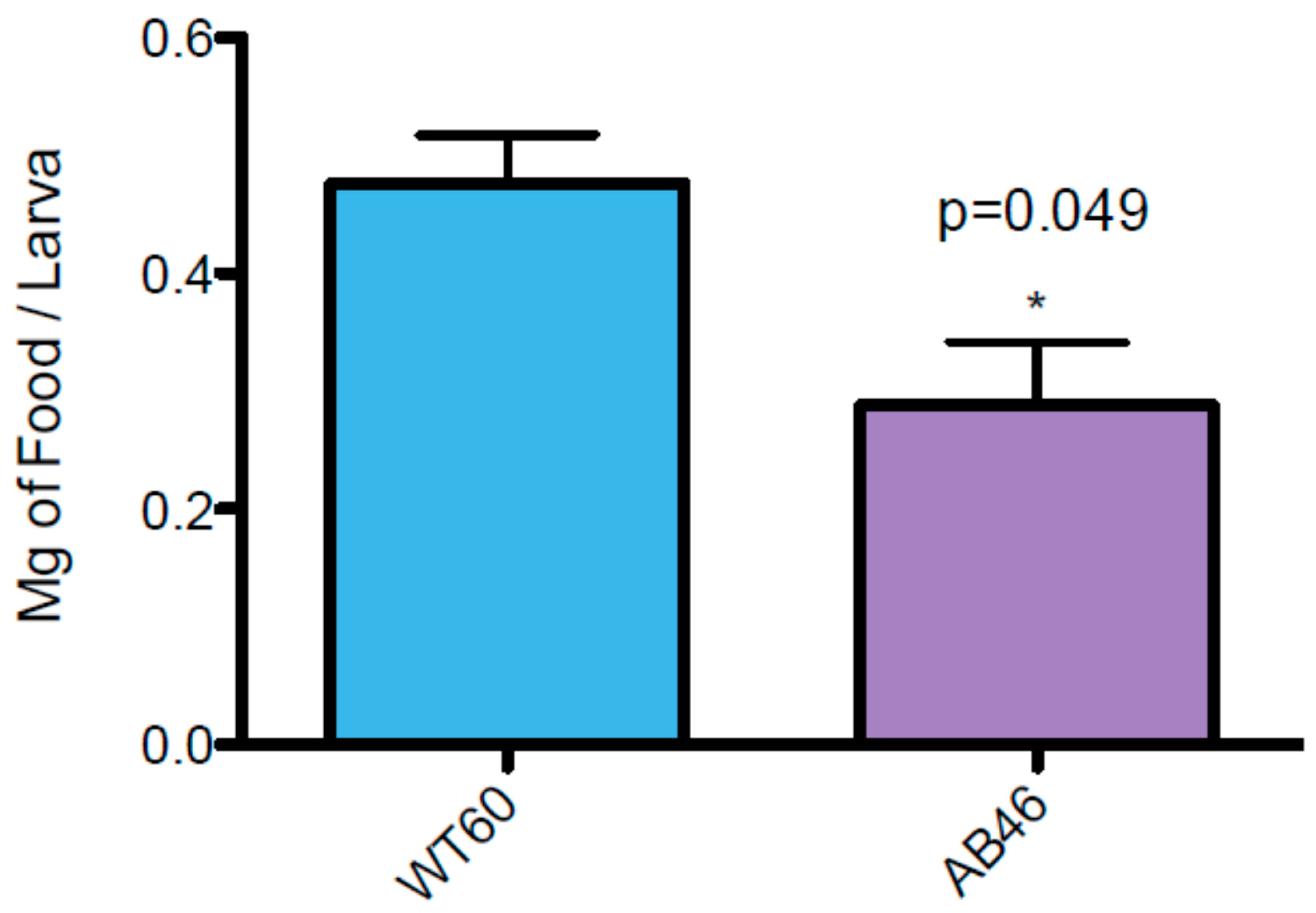
3.7. MSR-Deficient Larvae Consume Food at a Slower Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef] [PubMed]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S. Role of oxidative stress and protein oxidation in the aging process. Free Radic. Biol. Med. 2002, 33, 37–44. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Berlett, B.S. Reactive oxygen-mediated protein oxidation in aging and disease. Drug Metab. Rev. 1998, 30, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, K.; Khurana, S.; Tai, T.C. Oxidative stress in aging-matters of the heart and mind. Int. J. Mol. Sci. 2013, 14, 17897–17925. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of oxidant stress that are clinically relevant in aging and age-related disease. Mech. Ageing Dev. 2013, 134, 139–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.L.; Li, X.; Gu, Y.; Wright, P.M.; Chang, D.Y. Repair of oxidative DNA damage: Mechanisms and functions. Cell Biochem. Biophys. 2001, 35, 141–170. [Google Scholar] [CrossRef]
- Bolli, R.; Dawn, B.; Xuan, Y.T. Role of the JAK-STAT pathway in protection against myocardial ischemia/reperfusion injury. Trends Cardiovasc. Med. 2003, 13, 72–79. [Google Scholar] [CrossRef]
- Cudic, P.; Joshi, N.; Sagher, D.; Williams, B.T.; Stawikowski, M.J.; Weissbach, H. Identification of activators of methionine sulfoxide reductases A and B. Biochem. Biophys. Res. Commun. 2016, 469, 863–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, H. Ischemic preconditioning: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2008, 10, 207–247. [Google Scholar] [CrossRef] [PubMed]
- Weissbach, H.; Resnick, L.; Brot, N. Methionine sulfoxide reductases: History and cellular role in protecting against oxidative damage. Biochim. Biophys. Acta 2005, 1703, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.J.; Cupp, L.E.; Weissbach, H.; Brot, N. Synthesis of ribosomal proteins L7L12 in relaxed and stringent strains of Escherichia coli. J. Biol. Chem. 1976, 251, 5516–5521. [Google Scholar] [PubMed]
- Brot, N.; Weissbach, L.; Werth, J.; Weissbach, H. Enzymatic reduction of protein-bound methionine sulfoxide. Proc. Natl. Acad. Sci. USA 1981, 78, 2155–2158. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in protein. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef] [PubMed]
- Kantorow, M.; Hawse, J.R.; Cowell, T.L.; Benhamed, S.; Pizarro, G.O.; Reddy, V.N.; Hejtmancik, J.F. Methionine sulfoxide reductase a is important for lens cell viability and resistance to oxidative stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9654–9659. [Google Scholar] [CrossRef] [PubMed]
- Yermolaieva, O.; Xu, R.; Schinstock, C.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Hoshi, T. Methionine sulfoxide reductase a protects neuronal cells against brief hypoxia/reoxygenation. Proc. Natl. Acad. Sci. USA 2004, 101, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Avdonin, V.; Ciorba, M.A.; Heinemann, S.H.; Hoshi, T. Acceleration of P/C-type inactivation in voltage-gated K+ channels by methionine oxidation. Biophys. J. 2000, 78, 174–187. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Heinemann, S.H.; Weissbach, H.; Brot, N.; Hoshi, T. Modulation of potassium channel function by methionine oxidation and reduction. Proc. Natl. Acad. Sci. USA 1997, 94, 9932–9937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciorba, M.A.; Heinemann, S.H.; Wiessbach, H.; Brot, N.; Hoshi, T. Regulation of voltage-dependent K+ channels by methionine oxidation: Effect of nitric oxide and vitamin C. FEBS Lett. 1999, 44, 48–52. [Google Scholar] [CrossRef]
- Li, A.; Segui, J.; Heinemann, S.H.; Hoshi, T. Oxidation regulates cloned neuronal voltage-dependent Ca2+ channels expressed in xenopus oocytes. J. Neuro Sci. 1998, 18, 6740–6747. [Google Scholar] [CrossRef]
- Hung, R.J.; Spaeth, C.S.; Yesilyurt, H.G.; Terman, J.R. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 2013, 15, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskovitz, J. Methionine sulfoxide reductases: Ubiquitous enzymes involved in antioxidant defense, protein regulation, and prevention of aging-associated diseases. Biochim. Biophys. Acta 2005, 1703, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Bar-Noy, S.; Williams, W.M.; Requena, J.; Berlett, B.S.; Stadtman, E.R. Methionine sulfoxide reductase (MSRA) is a regulator of antioxidant defense and lifespan in mammals. Proc. Natl. Acad. Sci. USA 2001, 98, 12920–12925. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Berlett, B.S.; Poston, J.M.; Stadtman, E.R. The yeast peptide-methionine sulfoxide reductase functions as an antioxidant in vivo. Proc. Natl. Acad. Sci. USA 1997, 94, 9585–9589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskovitz, J.; Rahman, M.A.; Strassman, J.; Yancey, S.O.; Kushner, S.R.; Brot, N.; Weissbach, H. Escherichia coli peptide methionine sulfoxide reductase gene: Regulation of expression and role in protecting against oxidative damage. J. Bacteriol. 1995, 177, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Tang, X.D.; Chen, M.-L.; Joiner, M.A.; Sun, G.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Iverson, L.; Wu, C.-F.; et al. High-quality life extension by the enzyme peptide methionine sulfoxide reductase. Proc. Natl. Acad. Sci. USA 2002, 99, 2748–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Kim, A.K.; Jung, S.A.; Kim, S.W.; Yu, K.; Lee, J.H. The Drosophila homolog of methionine sulfoxide reductase a extends lifespan and increases nuclear localization of FOXO. FEBS Lett. 2010, 584, 3609–3614. [Google Scholar] [CrossRef] [PubMed]
- Minniti, A.N.; Cataldo, R.; Trigo, C.; Vasquez, L.; Mujica, P.; Leighton, F.; Inestrosa, N.C.; Aldunate, R. Methionine sulfoxide reductase A expression is regulated by the DAF-16/FOXO pathway in Caenorhabditis elegans. Aging Cell 2009, 8, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Koc, A.; Gasch, A.P.; Rutherford, J.C.; Kim, H.Y.; Gladyshev, V.N. Methionine sulfoxide reductase regulation of yeast lifespan reveals reactive oxygen species-dependent and -independent components of aging. Proc. Natl. Acad. Sci. USA 2004, 101, 7999–8004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, A.B.; Kim, G.; Liu, C.; Wren, J.D.; Georgescu, C.; Richardson, A.; Levine, R.L. Effects of transgenic methionine sulfoxide reductase a (MSRA) expression on lifespan and age-dependent changes in metabolic function in mice. Redox Biol. 2016, 10, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Stadtman, E.R. Selenium-deficient diet enhances protein oxidation and affects methionine sulfoxide reductase (MSRB) protein level in certain mouse tissues. Proc. Natl. Acad. Sci. USA 2003, 100, 7486–7490. [Google Scholar] [CrossRef] [PubMed]
- Shchedrina, V.A.; Vorbruggen, G.; Lee, B.C.; Kim, H.Y.; Kabil, H.; Harshman, L.G.; Gladyshev, V.N. Overexpression of methionine-r-sulfoxide reductases has no influence on fruit fly aging. Mech. Ageing Dev. 2009, 130, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.H.; Han, J.Y.; Kim, J.R.; Lee, Y.S.; Kim, H.Y. Methionine sulfoxide reductase B in the endoplasmic reticulum is critical for stress resistance and aging in drosophila. Biochem. Biophys. Res. Commun. 2012, 419, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Liu, C.; Levine, R. Generation of methionine sulfoxide reductase quadruple knockout mice. Free Radic. Biol. Med. 2016, 100, S103. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Williams, J.A.; Stadtman, E.R.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.K.; Neckameyer, W.S. Functional analysis of the larval feeding circuit in drosophila. J. Vis. Exp. JoVE 2013, 81, e51062. [Google Scholar] [CrossRef] [PubMed]
- Shimada-Niwa, Y.; Niwa, R. Serotonergic neurons respond to nutrients and regulate the timing of steroid hormone biosynthesis in drosophila. Nat. Commun. 2014, 5, 5778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Vaish, M.; Johansson, T.R.; Baum, K.R.; Ring, R.P.; Singh, S.; Shukla, S.K.; Moskovitz, J. Significance of four methionine sulfoxide reductases in Staphylococcus aureus. PLoS ONE 2015, 10, e0117594. [Google Scholar] [CrossRef] [PubMed]
- Salmon, A.B.; Perez, V.I.; Bokov, A.; Jernigan, A.; Kim, G.; Zhao, H.; Levine, R.L.; Richardson, A. Lack of methionine sulfoxide reductase a in mice increases sensitivity to oxidative stress but does not diminish life span. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 3601–3608. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Gladyshev, V.N. Methionine sulfoxide reduction in mammals: Characterization of methionine-R-sulfoxide reductases. Mol. Biol. Cell 2004, 15, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Weissbach, H.; Etienne, F.; Hoshi, T.; Heinemann, S.H.; Lowther, W.T.; Matthews, B.; St John, G.; Nathan, C.; Brot, N. Peptide methionine sulfoxide reductase: Structure, mechanism of action, and biological function. Arch. Biochem. Biophys. 2002, 397, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.A.; Koc, A.; Cerny, R.L.; Gladyshev, V.N. Reaction mechanism, evolutionary analysis, and role of zinc in drosophila methionine-r-sulfoxide reductase. J. Biol. Chem. 2002, 277, 37527–37535. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.D.; Zimmermann, G.; Suyama, K.; Meyer, T.; Scott, M.P. A nucleostemin family GTPase, NS3, acts in serotonergic neurons to regulate insulin signaling and control body size. Genes Dev. 2008, 22, 1877–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daubert, E.A.; Heffron, D.S.; Mandell, J.W.; Condron, B.G. Serotonergic dystrophy induced by excess serotonin. Mol. Cell. Neurosci. 2010, 44, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | Strain Designation | Genotype |
|---|---|---|
| Wild-type | WT60 | MSRA+/+MSRB+/+ |
| No MSRA | A63 | MSRAΔ/ΔMSRB+/+ |
| No MSRB | B54 | MSRA+/+MSRBΔ/Δ |
| No MSR | AB46 | MSRAΔ/ΔMSRBΔ/Δ |
| Developmental Stage | MSRA+/+MSRB+/+ | MSRAΔ/ΔMSRB+/+ | MSRA+/+MSRBΔ/Δ | MSRAΔ/ΔMSRBΔ/Δ |
|---|---|---|---|---|
| First Instar | 26.0 h | 26.0 h | 25.5 h | 25.7 h |
| Second Instar | 51.1 h | 51.3 h | 50.1 h | 51.1 h |
| Third Instar | 77.5 h | 77.2 h | 76.2 h | 78.0 h |
| Wandering Third Instar | 120.2 h | 120.0 h | 121.8 h | 141.6 h |
| Pupariation | 127.5 h | 127.3 h | 128.0 h | 147.4 h |
| Eclosion | 228.2 h | 228.6 h | 225.7 h | 247.6 h |
| Developmental Stage | MSRA+/+MSRB+/+ | MSRAΔ/ΔMSRB+/+ | MSRA+/+MSRBΔ/Δ | MSRAΔ/ΔMSRBΔ/Δ |
|---|---|---|---|---|
| 72 h | 0.37 mg | 0.38 mg | 0.37 mg | 0.34 mg |
| 96 h | 1.34 mg | 1.35 mg | 1.35 mg | 0.72 mg |
| Wandering Third Instar | 1.87 mg | 1.88 mg | 1.86 mg | 1.83 mg |
| Newly Eclosed Adult | 1.22 mg | 1.26 mg | 1.23 mg | 1.19 mg |
| 6-Day Old Adult | 1.51 mg | 1.51 mg | 1.57 mg | 1.44 mg |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruce, L.; Singkornrat, D.; Wilson, K.; Hausman, W.; Robbins, K.; Huang, L.; Foss, K.; Binninger, D. In Vivo Effects of Methionine Sulfoxide Reductase Deficiency in Drosophila melanogaster. Antioxidants 2018, 7, 155. https://doi.org/10.3390/antiox7110155
Bruce L, Singkornrat D, Wilson K, Hausman W, Robbins K, Huang L, Foss K, Binninger D. In Vivo Effects of Methionine Sulfoxide Reductase Deficiency in Drosophila melanogaster. Antioxidants. 2018; 7(11):155. https://doi.org/10.3390/antiox7110155
Chicago/Turabian StyleBruce, Lindsay, Diana Singkornrat, Kelsey Wilson, William Hausman, Kelli Robbins, Lingxi Huang, Katie Foss, and David Binninger. 2018. "In Vivo Effects of Methionine Sulfoxide Reductase Deficiency in Drosophila melanogaster" Antioxidants 7, no. 11: 155. https://doi.org/10.3390/antiox7110155