Identification of Phenolic Compounds-Rich Grape Pomace Extracts Urine Metabolites and Correlation with Gut Microbiota Modulation

 ,
, 

Abstract
1. Introduction
2. Short Methods
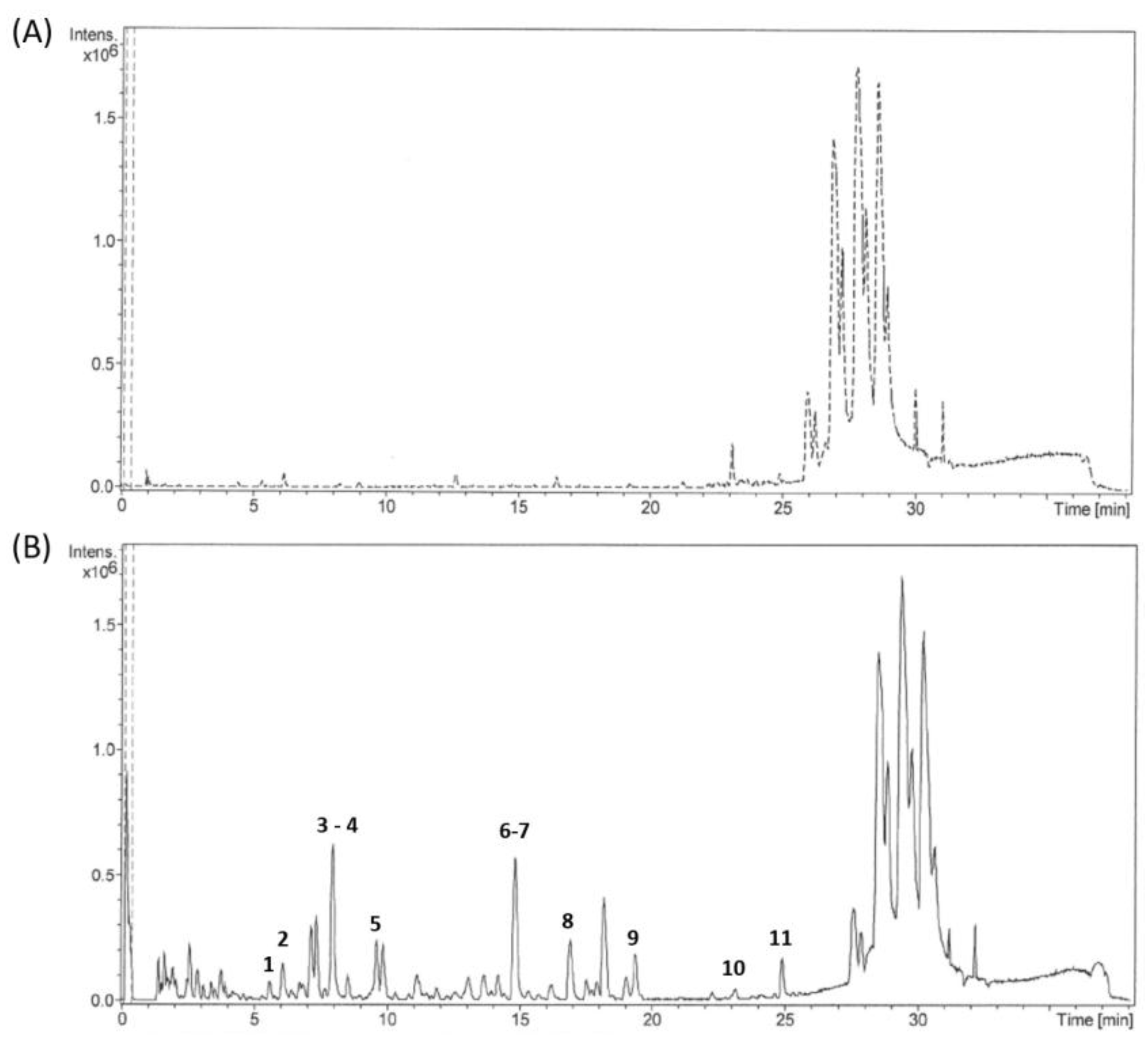
Urine Analysis by LC-QTOF Mass Spectrometry
3. Results
Identification of Urinary Metabolites Following Grape Pomace Extracts Consumption
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Feliciano, R.P.; Istas, G.; Heiss, C.; Rodriguez-Mateos, A. Plasma and Urinary Phenolic Profiles after Acute and Repetitive Intake of Wild Blueberry. Molecules 2016, 21, E1120. [Google Scholar] [CrossRef] [PubMed]
- Gasperotti, M.; Passamonti, S.; Tramer, F.; Masuero, D.; Guella, G.; Mattivi, F.; Vrhovsek, U. Fate of microbial metabolites of dietary polyphenols in rats: Is the brain their target destination? ACS Chem. Neurosci. 2015, 6, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Pimpao, R.C.; Dew, T.; Oliveira, P.B.; Williamson, G.; Ferreira, R.B.; Santos, C.N. Analysis of phenolic compounds in Portuguese wild and commercial berries after multienzyme hydrolysis. J. Agric. Food Chem. 2013, 61, 4053–4062. [Google Scholar] [CrossRef] [PubMed]
- Espin, J.C.; Gonzalez-Sarrias, A.; Tomas-Barberan, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Chacar, S.; Itani, T.; Hajal, J.; Saliba, Y.; Louka, N.; Faivre, J.F.; Maroun, R.; Fares, N. The Impact of Long-Term Intake of Phenolic Compounds-Rich Grape Pomace on Rat Gut Microbiota. J. Food Sci. 2018, 83, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Popa, D.E.; Dragoi, C.M.; Arsene, A.L.; Dumitrescu, I.B.; Nicolae, A.C.; Velescu, B.S.; Burcea-Dragomiroiu, G.T.A. The relationship between phenolic compounds from diet and microbiota. Food Funct. 2015, 6, 2424–2439. [Google Scholar]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Dolara, P.; Luceri, C.; De Filippo, C.; Femia, A.P.; Giovannelli, L.; Caderni, G.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Cresci, A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. 2005, 591, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Gwiazdowska, D.; Jus, K.; Jasnowska-Malecka, J.; Kluczynska, K. The impact of polyphenols on Bifidobacterium growth. Acta Biochim. Pol. 2015, 62, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Renouf, M.; Marmet, C.; Guy, P.A.; Beaumont, M.; Lepage, M.; Williamson, G.; Dionisi, F. Dose-response plasma appearance of green tea catechins in adults. Mol. Nutr. Food Res. 2013, 57, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Zaffarano, J.I. Minimum Inhibitory Concentrations of Two Common Food Phenolic Compounds and Their Effect on the Microbial Ecology of Swine Feces in Vitro. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2003. [Google Scholar]
- Cetin-Karaka, H.; Newman, M.C. Antimicrobial Efficacy of Natural Phenolic Compounds against Gram Positive Foodborne Pathogens. J. Food Res. 2015, 4, 14–27. [Google Scholar] [CrossRef]
- Dong, X.; Xu, W.; Sikes, R.A.; Wu, C. Combination of low dose of genistein and daidzein has synergistic preventive effects on isogenic human prostate cancer cells when compared with individual soy isoflavone. Food Chem. 2013, 141, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Compound Structure | PC Metabolites | PC 2.5 | PC 5 | PC 10 | PC 20 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 6 mo pt | 14 mo pt | 6 mo pt | 14 mo pt | 6 mo pt | 14 mo pt | 6 mo pt | 14 mo pt | ||
| M-1 derivatives | Coumaric acid | − | − | + | − | + | − | − | − |
| Genistein | + | − | + | − | + | + | + | + | |
| Daidzedin | + | + | + | + | + | + | + | + | |
| Valerolactone | + | − | − | − | − | + | − | − | |
| Tyrosol | − | − | − | + | − | − | − | − | |
| Trans-cinnamic acid | − | − | − | + | − | − | − | + | |
| Apigenin | − | − | − | − | − | − | − | + | |
| Phenolic sulfate derivatives | Pyrocatechol | + | + | + | + | + | + | + | + |
| 3-hydroxyphenylacetic acid | + | + | + | + | + | + | + | + | |
| Homovanillic acid | + | + | + | + | + | + | + | + | |
| Ferulic acid | + | + | + | + | + | + | + | + | |
| Enterolactone | − | − | − | − | − | − | − | + | |
| Benzoic acid | + | + | + | + | + | + | + | + | |
| Daidzedin | − | + | − | + | − | + | − | + | |
| Phenolic methyl derivatives | 3-hydroxyphenylacetic acid | + | + | + | + | − | − | − | − |
| Phenolic glucuronide derivatives | 2-(4-hydroxyphenyl) propionic acid | + | + | + | + | − | − | − | − |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacar, S.; Tarighi, M.; Fares, N.; Faivre, J.-F.; Louka, N.; Maroun, R.G. Identification of Phenolic Compounds-Rich Grape Pomace Extracts Urine Metabolites and Correlation with Gut Microbiota Modulation. Antioxidants 2018, 7, 75. https://doi.org/10.3390/antiox7060075
Chacar S, Tarighi M, Fares N, Faivre J-F, Louka N, Maroun RG. Identification of Phenolic Compounds-Rich Grape Pomace Extracts Urine Metabolites and Correlation with Gut Microbiota Modulation. Antioxidants. 2018; 7(6):75. https://doi.org/10.3390/antiox7060075
Chicago/Turabian StyleChacar, Stéphanie, Mehrad Tarighi, Nassim Fares, Jean-François Faivre, Nicolas Louka, and Richard G. Maroun. 2018. "Identification of Phenolic Compounds-Rich Grape Pomace Extracts Urine Metabolites and Correlation with Gut Microbiota Modulation" Antioxidants 7, no. 6: 75. https://doi.org/10.3390/antiox7060075
APA StyleChacar, S., Tarighi, M., Fares, N., Faivre, J.-F., Louka, N., & Maroun, R. G. (2018). Identification of Phenolic Compounds-Rich Grape Pomace Extracts Urine Metabolites and Correlation with Gut Microbiota Modulation. Antioxidants, 7(6), 75. https://doi.org/10.3390/antiox7060075